
Modulation of the Immune Response to Allergies Using Alternative Functional Foods

 ,
,
Abstract
:1. Introduction
2. Allergic Diseases
Immunological Mechanism of Allergic Reactions
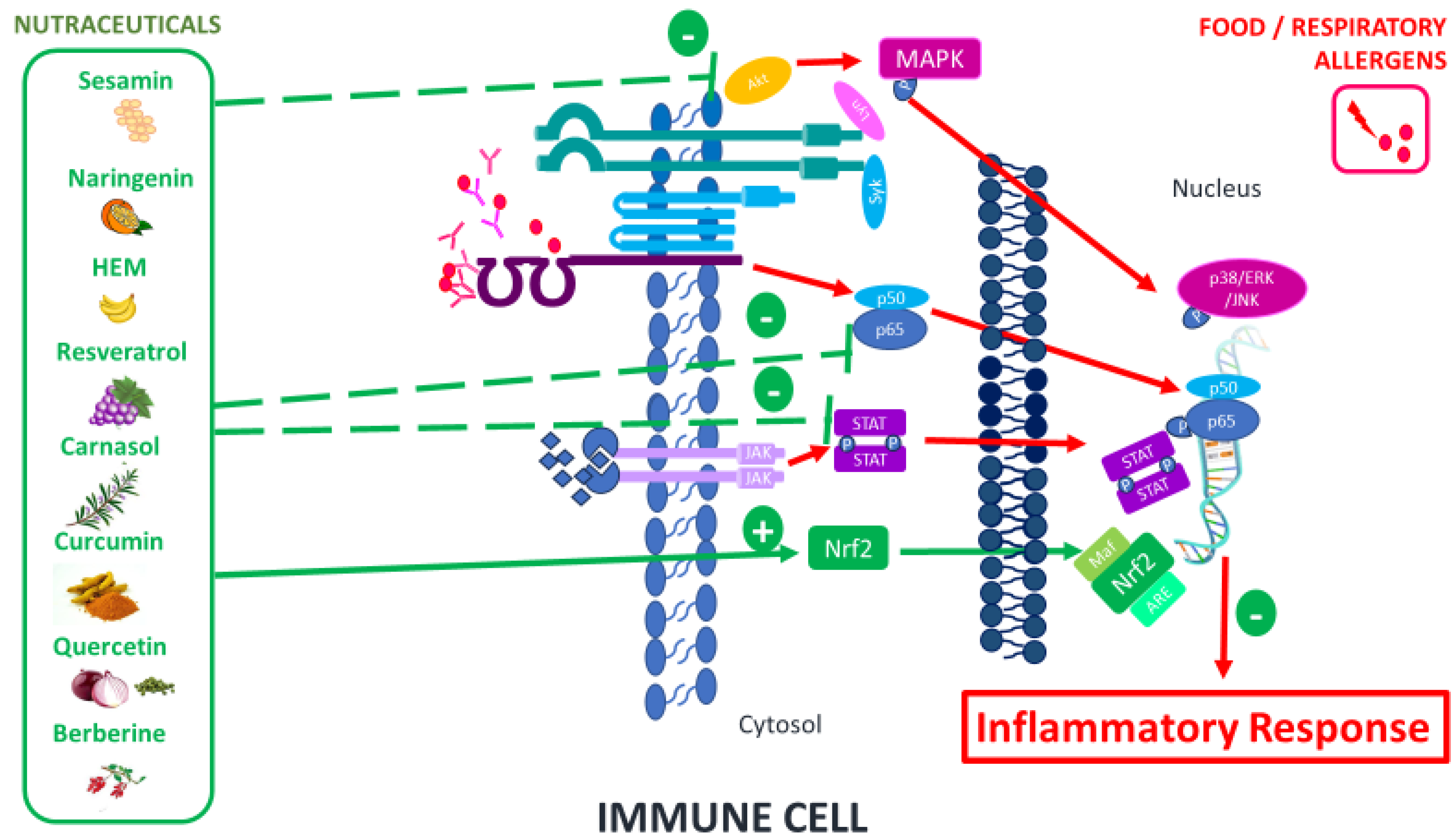
3. Functional Foods (Nutraceuticals)
Therapeutic Nutraceuticals and Their Immunomodulatory Mechanisms

{kind=link}
| Nutraceuticals | Study Design | Immunomodulatory Effect | Refs. |
|---|---|---|---|
| Sesamin | Asthma murine model | ↓ NFκB and ↓ (IL-4, IL-5, and IL-13) ↓ IgE | [21] |
| Naringenin | Asthma/Inflammatory murine model | ↓ NFκB and ↓ (IL-4 and IL-13) | [22] |
| ↓ MDA, ↓ (IL-4 and IL-13) in lung tissues, and ↓ total eosinophils ↑ GSH | [23] | ||
| C3G | Cutaneous reaction in murine model | ↓ β-Hex and ↓ histamine release IgE-mediated. ↓ Mast cell degranulation | [24] |
| MPI | Murine model, combining AR and asthma and human M1 macrophages | ↑ Th1 profile ↓ Type 2 immune cells ↓ NFκB pathway ↓ CD86 and ↓ HLA-DR | [25] |
| Resveratrol | Murine model | ↓ Mast cells degranulation with ↓ β-Hex by ↓ PI 4-kinase ↓ IgE-mediated histamine release | [10] |
| Human mast cells from AR patients | ↓ IgE- mediated β-Hex release ↓ pSTAT3 and pERK1/2 ↓ Nasal symptoms in AR patients with ↓ (IgE, IL-4, and TNF-α) ↓ Eosinophils in blood | ||
| Human mast cells from AR patients | ↓ PGD2 ↑ TNF in mast cells after IgE-dependent activation | [26] | |
| Rat basophilic leukemia-2H3 cells and AR murine model | ↓ β-Hex ↓ IgE-mediated histamine ↓ (DCs and B and mast cells) ↓ TXNIP pathway ↓ (PGD2, LTC4, ECP, IL-4, IL-5, IL-6, IL-33, and TNF) | [27] | |
| Asthma murine model | ↑ FOXP3+ cells ↓ miR-34a in lung infiltrating cells | [28] | |
| Carnosol | Asthma murine model | ↓ Mast cells degranulation ↓ Eosinophils ↓ (IL-4 and IL-13) | [29] |
| Allergic inflammation model | ↓ (Syk, Akt phosphorylation, and NFκB). ↓ Intracellular Ca2+ mobilization ↓ ROS production and β-Hex release ↓ Mast cells degranulation, and ↓ (IL-6, TNF, IL-13, CCL1, CCL2, CCL3, and CCL9) | [30] | |
| Curcumin | Allergic inflammation model | ↓ Mast cell activation and ↓ NFκB signaling pathway | [31] |
| Piperine | Asthma and AR murine model | ↓ Eosinophil infiltration ↓ Th2 response and ↑ Th1 cells ↓ STAT3 and NFκB signaling pathways | [32,33] |
| Quercetin | Human HMC-1 mast cells from AR patients | ↓ (Histamine, TNF-α, IL-1β, IL-6, and IL-8) and Ca2+ ↓ NFκB signaling pathway ↓ Allergic symptoms in AR patients | [34] |
| Baicalein | AR and FA murine model | ↓ (Histamine, OVA-IgE, IL-1β, IL-6, IL-8, and TNF) ↑ Treg in a mouse | [36] |
| Human HMC-1 mast cells | ↓ (Histamine and β-hex) ↓ JAK2-STAT5 and NF-kB signaling pathways | [37] | |
| Berberine | House dust mite AR murine model | ↓ (sIgE, GATA-3, and T-bet mRNA levels, L-10) ↓ Eosinophil infiltration and ↑Treg | [38] |
| Cutaneous anaphylaxis murine model | ↓ Mast cell activation by ↓ (β-hex, histamine, IL-4 and TNF) ↓ MAPK signaling pathway | [39] | |
| PUFAs | Transgenic Fat-1 murine model (allergic airway) | ↓ Th2-cell response | [41] |
| AR animal model | ↓ degranulation mast cells | [42] | |
| SCFAs | Keratinocyte humans | Protection and reprogramming of skin barrier function and metabolism | [43] |
| Pulmonary inflammation murine model | ↓ Eosinophils ↓ Th2 (IL-4 and IL13) | [45] | |
| Fiber (RS) | In vitro model: HEK-Blue™ TLR cells and macrophage cells RAW264.7 | ↑ Modulation of immune cells and chemokine secretion via CLRs or TLRs | [49] |
| ASX | Asthma murine model | ↓ Th1/Th2 cells ↓ (IL-4 and IL-5) and ↑ IFN-γ) and ↓ IgE | [50] |
| SFN | Asthma patients | ↑ Bronchoprotective response ↓ Nrf2 signaling pathway | [52] |
| Asthma animal model | ↑ antioxidant effects ↓ Th17 responses | [53] | |
| mRNA ↓ (TNF-α, SMAD2, IL6, IL-1β, IL-8, and MIP-1β) expression | [54] |
4. Nutraceuticals and Allergic Rhinitis (AR)
| Nutraceuticals | Refs. | Source of Nutraceuticals | Phase of Allergic Reaction | Achievements | Limitations |
|---|---|---|---|---|---|
| Quercetin | [62,63,64,65] | Isolated from a natural source. Commercially obtained | In vitro models under specific IL-14 stimulation. Onset phase in animal model, after sensitization phase. Clinical trial in AR patients in the sensitization phase. | Suppressive effect on NO production from nasal epithelial cells. Increase of TRX production in nasal epithelial cells and animal models. Significant decrease in allergy nasal symptoms (seizing and rubbing). | Lack of food matrix Low bioavailability Lack of research on dosage and forms of administration Limited research in humans and clinical trials Lack of studies of possible interactions with other drugs |
| Sulforaphane | [66] | Broccoli sprout extract, including sulphoraphane. | Clinical trial in AR patients in the sensitization phase. | Anti-inflammatory and anti-allergic properties in AR patients, with the improvement of nasal symptoms. | |
| Resveratrol | [27,28] | Isolated from natural sources (for example, Abies georgei). | Onset phase in animal model, after sensitization phase. | Reduction of nasal symptoms. Anti-allergic and anti-inflammatory properties. | |
| Piper | [69] | Piper nigrum extract. | Reduction of nasal symptoms and inflammatory mediators. | ||
| Baicalin | [36] | Isolated from natural sources. Commercially obtained. | Anti-allergic response in OVA-induced AR pigs and in LPS-stimulated human mast cells. | ||
| Berberine | [38] | Isolated from natural sources. Commercially obtained. | Reduction of the inflammatory and increase in regulatory response. | ||
| Omega-3 fatty acids | [67] | Dietary fatty acids on allergic models. | Before the sensitization phase in animal models. | Suppression of the allergic symptoms. | |
| Fiber | [68] | Natural formulation from Hordeum vulgare. | Clinical trial in AR patients in the sensitization phase. | All symptoms of AR except cough were significantly reduced. |
5. Nutraceuticals and Food Allergy (FA)
| Nutraceuticals | Refs. | Source of Nutraceuticals | Phase of Allergic Reaction | Achievements | Limitations |
|---|---|---|---|---|---|
| Naringenin and Baicalein | [69] | Isolated from a natural source. Commercially obtained. | In vitro models. | Reduction of basophils degranulation and improvement of the intestinal epithelial barrier. | Lack of research on dosage and forms of administration. Limited research in humans and clinical trials. Variability in the results obtained in the models studied. They do not replace food allergen avoidance. |
| C3G | [71] | Commercially obtained. | Onset phase in animal model, after sensitization phase. | Improve the clinical FA symptoms and regulate the T cell phenotype. | |
| Resveratrol | [27] | Isolated from natural sources (for example, Abies georgei). | Attenuation of allergic responses in anaphylactic models. | ||
| BDMC | [72] | Commercially obtained. | Attenuation of OVA-induced FA inhibiting the MAPK and NF-κB signaling pathways. | ||
| Quercetin | [73,74] | Commercially obtained and specific conjugation. | In vitro model. Before the sensitization phase in animal model. | Regulation of IgE-mediated mast cell responses and protection in front of effects during FA. Iron–quercetin complex confers resilience in cow’s milk allergy. | |
| Pipper | [75] | Piper nigrum extract. | Onset phase in animal model, after sensitization phase. | FA attenuation, decreasing Th2 cell response, and regulating the Th17/Treg balance. | |
| Berberine | [76,77] | Commercially obtained, and specific conjugation. | Onset phase in animal model, after sensitization phase. | Combination with OIT induces tolerance to peanuts. | |
| PUFAs | [80,81,83,84] | Omega-3 supplementation. | FA patients in the sensitization phase. | Prevention of FA in children and reduction in adverse effects in fish allergy. | |
| Fiber and SCFAs | [85,86,88] | Modification to the AIN93G control diet. Commercially obtained (SCFAs). | Onset phase in animal model, after sensitization phase. Children with non-IgE-mediated CMA. | Induction of the tolerance and protection in FA. Regulation mast cells degranulation (via epigenetic). Regulation of non-IgE mediated CMA in children. |
6. Nutraceuticals and Atopic Dermatitis (AD)
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AA | Allergic asthma |
| AD | Atopic dermatitis |
| AR | Allergic rhinitis |
| ASX | Astaxanthin |
| ALA | Alpha-linolenic acid |
| AIT | Allergen immunotherapy |
| BALF | Bronchoalveolar lavage fluid |
| β-Hex | β-hexosaminidase |
| BMMCs | Bone marrow-derived mast cells |
| BDMC | Bisdemethoxycurcumin |
| CRS | Chronic rhinosinusitis |
| CRSwNP | Chronic rhinosinusitis without nasal polyposis |
| C3G | Cyanidin-3-O-β-glucoside |
| CA | Carnosic acid |
| COX-2 | Cycloocxygenase |
| DA | Drug allergy |
| DCs | Dendritic cells |
| EPA | Eicosapentaenoic acid |
| FA | Food allergy |
| GM-CSF | Granulocyte- monocyte colony-stimulating factor. |
| GPR43- FFA2 | Free fatty acid receptor 2 |
| TXNIP | Thioredoxin-interacting protein. |
| GSH | Highly increased glutation |
| HNEpC | Human nasal epithelial cells |
| iNOS | Nitric oxide synthase |
| IL | Interleukin |
| Ig | Immunoglobulin |
| IgE | Inmunoglobulin E |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen-activated protein kinases |
| MDA | Malondialdehyde |
| MPI | Musa paradisiaca inflorescence |
| NO | Nitric oxide. |
| Nrf2 | Nuclear factor Erythroid 2- related factor 2 |
| NFκB | Kappa-light-chain-enhancer |
| NLF | Nasal lavage fluid |
| PA | Phthalic anhydride |
| PI | Phosphatidylinositol |
| PGD | Prostaglandin D2 |
| PNE | Piper nigrum extract |
| PCA | Passive cutaneous anaphylaxis |
| PUFA | Polyunsaturated fatty acids |
| PNIF | Peak nasal inspiratory flow |
| OVA | Ovalbumin |
| ROS | Allergen-induced reactive oxygen species |
| RS | Resistant starch |
| SCFAs | Short-chain fatty acids |
| SFN | Sulforaphane |
| TCI | Topical calcineurin inhibitors |
| TRX | Thioredoxin |
| Th2 | T helper type 2 cells |
| TNSS | aNsal symptoms |
| 15-HEPE | 15-hydroxyeicosapentaenoic acid |
References
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology, 10th ed.; Institute of Immunology, School of Life Sciences, University of Science and Technology of China: Hefei, China, 2022. [Google Scholar]
- Shin, Y.H.; Hwang, J.; Kwon, R.; Lee, S.W.; Kim, M.S.; GBD 2019 Allergic Disorders Collaborators; Shin, J.I.; Yon, D.K. Global, regional, and national burden of allergic disorders and their risk factors in 204 countries and territories, from 1990 to 2019: A systematic analysis for the Global Burden of Disease Study 2019. Allergy 2023, 78, 2232–2254. [Google Scholar] [CrossRef] [PubMed]
- Samitas, K.; Carter, A.; Kariyawasam, H.H.; Xanthou, G. Upper and lower airway remodelling mechanisms in asthma, allergic rhinitis and chronic rhinosinusitis: The one airway concept revisited. Allergy 2018, 73, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Fokkens, W.J.; Lund, V.J.; Hopkins, C.; Hellings, P.W.; Kern, R.; Reitsma, S.; Toppila-Salmi, S.; Bernal-Sprekelsen, M.; Mullol, J.; Alobid, I.; et al. European Position Paper on Rhinosinusitis and Nasal Polyps 2020. Rhinology 2020, 58 (Suppl. S29), 1–464. [Google Scholar] [CrossRef] [PubMed]
- Golden, D.B. Anaphylaxis to insect stings. Immunol. Allergy Clin. N. Am. 2015, 35, 287–302. [Google Scholar] [CrossRef]
- Breiteneder, H.; Peng, Y.Q.; Agache, I.; Diamant, Z.; Eiwegger, T.; Fokkens, W.J.; Traidl-Hoffmann, C.; Nadeau, K.; O’Hehir, R.E.; O’Mahony, L.; et al. Biomarkers for diagnosis and prediction of therapy responses in allergic diseases and asthma. Allergy 2020, 75, 3039–3068. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, S.E.; Guidos-Fogelbach, G.; Annesi-Maesano, I.; Pawankar, R.; Amato, G.D.; Latour-Staffeld, P.; Urrutia-Pereira, M.; Kesic, M.J.; Hernandez, M.L. Climate change and global issues in allergy and immunology. J. Allergy Clin. Immunol. 2021, 148, 1366–1377. [Google Scholar] [CrossRef]
- Palomares, O.; Akdis, M.; Martin-Fontecha, M.; Akdis, C.A. Mechanisms of immune regulation in allergic diseases: The role of regulatory T and B cells. Immunol. Rev. 2017, 278, 219–236. [Google Scholar] [CrossRef]
- Brasal-Prieto, M.; Fernandez-Prades, L.; Dakhaoui, H.; Sobrino, F.; Lopez-Enriquez, S.; Palomares, F. Update on In Vitro Diagnostic Tools and Treatments for Food Allergies. Nutrients 2023, 15, 3744. [Google Scholar] [CrossRef]
- Civelek, M.; Bilotta, S.; Lorentz, A. Resveratrol Attenuates Mast Cell Mediated Allergic Reactions: Potential for Use as a Nutraceutical in Allergic Diseases? Mol. Nutr. Food Res. 2022, 66, e2200170. [Google Scholar] [CrossRef]
- Alba, G.; Dakhaoui, H.; Santa-Maria, C.; Palomares, F.; Cejudo-Guillen, M.; Geniz, I.; Sobrino, F.; Montserrat-de la Paz, S.; Lopez-Enriquez, S. Nutraceuticals as Potential Therapeutic Modulators in Immunometabolism. Nutrients 2023, 15, 411. [Google Scholar] [CrossRef]
- Vranceanu, M.; Galimberti, D.; Banc, R.; Dragos, O.; Cozma-Petrut, A.; Heghes, S.C.; Vostinaru, O.; Cuciureanu, M.; Stroia, C.M.; Miere, D.; et al. The Anticancer Potential of Plant-Derived Nutraceuticals via the Modulation of Gene Expression. Plants 2022, 11, 2524. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Basketter, D.A.; McFadden, J.P.; Kimber, I. Assessing the severity of allergic reactions: A regulatory dilemma. Contact Dermat. 2012, 67, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, Y.; Zhang, H.; Hu, L.; Liu, J.; Wang, L.; Wang, T.; Zhang, H.; Cong, L.; Wang, Q. Pathogenesis of allergic diseases and implications for therapeutic interventions. Signal Transduct. Target. Ther. 2023, 8, 138. [Google Scholar] [CrossRef] [PubMed]
- Rajan, T.V. The Gell-Coombs classification of hypersensitivity reactions: A re-interpretation. Trends Immunol. 2003, 24, 376–379. [Google Scholar] [CrossRef]
- Johansson, S.G.; Bieber, T.; Dahl, R.; Friedmann, P.S.; Lanier, B.Q.; Lockey, R.F.; Motala, C.; Ortega Martell, J.A.; Platts-Mills, T.A.; Ring, J.; et al. Revised nomenclature for allergy for global use: Report of the Nomenclature Review Committee of the World Allergy Organization, October 2003. J. Allergy Clin. Immunol. 2004, 113, 832–836. [Google Scholar] [CrossRef]
- Dispenza, M.C. Classification of hypersensitivity reactions. Allergy Asthma Proc. 2019, 40, 470–473. [Google Scholar] [CrossRef]
- Lerch, M.; Pichler, W.J. The immunological and clinical spectrum of delayed drug-induced exanthems. Curr. Opin. Allergy Clin. Immunol. 2004, 4, 411–419. [Google Scholar] [CrossRef]
- McCarty, M.F.; Lerner, A.; DiNicolantonio, J.J.; Benzvi, C. Nutraceutical Aid for Allergies-Strategies for Down-Regulating Mast Cell Degranulation. J. Asthma Allergy 2021, 14, 1257–1266. [Google Scholar] [CrossRef]
- Lin, C.H.; Shen, M.L.; Zhou, N.; Lee, C.C.; Kao, S.T.; Wu, D.C. Protective effects of the polyphenol sesamin on allergen-induced T(H)2 responses and airway inflammation in mice. PLoS ONE 2014, 9, e96091. [Google Scholar] [CrossRef]
- Shi, Y.; Dai, J.; Liu, H.; Li, R.R.; Sun, P.L.; Du, Q.; Pang, L.L.; Chen, Z.; Yin, K.S. Naringenin inhibits allergen-induced airway inflammation and airway responsiveness and inhibits NF-kappaB activity in a murine model of asthma. Can. J. Physiol. Pharmacol. 2009, 87, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Jasemi, S.V.; Khazaei, H.; Fakhri, S.; Mohammadi-Noori, E.; Farzaei, M.H. Naringenin Improves Ovalbumin-Induced Allergic Asthma in Rats through Antioxidant and Anti-Inflammatory Effects. Evid. Based Complement. Altern. Med. 2022, 2022, 9110798. [Google Scholar] [CrossRef] [PubMed]
- Hiemori-Kondo, M.; Morikawa, E.; Fujikura, M.; Nagayasu, A.; Maekawa, Y. Inhibitory effects of cyanidin-3-O-glucoside in black soybean hull extract on RBL-2H3 cells degranulation and passive cutaneous anaphylaxis reaction in mice. Int. Immunopharmacol. 2021, 94, 107394. [Google Scholar] [CrossRef] [PubMed]
- Gadelha, F.A.A.F.; Cavalcanti, R.F.P.; Vieira, G.C.; Ferreira, L.K.D.P.; de Sousa, G.R.; Filho, J.M.B.; Barbosa, M.A.; dos Santos, S.G.; Piuvezam, M.R. Immunomodulatory properties of Musa paradisiaca L. inflorescence in Combined Allergic Rhinitis and Asthma Syndrome (CARAS) model towards NFκB pathway inhibition. J. Funct. Foods 2021, 83, 104540. [Google Scholar] [CrossRef]
- Shirley, D.; McHale, C.; Gomez, G. Resveratrol preferentially inhibits IgE-dependent PGD2 biosynthesis but enhances TNF production from human skin mast cells. Biochim. Biophys. Acta 2016, 1860, 678–685. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Liu, Q.M.; Gao, Y.Y.; Liu, B.; Liu, H.; Cao, M.J.; Yang, X.W.; Liu, G.M. Attenuation of allergic responses following treatment with resveratrol in anaphylactic models and IgE-mediated mast cells. Food Funct. 2019, 10, 2030–2039. [Google Scholar] [CrossRef]
- Alharris, E.; Alghetaa, H.; Seth, R.; Chatterjee, S.; Singh, N.P.; Nagarkatti, M.; Nagarkatti, P. Resveratrol Attenuates Allergic Asthma and Associated Inflammation in the Lungs Through Regulation of miRNA-34a That Targets FoxP3 in Mice. Front. Immunol. 2018, 9, 2992. [Google Scholar] [CrossRef]
- Lee, J.E.; Im, D.S. Suppressive Effect of Carnosol on Ovalbumin-Induced Allergic Asthma. Biomol. Ther. 2021, 29, 58–63. [Google Scholar] [CrossRef]
- Crozier, R.W.E.; Yousef, M.; Coish, J.M.; Fajardo, V.A.; Tsiani, E.; MacNeil, A.J. Carnosic acid inhibits secretion of allergic inflammatory mediators in IgE-activated mast cells via direct regulation of Syk activation. J. Biol. Chem. 2023, 299, 102867. [Google Scholar] [CrossRef]
- Kinney, S.R.; Carlson, L.; Ser-Dolansky, J.; Thompson, C.; Shah, S.; Gambrah, A.; Xing, W.; Schneider, S.S.; Mathias, C.B. Curcumin Ingestion Inhibits Mastocytosis and Suppresses Intestinal Anaphylaxis in a Murine Model of Food Allergy. PLoS ONE 2015, 10, e0132467. [Google Scholar] [CrossRef]
- Kim, S.H.; Lee, Y.C. Piperine inhibits eosinophil infiltration and airway hyperresponsiveness by suppressing T cell activity and Th2 cytokine production in the ovalbumin-induced asthma model. J. Pharm. Pharmacol. 2009, 61, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.T.; Fan, Y.; Piao, C.H.; Nguyen, T.V.; Shin, D.U.; Jung, S.Y.; Hyeon, E.; Song, C.H.; Lee, S.Y.; Shin, H.S.; et al. Piper Nigrum extract improves OVA-induced nasal epithelial barrier dysfunction via activating Nrf2/HO-1 signaling. Cell. Immunol. 2020, 351, 104035. [Google Scholar] [CrossRef] [PubMed]
- Park, H.H.; Lee, S.; Son, H.Y.; Park, S.B.; Kim, M.S.; Choi, E.J.; Singh, T.S.; Ha, J.H.; Lee, M.G.; Kim, J.E.; et al. Flavonoids inhibit histamine release and expression of proinflammatory cytokines in mast cells. Arch. Pharm. Res. 2008, 31, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Thornhill, S.M.; Kelly, A.M. Natural treatment of perennial allergic rhinitis. Altern. Med. Rev. 2000, 5, 448–454. [Google Scholar] [PubMed]
- Zhou, Y.J.; Wang, H.; Sui, H.H.; Li, L.; Zhou, C.L.; Huang, J.J. Inhibitory effect of baicalin on allergic response in ovalbumin-induced allergic rhinitis guinea pigs and lipopolysaccharide-stimulated human mast cells. Inflamm. Res. 2016, 65, 603–612. [Google Scholar] [CrossRef]
- Bae, M.J.; Shin, H.S.; See, H.J.; Jung, S.Y.; Kwon, D.A.; Shon, D.H. Baicalein induces CD4(+)Foxp3(+) T cells and enhances intestinal barrier function in a mouse model of food allergy. Sci. Rep. 2016, 6, 32225. [Google Scholar] [CrossRef]
- Kim, B.Y.; Park, H.R.; Jeong, H.G.; Kim, S.W. Berberine reduce allergic inflammation in a house dust mite allergic rhinitis mouse model. Rhinology 2015, 53, 353–358. [Google Scholar] [CrossRef]
- Fu, S.; Ni, S.; Wang, D.; Fu, M.; Hong, T. Berberine suppresses mast cell-mediated allergic responses via regulating FcvarepsilonRI-mediated and MAPK signaling. Int. Immunopharmacol. 2019, 71, 1–6. [Google Scholar] [CrossRef]
- Miyata, J.; Arita, M. Role of omega-3 fatty acids and their metabolites in asthma and allergic diseases. Allergol. Int. 2015, 64, 27–34. [Google Scholar] [CrossRef]
- Bilal, S.; Haworth, O.; Wu, L.; Weylandt, K.H.; Levy, B.D.; Kang, J.X. Fat-1 transgenic mice with elevated omega-3 fatty acids are protected from allergic airway responses. Biochim. Biophys. Acta 2011, 1812, 1164–1169. [Google Scholar] [CrossRef]
- Sawane, K.; Nagatake, T.; Hosomi, K.; Hirata, S.I.; Adachi, J.; Abe, Y.; Isoyama, J.; Suzuki, H.; Matsunaga, A.; Fukumitsu, S.; et al. Dietary Omega-3 Fatty Acid Dampens Allergic Rhinitis via Eosinophilic Production of the Anti-Allergic Lipid Mediator 15-Hydroxyeicosapentaenoic Acid in Mice. Nutrients 2019, 11, 2868. [Google Scholar] [CrossRef] [PubMed]
- Trompette, A.; Pernot, J.; Perdijk, O.; Alqahtani, R.A.A.; Domingo, J.S.; Camacho-Munoz, D.; Wong, N.C.; Kendall, A.C.; Wiederkehr, A.; Nicod, L.P.; et al. Gut-derived short-chain fatty acids modulate skin barrier integrity by promoting keratinocyte metabolism and differentiation. Mucosal Immunol. 2022, 15, 908–926. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, C.; Zhang, K.; Xue, W. The role of gut microbiota and its metabolites short-chain fatty acids in food allergy. Food Sci. Hum. Wellness 2023, 12, 702–710. [Google Scholar] [CrossRef]
- Theiler, A.; Barnthaler, T.; Platzer, W.; Richtig, G.; Peinhaupt, M.; Rittchen, S.; Kargl, J.; Ulven, T.; Marsh, L.M.; Marsche, G.; et al. Butyrate ameliorates allergic airway inflammation by limiting eosinophil trafficking and survival. J. Allergy Clin. Immunol. 2019, 144, 764–776. [Google Scholar] [CrossRef] [PubMed]
- Sobh, M.; Montroy, J.; Daham, Z.; Sibbald, S.; Lalu, M.; Stintzi, A.; Mack, D.; Fergusson, D.A. Tolerability and SCFA production after resistant starch supplementation in humans: A systematic review of randomized controlled studies. Am. J. Clin. Nutr. 2022, 115, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, J.; Song, Y.H.; Zhao, R.; Xia, L.; Chen, Y.; Cui, Y.P.; Rao, Z.Y.; Zhou, Y.; Zhuang, W.; et al. Effects of the resistant starch on glucose, insulin, insulin resistance, and lipid parameters in overweight or obese adults: A systematic review and meta-analysis. Nutr. Diabetes 2019, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Venter, C.; Meyer, R.W.; Greenhawt, M.; Pali-Scholl, I.; Nwaru, B.; Roduit, C.; Untersmayr, E.; Adel-Patient, K.; Agache, I.; Agostoni, C.; et al. Role of dietary fiber in promoting immune health-An EAACI position paper. Allergy 2022, 77, 3185–3198. [Google Scholar] [CrossRef]
- Sahasrabudhe, N.M.; Beukema, M.; Tian, L.; Troost, B.; Scholte, J.; Bruininx, E.; Bruggeman, G.; van den Berg, M.; Scheurink, A.; Schols, H.A.; et al. Dietary Fiber Pectin Directly Blocks Toll-Like Receptor 2-1 and Prevents Doxorubicin-Induced Ileitis. Front. Immunol. 2018, 9, 383. [Google Scholar] [CrossRef]
- Hwang, Y.H.; Hong, S.G.; Mun, S.K.; Kim, S.J.; Lee, S.J.; Kim, J.J.; Kang, K.Y.; Yee, S.T. The Protective Effects of Astaxanthin on the OVA-Induced Asthma Mice Model. Molecules 2017, 22, 2019. [Google Scholar] [CrossRef]
- Fernandez-Prades, L.; Brasal-Prieto, M.; Alba, G.; Martin, V.; Montserrat-de la Paz, S.; Cejudo-Guillen, M.; Santa-Maria, C.; Dakhaoui, H.; Granados, B.; Sobrino, F.; et al. Sulforaphane Reduces the Chronic Inflammatory Immune Response of Human Dendritic Cells. Nutrients 2023, 15, 3405. [Google Scholar] [CrossRef]
- Brown, R.H.; Reynolds, C.; Brooker, A.; Talalay, P.; Fahey, J.W. Sulforaphane improves the bronchoprotective response in asthmatics through Nrf2-mediated gene pathways. Respir. Res. 2015, 16, 106. [Google Scholar] [CrossRef]
- Al-Harbi, N.O.; Nadeem, A.; Ahmad, S.F.; AlThagfan, S.S.; Alqinyah, M.; Alqahtani, F.; Ibrahim, K.E.; Al-Harbi, M.M. Sulforaphane treatment reverses corticosteroid resistance in a mixed granulocytic mouse model of asthma by upregulation of antioxidants and attenuation of Th17 immune responses in the airways. Eur. J. Pharmacol. 2019, 855, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Royce, S.G.; Licciardi, P.V.; Beh, R.C.; Bourke, J.E.; Donovan, C.; Hung, A.; Khurana, I.; Liang, J.J.; Maxwell, S.; Mazarakis, N.; et al. Sulforaphane prevents and reverses allergic airways disease in mice via anti-inflammatory, antioxidant, and epigenetic mechanisms. Cell. Mol. Life Sci. 2022, 79, 579. [Google Scholar] [CrossRef] [PubMed]
- Savoure, M.; Bousquet, J.; Jaakkola, J.J.K.; Jaakkola, M.S.; Jacquemin, B.; Nadif, R. Worldwide prevalence of rhinitis in adults: A review of definitions and temporal evolution. Clin. Transl. Allergy 2022, 12, e12130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lan, F.; Zhang, L. Update on pathomechanisms and treatments in allergic rhinitis. Allergy 2022, 77, 3309–3319. [Google Scholar] [CrossRef] [PubMed]
- Nur Husna, S.M.; Tan, H.T.; Md Shukri, N.; Mohd Ashari, N.S.; Wong, K.K. Allergic Rhinitis: A Clinical and Pathophysiological Overview. Front. Med. 2022, 9, 874114. [Google Scholar] [CrossRef] [PubMed]
- Kabashima, K.; Nakashima, C.; Nonomura, Y.; Otsuka, A.; Cardamone, C.; Parente, R.; De Feo, G.; Triggiani, M. Biomarkers for evaluation of mast cell and basophil activation. Immunol. Rev. 2018, 282, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Lam, H.Y.; Tergaonkar, V.; Ahn, K.S. Mechanisms of allergen-specific immunotherapy for allergic rhinitis and food allergies. Biosci. Rep. 2020, 40, BSR20200256. [Google Scholar] [CrossRef]
- Jafarinia, M.; Sadat Hosseini, M.; Kasiri, N.; Fazel, N.; Fathi, F.; Ganjalikhani Hakemi, M.; Eskandari, N. Quercetin with the potential effect on allergic diseases. Allergy Asthma Clin. Immunol. 2020, 16, 36. [Google Scholar] [CrossRef]
- Ashifha, S.; Vijayashree, J.; Vudayana, K.; Chintada, D.; Unnikrishnan, P. A Study of Cutaneous Adverse Drug Reactions at a Tertiary Care Center in Andhra Pradesh, India. Cureus 2023, 15, e37596. [Google Scholar] [CrossRef]
- Ebihara, N.; Takahashi, K.; Takemura, H.; Akanuma, Y.; Asano, K.; Sunagawa, M. Suppressive Effect of Quercetin on Nitric Oxide Production from Nasal Epithelial Cells In Vitro. Evid. Based Complement. Alternat. Med. 2018, 2018, 6097625. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, J.; Wang, C.; Fukunaga, A.; Li, S.; Yodoi, J.; Tian, H. Thioredoxin-1: A Promising Target for the Treatment of Allergic Diseases. Front. Immunol. 2022, 13, 883116. [Google Scholar] [CrossRef] [PubMed]
- Edo, Y.; Otaki, A.; Asano, K. Quercetin Enhances the Thioredoxin Production of Nasal Epithelial Cells In Vitro and In Vivo. Medicines 2018, 5, 124. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Shirai, M.; Inaba, Y.; Takara, T. Effects of repeated oral intake of a quercetin-containing supplement on allergic reaction: A randomized, placebo-controlled, double-blind parallel-group study. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 4331–4345. [Google Scholar] [CrossRef] [PubMed]
- Yusin, J.; Wang, V.; Henning, S.M.; Yang, J.; Tseng, C.H.; Thames, G.; Arnold, I.; Heber, D.; Lee, R.P.; Sanavio, L.; et al. The Effect of Broccoli Sprout Extract on Seasonal Grass Pollen-Induced Allergic Rhinitis. Nutrients 2021, 13, 1337. [Google Scholar] [CrossRef] [PubMed]
- Venter, C.; Meyer, R.W.; Nwaru, B.I.; Roduit, C.; Untersmayr, E.; Adel-Patient, K.; Agache, I.; Agostoni, C.; Akdis, C.A.; Bischoff, S.C.; et al. EAACI position paper: Influence of dietary fatty acids on asthma, food allergy, and atopic dermatitis. Allergy 2019, 74, 1429–1444. [Google Scholar] [CrossRef] [PubMed]
- Derakhshan, A.; Khodadoost, M.; Ghanei, M.; Gachkar, L.; Hajimahdipour, H.; Taghipour, A.; Yousefi, J.; Khoshkhui, M.; Azad, F.J. Effects of a Novel Barley-Based Formulation on Allergic Rhinitis: A Randomized Controlled Trial. Endocr. Metab. Immune Disord. Drug Targets 2019, 19, 1224–1231. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, R.; Liu, Y.; Gao, J.; Wu, Y.; Tu, C.; Chen, H.; Yuan, J. In Vitro Effect of Flavonoids on Basophils Degranulation and Intestinal Epithelial Barrier Damage Induced by omega-5 Gliadin-Derived Peptide. Foods 2022, 11, 3857. [Google Scholar] [CrossRef]
- Mayorga, C.; Palomares, F.; Canas, J.A.; Perez-Sanchez, N.; Nunez, R.; Torres, M.J.; Gomez, F. New Insights in Therapy for Food Allergy. Foods 2021, 10, 1037. [Google Scholar] [CrossRef]
- Li, J.; Zou, C.; Liu, Y. Amelioration of Ovalbumin-Induced Food Allergy in Mice by Targeted Rectal and Colonic Delivery of Cyanidin-3-O-Glucoside. Foods 2022, 11, 1542. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, P.; Zhang, J.; Hong, T. Bisdemethoxycurcumin attenuates OVA-induced food allergy by inhibiting the MAPK and NF-kappaB signaling pathways. Exp. Ther. Med. 2022, 23, 401. [Google Scholar] [CrossRef] [PubMed]
- Krajewski, D.; Polukort, S.H.; Gelzinis, J.; Rovatti, J.; Kaczenski, E.; Galinski, C.; Pantos, M.; Shah, N.N.; Schneider, S.S.; Kennedy, D.R.; et al. Protein Disulfide Isomerases Regulate IgE-Mediated Mast Cell Responses and Their Inhibition Confers Protective Effects During Food Allergy. Front. Immunol. 2020, 11, 606837. [Google Scholar] [CrossRef]
- Roth-Walter, F.; Afify, S.M.; Pacios, L.F.; Blokhuis, B.R.; Redegeld, F.; Regner, A.; Petje, L.M.; Fiocchi, A.; Untersmayr, E.; Dvorak, Z.; et al. Cow’s milk protein beta-lactoglobulin confers resilience against allergy by targeting complexed iron into immune cells. J. Allergy Clin. Immunol. 2021, 147, 321–334. [Google Scholar] [CrossRef]
- Wang, L.; Jia, X.; Yu, Q.; Shen, S.; Gao, Y.; Lin, X.; Zhang, W. Piper nigrum extract attenuates food allergy by decreasing Th2 cell response and regulating the Th17/Treg balance. Phytother Res. 2021, 35, 3214–3225. [Google Scholar] [CrossRef]
- Yang, N.; Maskey, A.R.; Srivastava, K.; Kim, M.; Wang, Z.; Musa, I.; Shi, Y.; Gong, Y.; Fidan, O.; Wang, J.; et al. Inhibition of pathologic immunoglobulin E in food allergy by EBF-2 and active compound berberine associated with immunometabolism regulation. Front. Immunol. 2023, 14, 1081121. [Google Scholar] [CrossRef]
- Srivastava, K.; Cao, M.; Fidan, O.; Shi, Y.; Yang, N.; Nowak-Wegrzyn, A.; Miao, M.; Zhan, J.; Sampson, H.A.; Li, X.M. Berberine-containing natural-medicine with boiled peanut-OIT induces sustained peanut-tolerance associated with distinct microbiota signature. Front. Immunol. 2023, 14, 1174907. [Google Scholar] [CrossRef] [PubMed]
- Hoppenbrouwers, T.; Cvejic Hogervorst, J.H.; Garssen, J.; Wichers, H.J.; Willemsen, L.E.M. Long Chain Polyunsaturated Fatty Acids (LCPUFAs) in the Prevention of Food Allergy. Front. Immunol. 2019, 10, 1118. [Google Scholar] [CrossRef] [PubMed]
- Hoppenbrouwers, T.; Fogliano, V.; Garssen, J.; Pellegrini, N.; Willemsen, L.E.M.; Wichers, H.J. Specific Polyunsaturated Fatty Acids Can Modulate in vitro Human moDC2s and Subsequent Th2 Cytokine Release. Front. Immunol. 2020, 11, 748. [Google Scholar] [CrossRef]
- Sartorio, M.U.A.; Pendezza, E.; Coppola, S.; Paparo, L.; D’Auria, E.; Zuccotti, G.V.; Berni Canani, R. Potential Role of Omega-3 Polyunsaturated Fatty Acids in Pediatric Food Allergy. Nutrients 2021, 14, 152. [Google Scholar] [CrossRef]
- Feketea, G.; Kostara, M.; Bumbacea, R.S.; Vassilopoulou, E.; Tsabouri, S. Vitamin D and Omega-3 (Fatty Acid) Supplementation in Pregnancy for the Primary Prevention of Food Allergy in Children-Literature Review. Children 2023, 10, 458. [Google Scholar] [CrossRef]
- Jerzynska, A.; Polanska, A.; Trafalska, E.; Jankowska, A.; Podlecka, D.; Brzozowska, A. Prenatal polyunsaturated fatty acids and atopic dermatitis and food allergy in children from Polish Mother and Child Cohort study. Int. J. Occup. Med. Environ. Health 2023, 36, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Huynh, L.B.P.; Nguyen, N.N.; Fan, H.Y.; Huang, S.Y.; Huang, C.H.; Chen, Y.C. Maternal Omega-3 Supplementation During Pregnancy, but Not Childhood Supplementation, Reduces the Risk of Food Allergy Diseases in Offspring. J. Allergy Clin. Immunol. Pract. 2023, 11, 2862–2871. [Google Scholar] [CrossRef] [PubMed]
- Pecoraro, L.; Dalle Carbonare, L.; Castagnoli, R.; Marseglia, G.L.; Piacentini, G.; Pietrobelli, A. IgE-mediated fish allergy in children: Is omega-3 supplementation useful? Int. J. Food Sci. Nutr. 2022, 73, 154–157. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Vuillermin, P.J.; Goverse, G.; Vinuesa, C.G.; Mebius, R.E.; Macia, L.; Mackay, C.R. Dietary Fiber and Bacterial SCFA Enhance Oral Tolerance and Protect against Food Allergy through Diverse Cellular Pathways. Cell Rep. 2016, 15, 2809–2824. [Google Scholar] [CrossRef] [PubMed]
- Folkerts, J.; Redegeld, F.; Folkerts, G.; Blokhuis, B.; van den Berg, M.P.M.; de Bruijn, M.J.W.; van Ijcken, W.F.J.; Junt, T.; Tam, S.Y.; Galli, S.J.; et al. Butyrate inhibits human mast cell activation via epigenetic regulation of FcepsilonRI-mediated signaling. Allergy 2020, 75, 1966–1978. [Google Scholar] [CrossRef] [PubMed]
- Roduit, C.; Frei, R.; Ferstl, R.; Loeliger, S.; Westermann, P.; Rhyner, C.; Schiavi, E.; Barcik, W.; Rodriguez-Perez, N.; Wawrzyniak, M.; et al. High levels of butyrate and propionate in early life are associated with protection against atopy. Allergy 2019, 74, 799–809. [Google Scholar] [CrossRef]
- Berni Canani, R.; De Filippis, F.; Nocerino, R.; Paparo, L.; Di Scala, C.; Cosenza, L.; Della Gatta, G.; Calignano, A.; De Caro, C.; Laiola, M.; et al. Gut microbiota composition and butyrate production in children affected by non-IgE-mediated cow’s milk allergy. Sci. Rep. 2018, 8, 12500. [Google Scholar] [CrossRef]
- Tian, L.; Wang, M.; Wang, Y.; Li, W.; Yang, Y. Naringenin ameliorates atopic dermatitis by inhibiting inflammation and enhancing immunity through the JAK2/STAT3 pathway. Genes Genom. 2023. online ahead of print. [Google Scholar] [CrossRef]
- Sun, R.; Liu, C.; Liu, J.; Yin, S.; Song, R.; Ma, J.; Cao, G.; Lu, Y.; Zhang, G.; Wu, Z.; et al. Integrated network pharmacology and experimental validation to explore the mechanisms underlying naringenin treatment of chronic wounds. Sci. Rep. 2023, 13, 132. [Google Scholar] [CrossRef]
- Lee, C.H.; Yang, H.; Park, J.H.Y.; Kim, J.E.; Lee, K.W. Piceatannol, a metabolite of resveratrol, attenuates atopic dermatitis by targeting Janus kinase 1. Phytomedicine 2022, 99, 153981. [Google Scholar] [CrossRef]
- Shen, Y.; Xu, J. Resveratrol Exerts Therapeutic Effects on Mice with Atopic Dermatitis. Wounds 2019, 31, 279–284. [Google Scholar] [PubMed]
- Bangash, Y.; Saleem, A.; Akhtar, M.F.; Anwar, F.; Akhtar, B.; Sharif, A.; Khan, M.I.; Khan, A. Pterostilbene reduces the progression of atopic dermatitis via modulating inflammatory and oxidative stress biomarkers in mice. Inflammopharmacology 2023, 31, 1289–1303. [Google Scholar] [CrossRef] [PubMed]
- Cassano, R.; Serini, S.; Curcio, F.; Trombino, S.; Calviello, G. Preparation and Study of Solid Lipid Nanoparticles Based on Curcumin, Resveratrol and Capsaicin Containing Linolenic Acid. Pharmaceutics 2022, 14, 1593. [Google Scholar] [CrossRef] [PubMed]
- Conte, R.; De Luca, I.; Valentino, A.; Cerruti, P.; Pedram, P.; Cabrera-Barjas, G.; Moeini, A.; Calarco, A. Hyaluronic Acid Hydrogel Containing Resveratrol-Loaded Chitosan Nanoparticles as an Adjuvant in Atopic Dermatitis Treatment. J. Funct. Biomater. 2023, 14, 82. [Google Scholar] [CrossRef]
- Nakagawa, S.; Hillebrand, G.G.; Nunez, G. Rosmarinus officinalis L. (Rosemary) Extracts Containing Carnosic Acid and Carnosol are Potent Quorum Sensing Inhibitors of Staphylococcus aureus Virulence. Antibiotics 2020, 9, 149. [Google Scholar] [CrossRef]
- Lee, D.Y.; Hwang, C.J.; Choi, J.Y.; Park, M.H.; Song, M.J.; Oh, K.W.; Son, D.J.; Lee, S.H.; Han, S.B.; Hong, J.T. Inhibitory Effect of Carnosol on Phthalic Anhydride-Induced Atopic Dermatitis via Inhibition of STAT3. Biomol. Ther. 2017, 25, 535–544. [Google Scholar] [CrossRef]
- Yeo, I.J.; Park, J.H.; Jang, J.S.; Lee, D.Y.; Park, J.E.; Choi, Y.E.; Joo, J.H.; Song, J.K.; Jeon, H.O.; Hong, J.T. Inhibitory effect of Carnosol on UVB-induced inflammation via inhibition of STAT3. Arch. Pharm. Res. 2019, 42, 274–283. [Google Scholar] [CrossRef]
- Vollono, L.; Falconi, M.; Gaziano, R.; Iacovelli, F.; Dika, E.; Terracciano, C.; Bianchi, L.; Campione, E. Potential of Curcumin in Skin Disorders. Nutrients 2019, 11, 2169. [Google Scholar] [CrossRef]
- Sharma, S.; Sethi, G.S.; Naura, A.S. Curcumin Ameliorates Ovalbumin-Induced Atopic Dermatitis and Blocks the Progression of Atopic March in Mice. Inflammation 2020, 43, 358–369. [Google Scholar] [CrossRef]
- Chen, Y.L.; Chang, C.C.; Lin, Y.C.; Chen, M.C. Double-layered PLGA/HA microneedle systems as a long-acting formulation of polyphenols for effective and long-term management of atopic dermatitis. Biomater. Sci. 2023, 11, 4995–5011. [Google Scholar] [CrossRef]
- Rakha, A.; Umar, N.; Rabail, R.; Butt, M.S.; Kieliszek, M.; Hassoun, A.; Aadil, R.M. Anti-inflammatory and anti-allergic potential of dietary flavonoids: A review. Biomed. Pharmacother. 2022, 156, 113945. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.N.; Shin, S.A.; Choo, G.S.; Kim, H.J.; Park, Y.S.; Kim, B.S.; Kim, S.K.; Cho, S.D.; Nam, J.S.; Choi, C.S.; et al. Anti-inflammatory effect of quercetin and galangin in LPS-stimulated RAW264.7 macrophages and DNCB-induced atopic dermatitis animal models. Int. J. Mol. Med. 2018, 41, 888–898. [Google Scholar] [CrossRef] [PubMed]
- Beken, B.; Serttas, R.; Yazicioglu, M.; Turkekul, K.; Erdogan, S. Quercetin Improves Inflammation, Oxidative Stress, and Impaired Wound Healing in Atopic Dermatitis Model of Human Keratinocytes. Pediatr. Allergy Immunol. Pulmonol. 2020, 33, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xian, Y.F.; Loo, S.K.F.; Ip, S.P.; Yang, W.; Chan, W.Y.; Lin, Z.X.; Wu, J.C.Y. Baicalin ameliorates 2,4-dinitrochlorobenzene-induced atopic dermatitis-like skin lesions in mice through modulating skin barrier function, gut microbiota and JAK/STAT pathway. Bioorg. Chem. 2022, 119, 105538. [Google Scholar] [CrossRef] [PubMed]
- Andoh, T.; Yoshihisa, Y.; Rehman, M.U.; Tabuchi, Y.; Shimizu, T. Berberine induces anti-atopic dermatitis effects through the downregulation of cutaneous EIF3F and MALT1 in NC/Nga mice with atopy-like dermatitis. Biochem. Pharmacol. 2021, 185, 114439. [Google Scholar] [CrossRef]
- Balic, A.; Vlasic, D.; Zuzul, K.; Marinovic, B.; Bukvic Mokos, Z. Omega-3 Versus Omega-6 Polyunsaturated Fatty Acids in the Prevention and Treatment of Inflammatory Skin Diseases. Int. J. Mol. Sci. 2020, 21, 741. [Google Scholar] [CrossRef]
- Huang, T.H.; Wang, P.W.; Yang, S.C.; Chou, W.L.; Fang, J.Y. Cosmetic and Therapeutic Applications of Fish Oil’s Fatty Acids on the Skin. Mar. Drugs 2018, 16, 256. [Google Scholar] [CrossRef]
- Kang, J.; Im, D.S. FFA2 Activation Ameliorates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis in Mice. Biomol. Ther. 2020, 28, 267–271. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, K.; Kim, W. Cream Cheese-Derived Lactococcus chungangensis CAU 28 Modulates the Gut Microbiota and Alleviates Atopic Dermatitis in BALB/c Mice. Sci. Rep. 2019, 9, 446. [Google Scholar] [CrossRef]
- Lee, Y.H.; Kalailingam, P.; Delcour, J.A.; Fogliano, V.; Thanabalu, T. Olive-Derived Antioxidant Dietary Fiber Modulates Gut Microbiota Composition and Attenuates Atopic Dermatitis Like Inflammation in Mice. Mol. Nutr. Food Res. 2023, 67, e2200127. [Google Scholar] [CrossRef]
- Lee, M.J.; Park, Y.M.; Kim, B.; Tae, I.H.; Kim, N.E.; Pranata, M.; Kim, T.; Won, S.; Kang, N.J.; Lee, Y.K.; et al. Disordered development of gut microbiome interferes with the establishment of the gut ecosystem during early childhood with atopic dermatitis. Gut Microbes 2022, 14, 2068366. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.C.; Lo, Y.H.; Huang, S.Y.; Liu, H.L.; Yao, Z.K.; Chang, C.I.; Wen, Z.H. The anti-inflammatory properties of ethyl acetate fraction in ethanol extract from Sarcodia suiae sp. alleviates atopic dermatitis-like lesion in mice. Biosci. Biotechnol. Biochem. 2022, 86, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Matano, Y.; Morita, T.; Ito, M.; Okazaki, S.; Koto, M.; Ichikawa, Y.; Takayama, R.; Hoashi, T.; Saeki, H.; Kanda, N. Dietary habits in Japanese patients with chronic spontaneous urticaria. Australas. J. Dermatol. 2020, 61, e333–e338. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Jeon, S.H.; Ham, H.J.; Lee, H.P.; Song, M.J.; Hong, J.T. Improved Anti-Inflammatory Effects of Liposomal Astaxanthin on a Phthalic Anhydride-Induced Atopic Dermatitis Model. Front. Immunol. 2020, 11, 565285. [Google Scholar] [CrossRef]
- Park, J.H.; Yeo, I.J.; Jang, J.S.; Kim, K.C.; Park, M.H.; Lee, H.P.; Han, S.B.; Hong, J.T. Combination Effect of Titrated Extract of Centella asiatica and Astaxanthin in a Mouse Model of Phthalic Anhydride-Induced Atopic Dermatitis. Allergy Asthma Immunol. Res. 2019, 11, 548–559. [Google Scholar] [CrossRef]
- Alyoussef, A. Attenuation of experimentally induced atopic dermatitis in mice by sulforaphane: Effect on inflammation and apoptosis. Toxicol. Mech. Methods 2022, 32, 224–232. [Google Scholar] [CrossRef]
- Wu, W.; Peng, G.; Yang, F.; Zhang, Y.; Mu, Z.; Han, X. Sulforaphane has a therapeutic effect in an atopic dermatitis murine model and activates the Nrf2/HO-1 axis. Mol. Med. Rep. 2019, 20, 1761–1771. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Enríquez, S.; Múnera-Rodríguez, A.M.; Leiva-Castro, C.; Sobrino, F.; Palomares, F. Modulation of the Immune Response to Allergies Using Alternative Functional Foods. Int. J. Mol. Sci. 2024, 25, 467. https://doi.org/10.3390/ijms25010467
López-Enríquez S, Múnera-Rodríguez AM, Leiva-Castro C, Sobrino F, Palomares F. Modulation of the Immune Response to Allergies Using Alternative Functional Foods. International Journal of Molecular Sciences. 2024; 25(1):467. https://doi.org/10.3390/ijms25010467
Chicago/Turabian StyleLópez-Enríquez, Soledad, Ana M. Múnera-Rodríguez, Camila Leiva-Castro, Francisco Sobrino, and Francisca Palomares. 2024. "Modulation of the Immune Response to Allergies Using Alternative Functional Foods" International Journal of Molecular Sciences 25, no. 1: 467. https://doi.org/10.3390/ijms25010467
APA StyleLópez-Enríquez, S., Múnera-Rodríguez, A. M., Leiva-Castro, C., Sobrino, F., & Palomares, F. (2024). Modulation of the Immune Response to Allergies Using Alternative Functional Foods. International Journal of Molecular Sciences, 25(1), 467. https://doi.org/10.3390/ijms25010467