
Structure–Functional Examination of Novel Ribonucleoside Hydrolase C (RihC) from Limosilactobacillus reuteri LR1

 , , , , , , and
, , , , , , and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Selecting a Proper His-Tag Position
2.2. Obtaining the LreRihC Enzymes with His-Tag
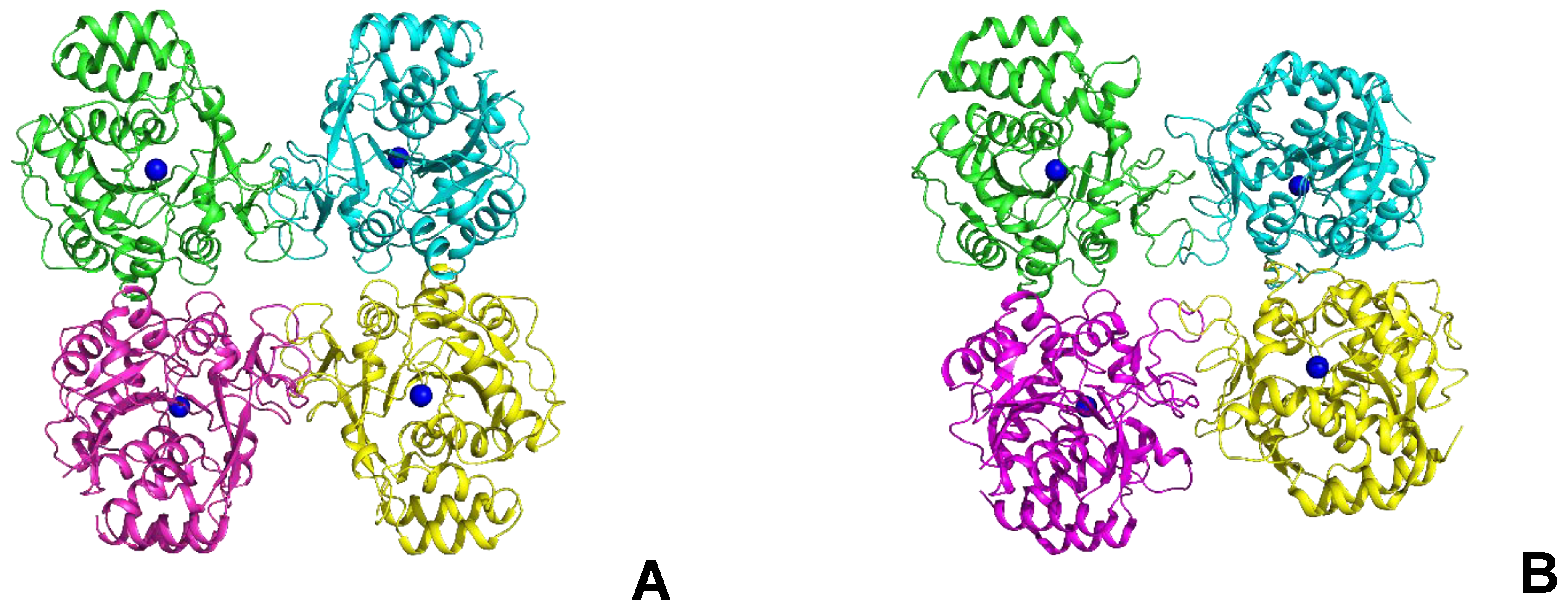
2.3. Determination of the Oligomeric Composition of the Enzyme
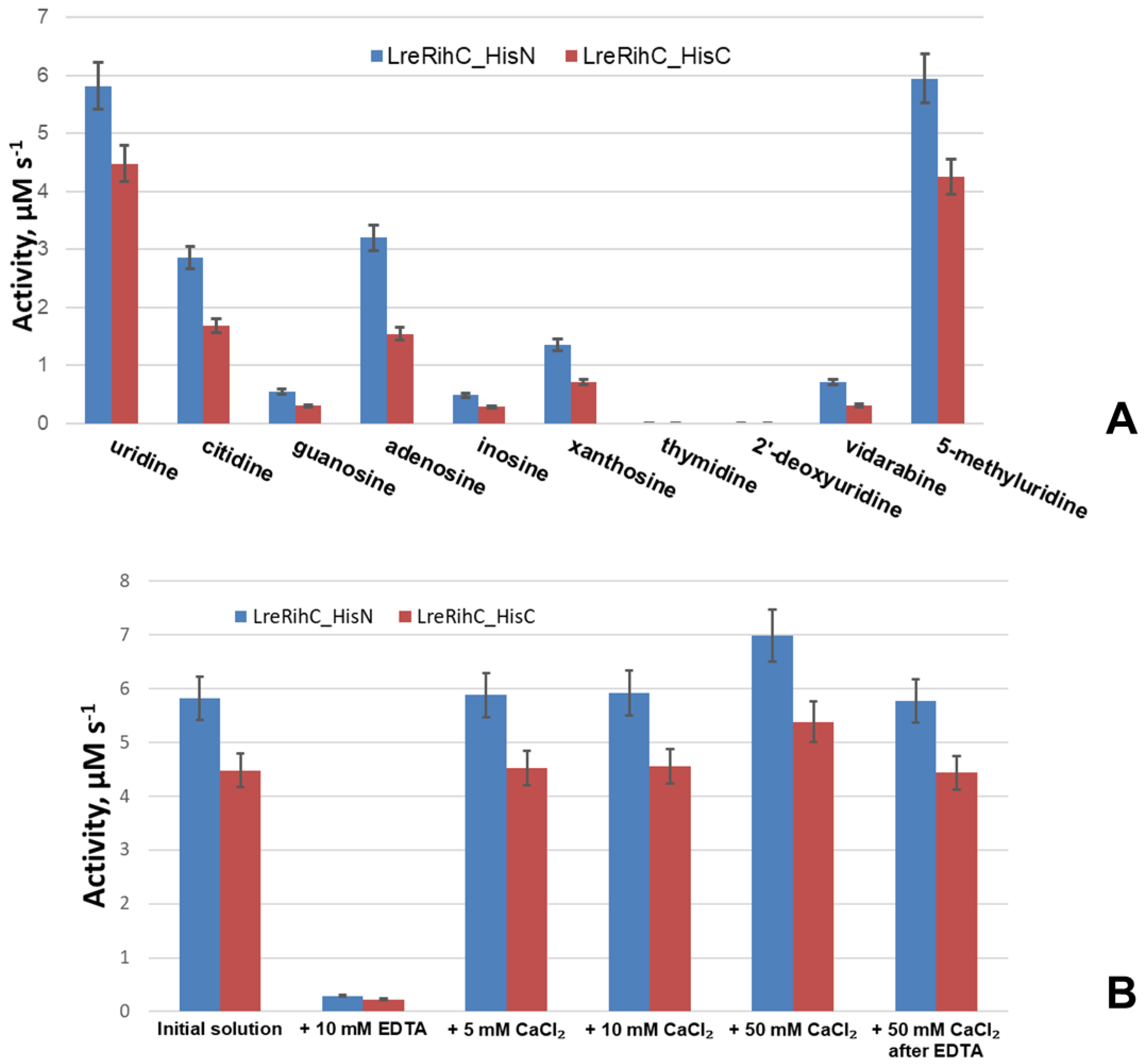
2.4. Kinetic Parameters of LreRihC

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | LreRihC_HisN | LreRihC_HisC | EcoRihC b | CfaRihC | SenRihC | LmaRihC |
|---|---|---|---|---|---|---|
| kcaturidine, s−1 | 167 ± 6 | 134 ± 6 a | 10.85 ± 0.23 | 143 | 46 ± 3 c | 32 ± 6 |
| KMuridine, µM | 320 ± 40 | 320 ± 40 a | 408 ± 184 | 1220 ± 40 | 1060 ± 100 c | 234 ± 112 |
| kcatcytidine, s−1 | 112 ± 4 | 58 ± 6 | 1.12 ± 0.53 | 20 | 7.8 ± 1.3 c | 0.36 ± 0.05 |
| KMcytidine, µM | 680 ± 80 | 620 ± 60 | 682 ± 298 | 4700 ± 500 | 9200 ± 1200 c | 422 ± 175 |
| kcatinosine, s−1 | 30 ± 5 | 18 ± 6 | 4.31 ± 0.22 | 32 | 9.0 ± 0.15 c 8.1 ± 0.14 d | 119 ± 34 |
| KMinosine, µM | 2500 ± 600 | 2600 ± 600 | 422 ± 225 | 380 ± 30 | 650 ± 60 c 1280 ± 130 d | 445 ± 209 |
| kcatxanthosine, s−1 | 57 ± 8 | 40 ± 6 | 6.30 ± 0.05 | ND | 62 ± 8 c 26.3 ± 0.3 d | ND |
| KMxanthosine, µM | 1200 ± 200 | 1300 ± 200 | 454 ± 165 | ND | 5900 ± 1000 c 790 ± 50 d | ND |
| kcatadenosine, s−1 | 118 ± 4 | 65 ± 7 | 1.15 ± 0.47 | 4.3 | 2.06 ± 0.07 c | 0.57 ± 0.04 |
| KMadenosine, µM | 420 ± 50 | 480 ± 90 | 416 ± 249 | 460 ± 30 | 160 ± 20 c | 185 ± 46 |
| kcatguanosine, s−1 | ND | ND | ND | 2 | ND | 0.59 ± 0.03 |
| KMguanosine, µM | ND | ND | ND | 420 ± 10 | ND | 140 ± 23 |
| Source | This work | This work | [7] | [10,14] | [15] | [14] |
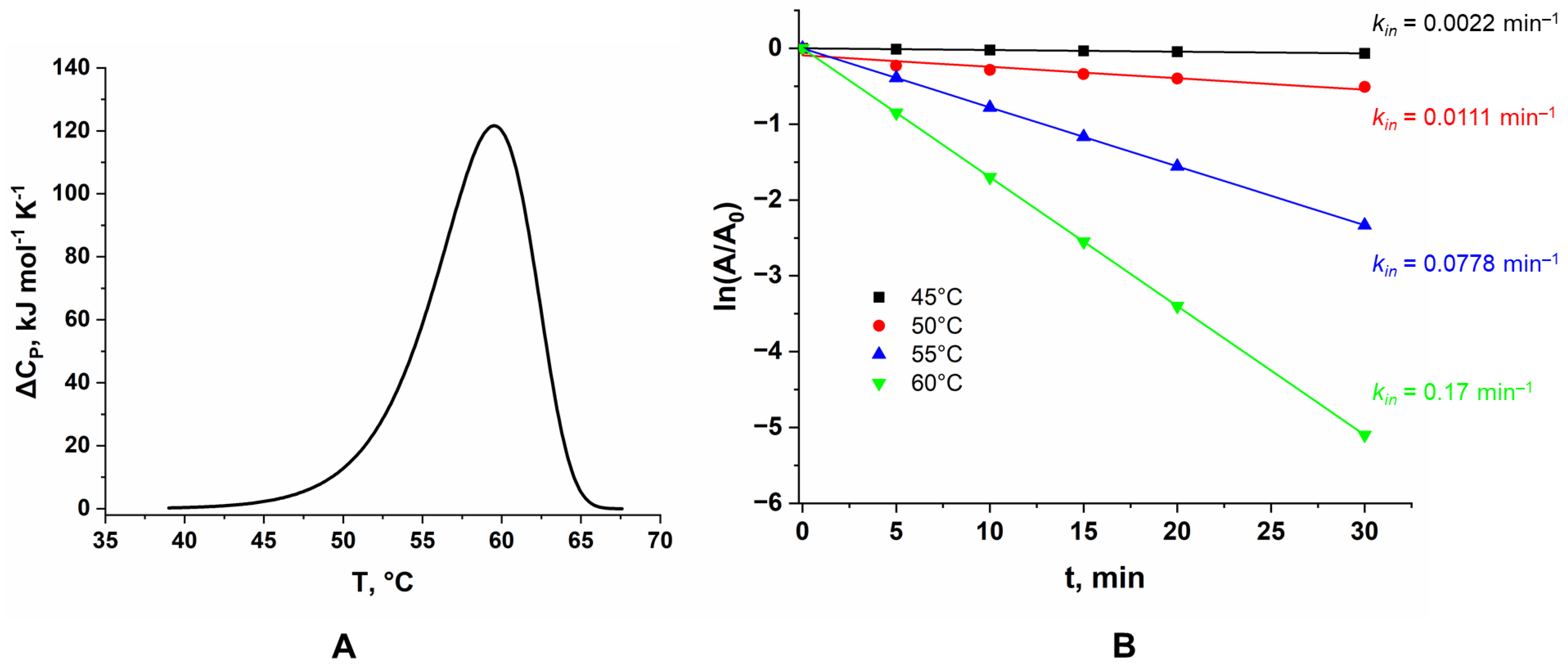
2.5. Temperature Stability of LreRihC_HisN
2.6. Structural Studies of LreRihC
3. Materials and Methods
3.1. Obtaining Model Protein Structures
3.2. Obtaining the LreRihC Genetic Constructs
3.3. Enzyme Expression in E. coli Cells
3.4. Enzyme Purification
3.5. Determination of the Oligomeric Composition of the Enzyme
3.6. Carrying Out an Enzymatic Reaction
3.7. Conducting the Analysis Using HILIC
3.8. Kinetic Parameters Determination for RihC Enzyme
3.9. Study of Enzyme Thermostability
3.10. Crystallization and Data Collection
3.11. Structure Solution and Refinement
3.12. Structure Analysis and Validation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Wyres, K.L.; Nguyen, T.N.T.; Lam, M.M.C.; Judd, L.M.; van Vinh Chau, N.; Dance, D.A.B.; Ip, M.; Karkey, A.; Ling, C.L.; Miliya, T.; et al. Genomic Surveillance for Hypervirulence and Multi-Drug Resistance in Invasive Klebsiella pneumoniae from South and Southeast Asia. Genome Med. 2020, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Nirwati, H.; Sinanjung, K.; Fahrunissa, F.; Wijaya, F.; Napitupulu, S.; Hati, V.P.; Hakim, M.S.; Meliala, A.; Aman, A.T.; Nuryastuti, T. Biofilm Formation and Antibiotic Resistance of Klebsiella pneumoniae Isolated from Clinical Samples in a Tertiary Care Hospital, Klaten, Indonesia. BMC Proc. 2019, 13, 20. [Google Scholar] [CrossRef] [PubMed]
- Kochish, A.A.; Bozhkova, S.A.; Netylko, G.I.; Anisimova, L.I. Experimental Study of Antimicrobial Polymeric Composition with Hemostatic Effect in Treatment of Implant Associated Infection. Genij Ortop. 2019, 25, 180–187. [Google Scholar] [CrossRef]
- Savinova, O.S.; Glazunova, O.A.; Moiseenko, K.V.; Begunova, A.V.; Rozhkova, I.V.; Fedorova, T.V. Exoproteome Analysis of Antagonistic Interactions between the Probiotic Bacteria Limosilactobacillus reuteri Lr1 and Lacticaseibacillus rhamnosus f and Multidrug Resistant Strain of Klebsiella Pneumonia. Int. J. Mol. Sci. 2021, 22, 10999. [Google Scholar] [CrossRef] [PubMed]
- Shaposhnikov, L.A.; Savin, S.S.; Tishkov, V.I.; Pometun, A.A. Ribonucleoside Hydrolases–Structure, Functions, Physiological Role and Practical Uses. Biomolecules 2023, 13, 1375. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.; Møller, L.B. The RihA, RihB, and RihC Ribonucleoside Hydrolases of Escherichia coli. Substrate Specificity, Gene Expression, and Regulation. J. Biol. Chem. 2001, 276, 884–894. [Google Scholar] [CrossRef]
- Arivett, B.; Farone, M.; Masiragani, R.; Burden, A.; Judge, S.; Osinloye, A.; Minici, C.; Degano, M.; Robinson, M.; Kline, P. Characterization of Inosine-Uridine Nucleoside Hydrolase (RihC) from Escherichia coli. Biochim. Biophys. Acta Proteins Proteom. 2014, 1844, 656–662. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making Protein Folding Accessible to All. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Parkin, D.W.; Horenstein, B.A.; Abdulah, D.R.; Estupinan, B.; Schramm, V.L. Nucleoside Hydrolase from Crithidia fasciculuta. Metabolic Role, Purification, Specificity, and Kinetic Mechanism. J. Biol. Chem. 1991, 266, 20658–20665. [Google Scholar] [CrossRef]
- Vandemeulebroucke, A.; Minici, C.; Bruno, I.; Muzzolini, L.; Tornaghi, P.; Parkin, D.W.; Versées, W.; Steyaert, J.; Degano, M. Structure and Mechanism of the 6-Oxopurine Nucleosidase from Trypanosoma brucei brucei. Biochemistry 2010, 49, 8999–9010. [Google Scholar] [CrossRef] [PubMed]
- Shaposhnikov, L.A.; Chikurova, N.Y.; Chernobrovkina, A.V.; Tishkov, V.I.; Pometun, A.A. Development of an Approach to Determining Enzymatic Activity of Ribonucleoside Hydrolase C Using Hydrophilic Interaction Liquid Chromatography. J. Chromatogr. A 2024, 1715, 464561. [Google Scholar] [CrossRef]
- Traut, T.W. Physiological Concentrations of Purines and Pyrimidines. Mol. Cell. Biochem. 1994, 140, 1–22. [Google Scholar] [CrossRef]
- Shi, W.; Schramm, V.L.; Almo, S.C. Nucleoside Hydrolase from Leishmania major. Cloning, Expression, Catalytic Properties, Transition State Inhibitors, and the 2.5-Å Crystal Structure. J. Biol. Chem. 1999, 274, 21114–21120. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.R.; Dandanell, G. Purification and Characterization of RihC, a Xanthosine-Inosine-Uridine- Adenosine-Preferring Hydrolase from Salmonella enterica serovar Typhimurium. Biochim. Biophys. Acta Gen. Subj. 2005, 1723, 55–62. [Google Scholar] [CrossRef] [PubMed]
- The PyMOL Molecular Graphics System; Version 2.0; Schrödinger, LLC: New York, NY, USA, 2015.
- Koshkina, M.K.; Shelomov, M.D.; Pometun, A.A.; Savin, S.S.; Tishkov, V.I.; Atroshenko, D.L. Speeding up SDS–PAGE: Theory and Experiment. Electrophoresis 2023, 44, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Kabsch, W. XDS. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef]
- Evans, P. Scaling and Assessment of Data Quality. Acta Crystallogr. D Biol. Crystallogr. 2006, 62, 72–82. [Google Scholar] [CrossRef]
- Vagin, A.A.; Isupov, M.N. Spherically Averaged Phased Translation Function and Its Application to the Search for Molecules and Fragments in Electron-Density Maps. Acta Crystallogr. D Biol. Crystallogr. 2001, 57, 1451–1456. [Google Scholar] [CrossRef]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.W.; McCoy, A.; et al. Overview of the CCP4 Suite and Current Developments. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and Development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E.; Henrick, K. Secondary-Structure Matching (SSM), a New Tool for Fast Protein Structure Alignment in Three Dimensions. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2256–2268. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E.; Henrick, K. Inference of Macromolecular Assemblies from Crystalline State. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]
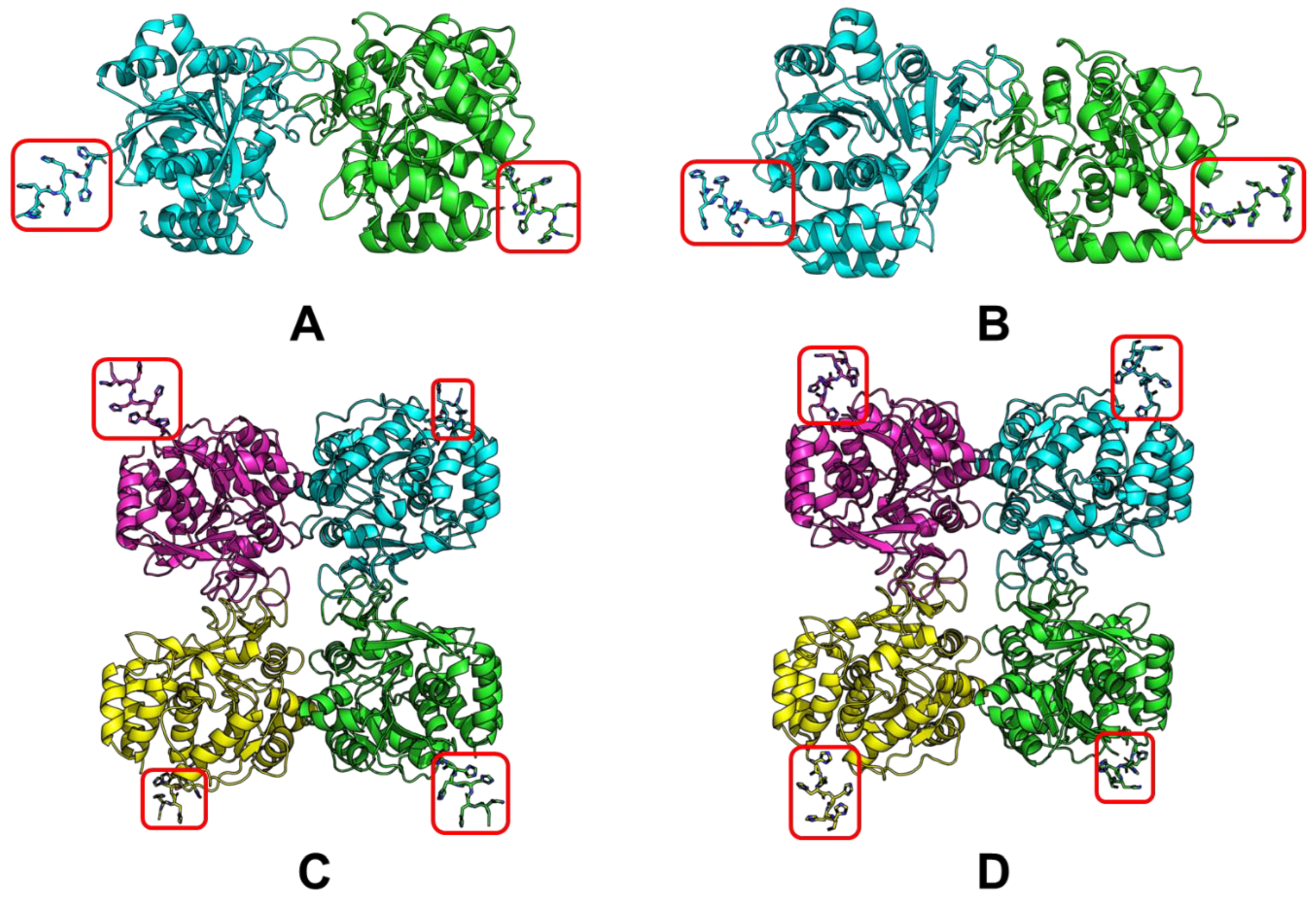

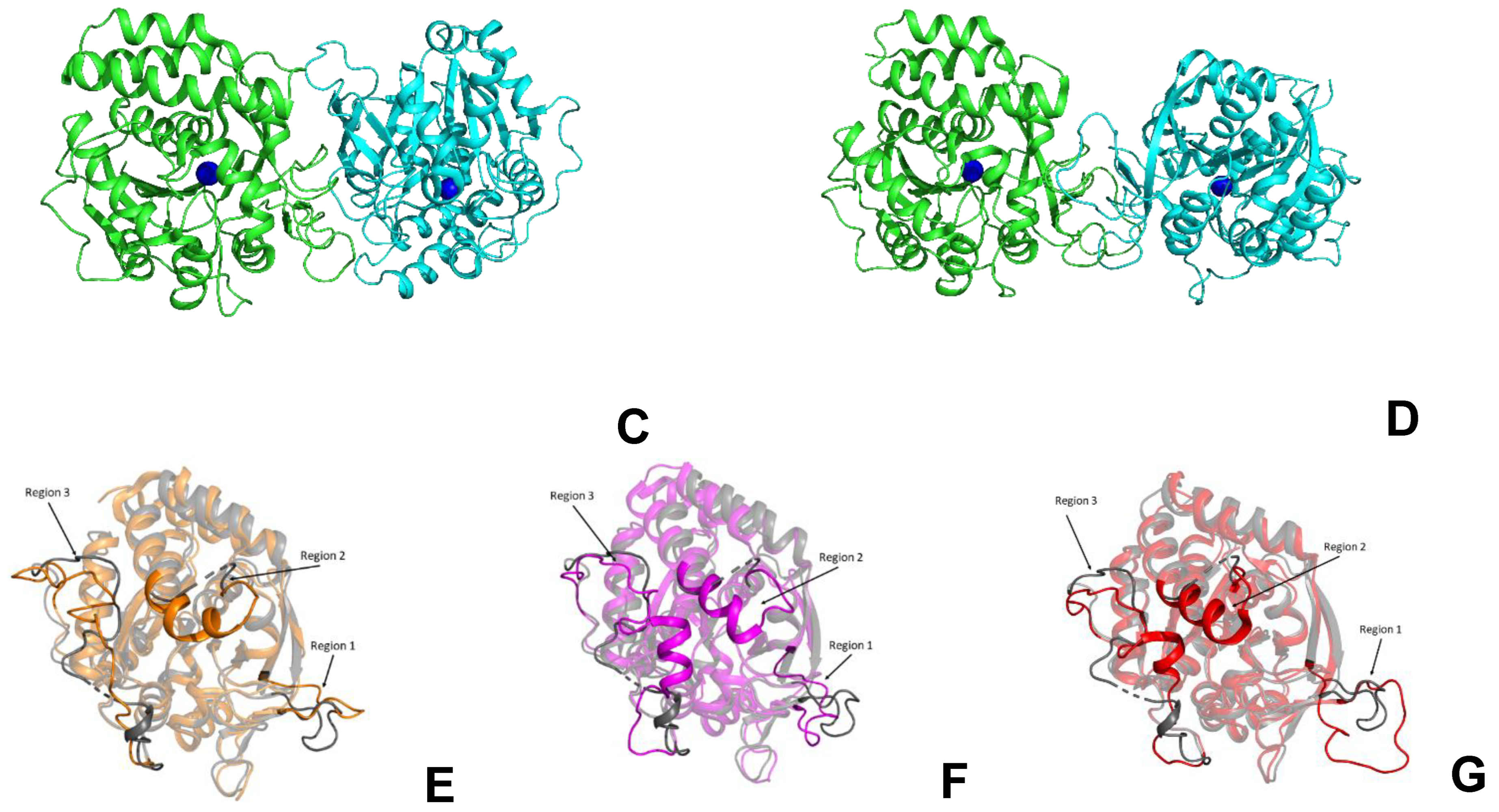
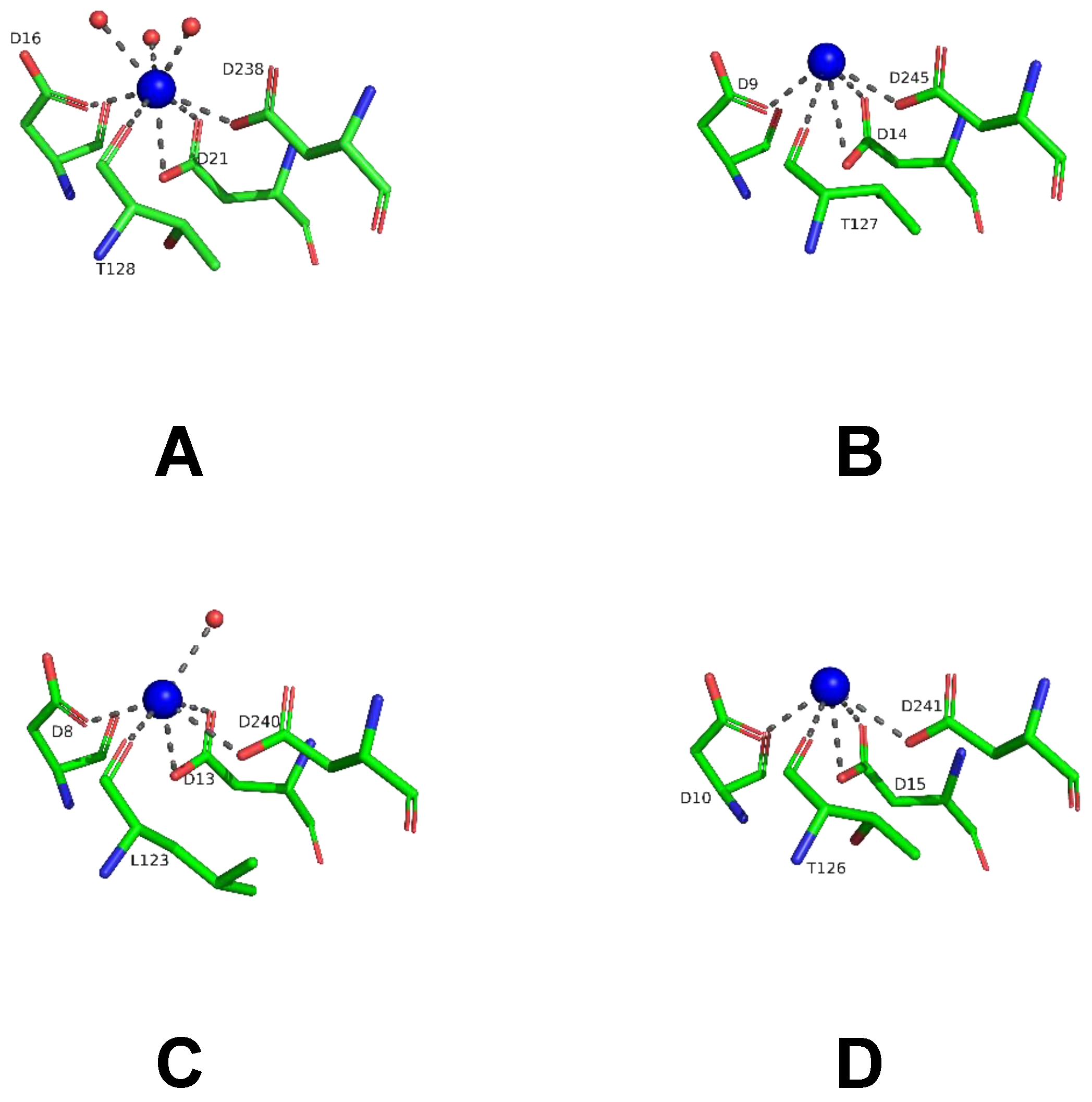
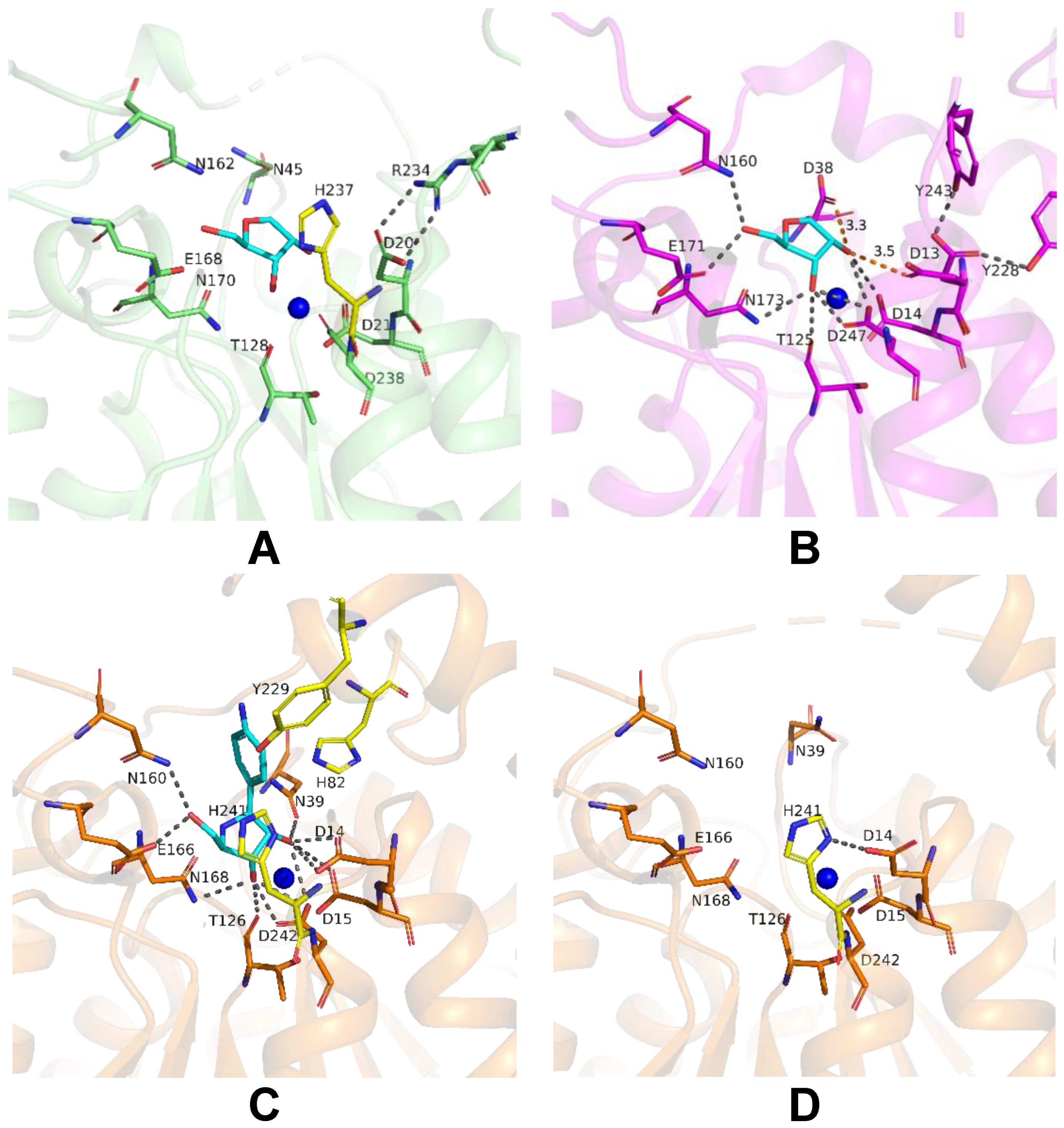
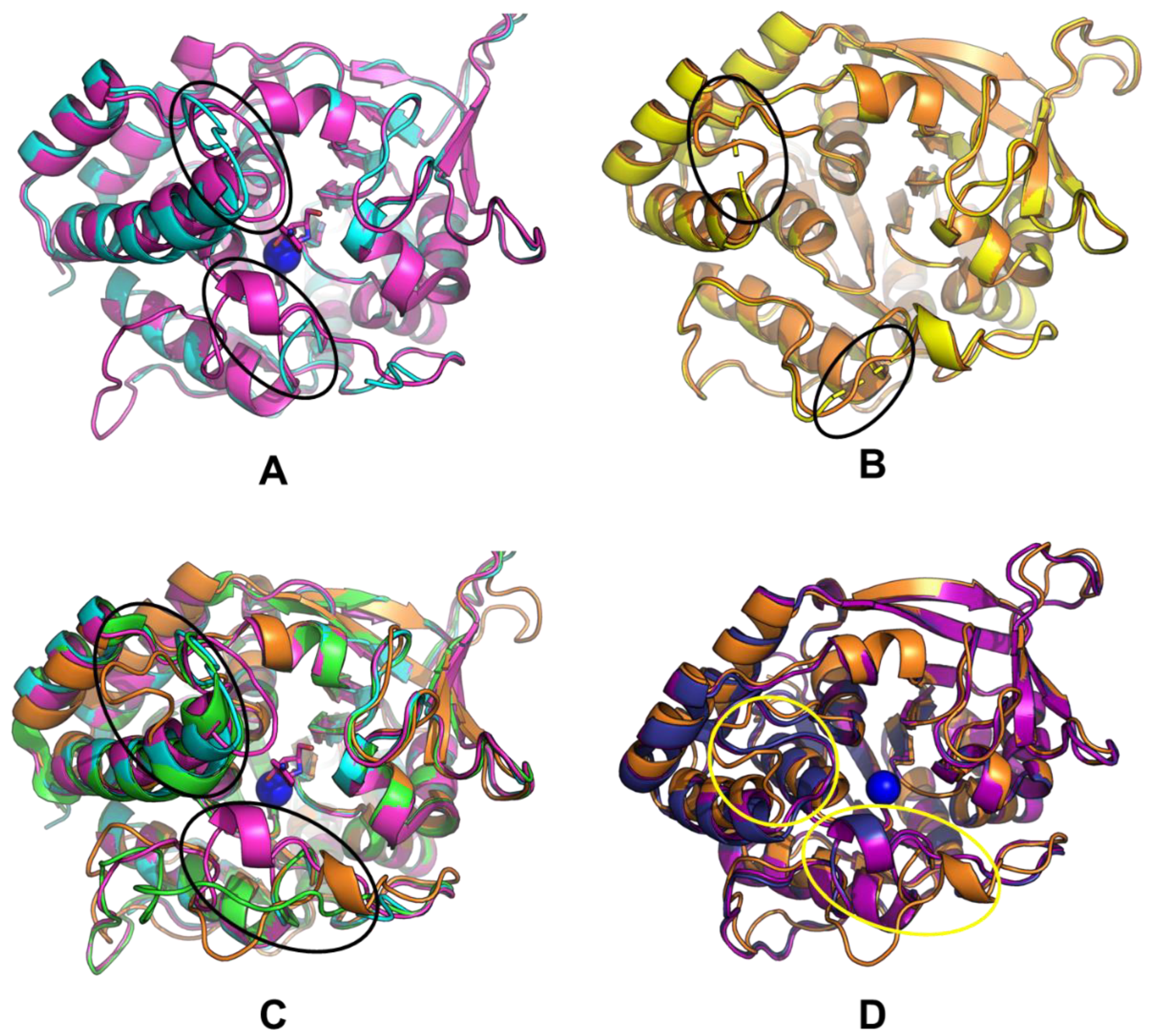
| Organism Type | Organism | PDB Code | Resolution | RMSD, Å | Sequence Identity, % | Percentage of the Aligned Residues, % |
|---|---|---|---|---|---|---|
| Bacteria * | Limosilactobacillus reuteri LR1(LreRihC) * | 8QND | 1.9 | - | - | - |
| Bacteria | Bacillus anthracis (BanRihC) | 2C40 (holo form) | 2.2 Å | 1.23 | 24 | 87 |
| Bacteria | Gardnerella vaginalis 315–A (GvaRihC) | 6BA1 (apo form) | 2.9 Å | 1.43 | 32 | 87 |
| Plants | Physcomitrella patens (PpaRihC) | 4KPN (apo form) | 3.35 Å | 1.17 | 32 | 83 |
| Plants | Zea mays (ZmaRihC) | 4KPO (apo form) | 2.49 Å | 1.12 | 33 | 87 |
| Protozoa | Crithidia fasciculate (CfaRihC) | 1MAS (apo form) | 2.5 Å | 1.02 | 34 | 91 |
| 2MAS (holo form) | 2.3 Å | 1.06 | 34 | 91 | ||
| Protozoa | Leishmania braziliensis (LbrRihC) | 5TSQ (holo form) | 1.53 Å | 1.02 | 33 | 87 |
| Protozoa | Leishmania major (LmaRihC) | 1EZR (apo porm) | 2.5 Å | 1.04 | 34 | 91 |
| Organism Type | Organism Name | Shortened RihC Name |
|---|---|---|
| Bacteria | Bacillus anthracis | BanRihC |
| Bacteria | Escherichia coli | EcoRihC |
| Bacteria | Gardnerella vaginalis 315-A | GvaRihC |
| Bacteria | Limosilactobacillus reuteri LR1 | LreRihC |
| Bacteria | Salmonella enterica | SenRihC |
| Plants | Physcomitrella patens | PpaRihC |
| Plants | Zea mays | ZmaRihC |
| Protozoa | Crithidia fasciculata | CfaRihC |
| Protozoa | Leishmania braziliensis | LbrRihC |
| Protozoa | Leishmania major | LmaRihC |
| Diffraction Source | Institute of Organic Chemistry RAS (Rigaku OD XtaLAB Synergy–S) |
|---|---|
| Wavelength (Å) | 1.54 |
| Temperature (K) | 100 |
| Detector | HyPix–6000HE |
| Crystal–to–detector distance (mm) | 32.0 |
| Rotation range per image (°) | 0.35 |
| Total rotation range (°) | 344.75 |
| Space group | P1211 |
| a, b, c (Å) | 84.11, 81.53, 86.85 |
| α, β, γ (°) | 90.0, 95.8, 90.0 |
| Average mosaicity (°) | 0.27 |
| Resolution range (Å) | 63.38–1.90 (1.93–1.90) |
| Completeness (%) | 99.8 (100) |
| Average redundancy | 6.4 (6.6) |
| I/σ(I) | 19.5 (5.5) |
| Rmeas (%) | 9.1 (37.7) |
| CC1/2 | 91.4 (95.6) |
| Rfact (%) | 16.9 |
| Rfree. (%) | 20.1 |
| RMSD Bonds (Å) | 0.01 |
| RMSD Angles (°) | 1.85 |
| Ramachandran favored (%) | 96.8 |
| Ramachandran allowed (%) | 3.1 |
| PDB entry code | 8QND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaposhnikov, L.A.; Chikurova, N.Y.; Atroshenko, D.L.; Savin, S.S.; Kleymenov, S.Y.; Chernobrovkina, A.V.; Pometun, E.V.; Minyaev, M.E.; Matyuta, I.O.; Hushpulian, D.M.; et al. Structure–Functional Examination of Novel Ribonucleoside Hydrolase C (RihC) from Limosilactobacillus reuteri LR1. Int. J. Mol. Sci. 2024, 25, 538. https://doi.org/10.3390/ijms25010538
Shaposhnikov LA, Chikurova NY, Atroshenko DL, Savin SS, Kleymenov SY, Chernobrovkina AV, Pometun EV, Minyaev ME, Matyuta IO, Hushpulian DM, et al. Structure–Functional Examination of Novel Ribonucleoside Hydrolase C (RihC) from Limosilactobacillus reuteri LR1. International Journal of Molecular Sciences. 2024; 25(1):538. https://doi.org/10.3390/ijms25010538
Chicago/Turabian StyleShaposhnikov, Leonid A., Natalia Yu. Chikurova, Denis L. Atroshenko, Svyatoslav S. Savin, Sergei Yu. Kleymenov, Alla V. Chernobrovkina, Evgenii V. Pometun, Mikhail E. Minyaev, Ilya O. Matyuta, Dmitry M. Hushpulian, and et al. 2024. "Structure–Functional Examination of Novel Ribonucleoside Hydrolase C (RihC) from Limosilactobacillus reuteri LR1" International Journal of Molecular Sciences 25, no. 1: 538. https://doi.org/10.3390/ijms25010538
APA StyleShaposhnikov, L. A., Chikurova, N. Y., Atroshenko, D. L., Savin, S. S., Kleymenov, S. Y., Chernobrovkina, A. V., Pometun, E. V., Minyaev, M. E., Matyuta, I. O., Hushpulian, D. M., Boyko, K. M., Tishkov, V. I., & Pometun, A. A. (2024). Structure–Functional Examination of Novel Ribonucleoside Hydrolase C (RihC) from Limosilactobacillus reuteri LR1. International Journal of Molecular Sciences, 25(1), 538. https://doi.org/10.3390/ijms25010538