
Cannabinoids, Endocannabinoids, and Synthetic Cannabimimetic Molecules in Neuromuscular Disorders


{kind=link}
Abstract
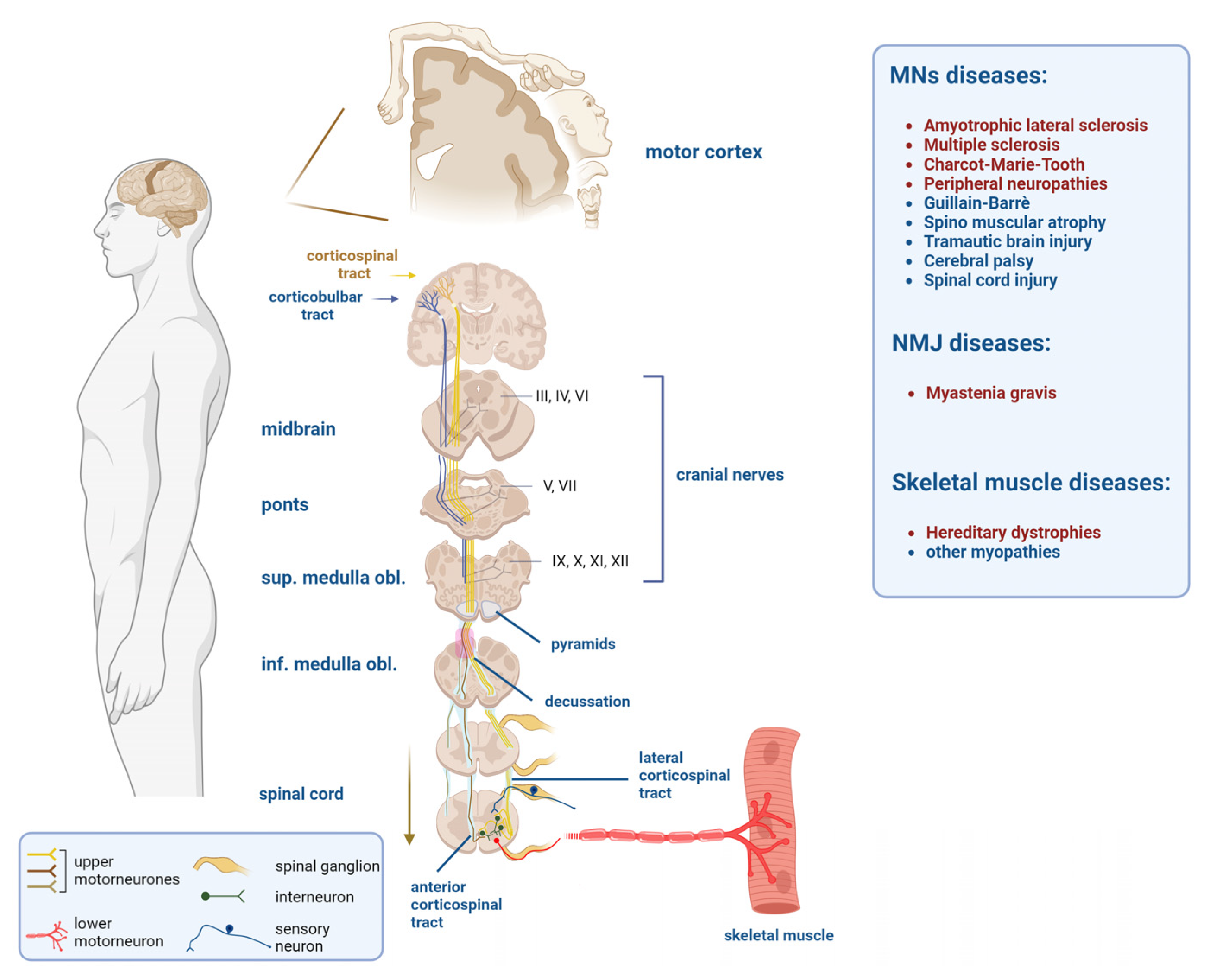
1. Neuromuscular Diseases
2. Cannabinoids, Endocannabinoids, and Synthetic Cannabimimetic Molecules
3. Amyotrophic Lateral Sclerosis
4. Multiple Sclerosis
5. Charcot–Marie–Tooth (CMT)
6. Peripheral Neuropathy
7. Myasthenia Gravis
8. Muscular Dystrophies
9. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, L.A.; Macken, W.L.; Perry, L.D.; Record, C.J.; Schon, K.R.; Frezatti, R.S.S.; Raga, S.; Naidu, K.; Köken, Ö.Y.; Polat, I.; et al. Neuromuscular Disease Genetics in Underrepresented Populations: Increasing Data Diversity. Brain 2023, 146, awad254. [Google Scholar] [CrossRef]
- Porter, R. The Corticomotoneuronal Component of the Pyramidal Tract: Corticomotoneuronal Connections and Functions in Primates. Brain Res. 1985, 357, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, N.; Yadala, S. Neuroanatomy, Corticobulbar Tract. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Morrison, B.M. Neuromuscular Diseases. Semin. Neurol. 2016, 36, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Mary, P.; Servais, L.; Vialle, R. Neuromuscular Diseases: Diagnosis and Management. Orthop. Traumatol. Surg. Res. 2018, 104, S89–S95. [Google Scholar] [CrossRef] [PubMed]
- McDonald, C.M. Clinical approach to the diagnostic evaluation of herditary and acquired neuromuscular diseases. Phys. Med. Rehabil. Clin. 2012, 23, 495–563. [Google Scholar] [CrossRef] [PubMed]
- Kokontis, L.; Gutmann, L. Current Treatment of Neuromuscular Diseases. Arch. Neurol. 2000, 57, 939–943. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tahir, M.N.; Shahbazi, F.; Rondeau-Gagné, S.; Trant, J.F. The Biosynthesis of the Cannabinoids. J. Cannabis Res. 2021, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Cascio, M.G.; Pertwee, R.G. Known Pharmacological Actions of Nine Nonpsychotropic Phytocannabinoids. In Handbook of Cannabis; Pertwee, R., Ed.; Oxford University Press: Oxford, UK, 2014; ISBN 978-0-19-966268-5. [Google Scholar]
- Di Marzo, V.; Piscitelli, F. The Endocannabinoid System and Its Modulation by Phytocannabinoids. Neurotherapeutics 2015, 12, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Vitale, R.M.; Iannotti, F.A.; Amodeo, P. The (Poly)Pharmacology of Cannabidiol in Neurological and Neuropsychiatric Disorders: Molecular Mechanisms and Targets. Int. J. Mol. Sci. 2021, 22, 4876. [Google Scholar] [CrossRef]
- Morales, P.; Reggio, P.H. An Update on Non-CB1, Non-CB2 Cannabinoid Related G-Protein-Coupled Receptors. Cannabis Cannabinoid Res. 2017, 2, 265–273. [Google Scholar] [CrossRef]
- O’Sullivan, S.E. An Update on PPAR Activation by Cannabinoids. Br. J. Pharmacol. 2016, 173, 1899–1910. [Google Scholar] [CrossRef] [PubMed]
- de Brito Siqueira, A.L.G.; Cremasco, P.V.V.; Bahú, J.O.; Pioli da Silva, A.; Melo de Andrade, L.R.; González, P.G.A.; Crivellin, S.; Cárdenas Concha, V.O.; Krambeck, K.; Lodi, L.; et al. Phytocannabinoids: Pharmacological Effects, Biomedical Applications, and Worldwide Prospection. J. Tradit. Complement. Med. 2023, 13, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Ligresti, A.; De Petrocellis, L.; Di Marzo, V. From Phytocannabinoids to Cannabinoid Receptors and Endocannabinoids: Pleiotropic Physiological and Pathological Roles Through Complex Pharmacology. Physiol. Rev. 2016, 96, 1593–1659. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Ruiz, J.; Galve-Roperh, I.; Sagredo, O.; Guzmán, M. Possible Therapeutic Applications of Cannabis in the Neuropsychopharmacology Field. Eur. Neuropsychopharmacol. 2020, 36, 217–234. [Google Scholar] [CrossRef] [PubMed]
- Stone, N.L.; Murphy, A.J.; England, T.J.; O’Sullivan, S.E. A Systematic Review of Minor Phytocannabinoids with Promising Neuroprotective Potential. Br. J. Pharmacol. 2020, 177, 4330–4352. [Google Scholar] [CrossRef] [PubMed]
- Iannotti, F.A.; Di Marzo, V.; Petrosino, S. Endocannabinoids and Endocannabinoid-Related Mediators: Targets, Metabolism and Role in Neurological Disorders. Prog. Lipid Res. 2016, 62, 107–128. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; Stella, N.; Zimmer, A. Endocannabinoid Signalling and the Deteriorating Brain. Nat. Rev. Neurosci. 2015, 16, 30–42. [Google Scholar] [CrossRef]
- Galve-Roperh, I.; Aguado, T.; Palazuelos, J.; Guzmán, M. The Endocannabinoid System and Neurogenesis in Health and Disease. Neuroscientist 2007, 13, 109–114. [Google Scholar] [CrossRef]
- Mackie, K. Mechanisms of CB1 Receptor Signaling: Endocannabinoid Modulation of Synaptic Strength. Int. J. Obes. 2006, 30 (Suppl. S1), S19–S23. [Google Scholar] [CrossRef]
- Kettunen, P.; Kyriakatos, A.; Hallén, K.; El Manira, A. Neuromodulation via Conditional Release of Endocannabinoids in the Spinal Locomotor Network. Neuron 2005, 45, 95–104. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J. The Endocannabinoid System as a Target for the Treatment of Motor Dysfunction. Br. J. Pharmacol. 2009, 156, 1029–1040. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, R.; Nielsen, L.M.; Holm, N.B.; Rasmussen, H.B.; Linnet, K.; Indices Consortium. Synthetic Cannabimimetic Agents Metabolized by Carboxylesterases. Drug Test. Anal. 2015, 7, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Petrosino, S.; Di Marzo, V. FAAH and MAGL Inhibitors: Therapeutic Opportunities from Regulating Endocannabinoid Levels. Curr. Opin. Investig. Drugs 2010, 11, 51–62. [Google Scholar] [PubMed]
- Talbott, E.O.; Malek, A.M.; Lacomis, D. The Epidemiology of Amyotrophic Lateral Sclerosis. Handb. Clin. Neurol. 2016, 138, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.; Smoum, R.; Mechoulam, R.; Steinberg, D. Potential Combinations of Endocannabinoid/Endocannabinoid-like Compounds and Antibiotics against Methicillin-Resistant Staphylococcus Aureus. PLoS ONE 2020, 15, e0231583. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Guan, L.; Deng, M. Recent Progress of the Genetics of Amyotrophic Lateral Sclerosis and Challenges of Gene Therapy. Front. Neurosci. 2023, 17, 1170996. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J. Disease-Modifying Treatment of Amyotrophic Lateral Sclerosis. Am. J. Manag. Care 2018, 24, S327–S335. [Google Scholar] [PubMed]
- Witzel, S.; Maier, A.; Steinbach, R.; Grosskreutz, J.; Koch, J.C.; Sarikidi, A.; Petri, S.; Günther, R.; Wolf, J.; Hermann, A.; et al. Safety and Effectiveness of Long-Term Intravenous Administration of Edaravone for Treatment of Patients With Amyotrophic Lateral Sclerosis. JAMA Neurol. 2022, 79, 121–130. [Google Scholar] [CrossRef]
- Raman, C.; McAllister, S.D.; Rizvi, G.; Patel, S.G.; Moore, D.H.; Abood, M.E. Amyotrophic Lateral Sclerosis: Delayed Disease Progression in Mice by Treatment with a Cannabinoid. Amyotroph. Lateral Scler. Other Mot. Neuron Disord. 2004, 5, 33–39. [Google Scholar] [CrossRef]
- Weydt, P.; Hong, S.; Witting, A.; Möller, T.; Stella, N.; Kliot, M. Cannabinol Delays Symptom Onset in SOD1 (G93A) Transgenic Mice without Affecting Survival. Amyotroph. Lateral Scler. 2005, 6, 182–184. [Google Scholar] [CrossRef]
- Bilsland, L.G.; Dick, J.R.T.; Pryce, G.; Petrosino, S.; Di Marzo, V.; Baker, D.; Greensmith, L. Increasing Cannabinoid Levels by Pharmacological and Genetic Manipulation Delay Disease Progression in SOD1 Mice. FASEB J. 2006, 20, 1003–1005. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Moore, D.H.; Makriyannis, A.; Abood, M.E. AM1241, a Cannabinoid CB2 Receptor Selective Compound, Delays Disease Progression in a Mouse Model of Amyotrophic Lateral Sclerosis. Eur. J. Pharmacol. 2006, 542, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, J.L.; Seely, K.A.; Reed, R.L.; Crow, J.P.; Prather, P.L. The CB2 Cannabinoid Agonist AM-1241 Prolongs Survival in a Transgenic Mouse Model of Amyotrophic Lateral Sclerosis When Initiated at Symptom Onset. J. Neurochem. 2007, 101, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cueto, C.; Santos-García, I.; García-Toscano, L.; Espejo-Porras, F.; Bellido, M.; Fernández-Ruiz, J.; Muñoz, E.; de Lago, E. Neuroprotective Effects of the Cannabigerol Quinone Derivative VCE-003.2 in SOD1G93A Transgenic Mice, an Experimental Model of Amyotrophic Lateral Sclerosis. Biochem. Pharmacol. 2018, 157, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Martet, M.; Espejo-Porras, F.; Fernández-Ruiz, J.; de Lago, E. Changes in Endocannabinoid Receptors and Enzymes in the Spinal Cord of SOD1(G93A) Transgenic Mice and Evaluation of a Sativex(®) -like Combination of Phytocannabinoids: Interest for Future Therapies in Amyotrophic Lateral Sclerosis. CNS Neurosci. Ther. 2014, 20, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Espejo-Porras, F.; Piscitelli, F.; Verde, R.; Ramos, J.A.; Di Marzo, V.; de Lago, E.; Fernández-Ruiz, J. Changes in the Endocannabinoid Signaling System in CNS Structures of TDP-43 Transgenic Mice: Relevance for a Neuroprotective Therapy in TDP-43-Related Disorders. J. Neuroimmune Pharmacol. 2015, 10, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cueto, C.; Gómez-Almería, M.; García Toscano, L.; Romero, J.; Hillard, C.J.; de Lago, E.; Fernández-Ruiz, J. Inactivation of the CB2 Receptor Accelerated the Neuropathological Deterioration in TDP-43 Transgenic Mice, a Model of Amyotrophic Lateral Sclerosis. Brain Pathol. 2021, 31, e12972. [Google Scholar] [CrossRef] [PubMed]
- Riva, N.; Mora, G.; Sorarù, G.; Lunetta, C.; Ferraro, O.E.; Falzone, Y.; Leocani, L.; Fazio, R.; Comola, M.; Comi, G.; et al. Safety and Efficacy of Nabiximols on Spasticity Symptoms in Patients with Motor Neuron Disease (CANALS): A Multicentre, Double-Blind, Randomised, Placebo-Controlled, Phase 2 Trial. Lancet Neurol. 2019, 18, 155–164. [Google Scholar] [CrossRef]
- Meyer, T.; Funke, A.; Münch, C.; Kettemann, D.; Maier, A.; Walter, B.; Thomas, A.; Spittel, S. Real World Experience of Patients with Amyotrophic Lateral Sclerosis (ALS) in the Treatment of Spasticity Using Tetrahydrocannabinol:Cannabidiol (THC:CBD). BMC Neurol. 2019, 19, 222. [Google Scholar] [CrossRef]
- Lacroix, C.; Alleman-Brimault, I.; Zalta, A.; Rouby, F.; Cassé-Perrot, C.; Jouve, E.; Attolini, L.; Guilhaumou, R.; Micallef, J.; Blin, O. What Do We Know About Medical Cannabis in Neurological Disorders and What Are the Next Steps? Front. Pharmacol. 2022, 13, 883987. [Google Scholar] [CrossRef]
- Prashad, S.; Filbey, F.M. Cognitive Motor Deficits in Cannabis Users. Curr. Opin. Behav. Sci. 2017, 13, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tafti, D.; Ehsan, M.; Xixis, K.L. Multiple Sclerosis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Ziemssen, T.; Bhan, V.; Chataway, J.; Chitnis, T.; Campbell Cree, B.A.; Havrdova, E.K.; Kappos, L.; Labauge, P.; Miller, A.; Nakahara, J.; et al. Secondary Progressive Multiple Sclerosis. Neurol. Neuroimmunol. Neuroinflamm 2022, 10, e200064. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.; Kingwell, E.; Rieckmann, P.; Tremlett, H.; Neurologists, U.M.C. The Natural History of Secondary Progressive Multiple Sclerosis. J. Neurol. Neurosurg. Psychiatry 2010, 81, 1039–1043. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, T. Time to Reconsider the Classification of Multiple Sclerosis. Lancet Neurol. 2023, 22, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Samkoff, L.M.; Goodman, A.D. Symptomatic Management in Multiple Sclerosis. Neurol. Clin. 2011, 29, 449–463. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G. Cannabinoids and Multiple Sclerosis. Pharmacol. Ther. 2002, 95, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Zettl, U.K.; Rommer, P.; Hipp, P.; Patejdl, R. Evidence for the Efficacy and Effectiveness of THC-CBD Oromucosal Spray in Symptom Management of Patients with Spasticity Due to Multiple Sclerosis. Ther. Adv. Neurol. Disord. 2016, 9, 9–30. [Google Scholar] [CrossRef]
- Haddad, M. The Impact of CB1 Receptor on Inflammation in Skeletal Muscle Cells. JIR 2021, 14, 3959–3967. [Google Scholar] [CrossRef]
- Flachenecker, P.; Henze, T.; Zettl, U.K. Nabiximols (THC/CBD Oromucosal Spray, Sativex®) in Clinical Practice--Results of a Multicenter, Non-Interventional Study (MOVE 2) in Patients with Multiple Sclerosis Spasticity. Eur. Neurol. 2014, 71, 271–279. [Google Scholar] [CrossRef]
- Patti, F.; Messina, S.; Solaro, C.; Amato, M.P.; Bergamaschi, R.; Bonavita, S.; Bruno Bossio, R.; Brescia Morra, V.; Costantino, G.F.; Cavalla, P.; et al. Efficacy and Safety of Cannabinoid Oromucosal Spray for Multiple Sclerosis Spasticity. J. Neurol. Neurosurg. Psychiatry 2016, 87, 944–951. [Google Scholar] [CrossRef]
- da Rovare, V.P.; Magalhães, G.P.A.; Jardini, G.D.A.; Beraldo, M.L.; Gameiro, M.O.; Agarwal, A.; Luvizutto, G.J.; Paula-Ramos, L.; Camargo, S.E.A.; de Oliveira, L.D.; et al. Cannabinoids for Spasticity Due to Multiple Sclerosis or Paraplegia: A Systematic Review and Meta-Analysis of Randomized Clinical Trials. Complement. Ther. Med. 2017, 34, 170–185. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.S.; Gustavsen, S.; Roshanisefat, H.; Kant, M.; Biering-Sørensen, F.; Andersen, C.; Olsson, A.; Chow, H.H.; Asgari, N.; Hansen, J.R.; et al. Cannabis-Based Medicine for Neuropathic Pain and Spasticity—A Multicenter, Randomized, Double-Blinded, Placebo-Controlled Trial. Pharmaceuticals 2023, 16, 1079. [Google Scholar] [CrossRef] [PubMed]
- Corey-Bloom, J.; Wolfson, T.; Gamst, A.; Jin, S.; Marcotte, T.D.; Bentley, H.; Gouaux, B. Smoked Cannabis for Spasticity in Multiple Sclerosis: A Randomized, Placebo-Controlled Trial. Cmaj 2012, 184, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Jones, É.; Vlachou, S. A Critical Review of the Role of the Cannabinoid Compounds Δ9-Tetrahydrocannabinol (Δ9-THC) and Cannabidiol (CBD) and Their Combination in Multiple Sclerosis Treatment. Molecules 2020, 25, 4930. [Google Scholar] [CrossRef] [PubMed]
- Pisciotta, C.; Pareyson, D. Gene Therapy and Other Novel Treatment Approaches for Charcot-Marie-Tooth Disease. Neuromuscul. Disord. 2023, 33, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Pisciotta, C.; Saveri, P.; Pareyson, D. Updated Review of Therapeutic Strategies for Charcot-Marie-Tooth Disease and Related Neuropathies. Expert. Rev. Neurother. 2021, 21, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Pisciotta, C.; Saveri, P.; Pareyson, D. Challenges in Treating Charcot-Marie-Tooth Disease and Related Neuropathies: Current Management and Future Perspectives. Brain Sci. 2021, 11, 1447. [Google Scholar] [CrossRef]
- Canals, P.C.; Aguilar, A.G.; Carter, G.T.; Shields, C.M.; Westerkamp, A.; D’Elia, M.; Aldrich, J.; Moore, R.N.; Moore, A.T.; Piper, B.J. Patient Reported Outcomes Using Medical Cannabis for Managing Pain in Charcot-Marie-Tooth Disease. Am. J. Hosp. Palliat. Care 2023, 40, 1163–1167. [Google Scholar] [CrossRef]
- Quintão, N.L.M.; Santin, J.R.; Stoeberl, L.C.; Corrêa, T.P.; Melato, J.; Costa, R. Pharmacological Treatment of Chemotherapy-Induced Neuropathic Pain: PPARγ Agonists as a Promising Tool. Front. Neurosci. 2019, 13, 907. [Google Scholar] [CrossRef]
- Clark, M.R.; Cox, T.S. Refractory Chronic Pain. Psychiatr. Clin. N. Am. 2002, 25, 71–88. [Google Scholar] [CrossRef]
- Taylor, R.S. Epidemiology of Refractory Neuropathic Pain. Pain. Pract. 2006, 6, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Maione, S.; Costa, B.; Di Marzo, V. Endocannabinoids: A Unique Opportunity to Develop Multitarget Analgesics. Pain 2013, 154, S87–S93. [Google Scholar] [CrossRef] [PubMed]
- Guindon, J.; Hohmann, A.G. The Endocannabinoid System and Pain. CNS Neurol. Disord. Drug Targets 2009, 8, 403–421. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, E.; Luongo, L.; de Novellis, V.; Rossi, F.; Maione, S. The Role of Cannabinoid Receptors in the Descending Modulation of Pain. Pharmaceuticals 2010, 3, 2661–2673. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, E.; Rossi, F.; Maione, S. Role of TRPV1 Receptors in Descending Modulation of Pain. Mol. Cell Endocrinol. 2008, 286, S79–S83. [Google Scholar] [CrossRef] [PubMed]
- Suplita, R.L.; Farthing, J.N.; Gutierrez, T.; Hohmann, A.G. Inhibition of Fatty-Acid Amide Hydrolase Enhances Cannabinoid Stress-Induced Analgesia: Sites of Action in the Dorsolateral Periaqueductal Gray and Rostral Ventromedial Medulla. Neuropharmacology 2005, 49, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Vučković, S.; Srebro, D.; Vujović, K.S.; Vučetić, Č.; Prostran, M. Cannabinoids and Pain: New Insights From Old Molecules. Front. Pharmacol. 2018, 9, 1259. [Google Scholar] [CrossRef]
- Luongo, L.; Maione, S.; Di Marzo, V. Endocannabinoids and Neuropathic Pain: Focus on Neuron-Glia and Endocannabinoid-Neurotrophin Interactions. Eur. J. Neurosci. 2014, 39, 401–408. [Google Scholar] [CrossRef]
- Starowicz, K.; Finn, D.P. Cannabinoids and Pain: Sites and Mechanisms of Action. Adv. Pharmacol. 2017, 80, 437–475. [Google Scholar] [CrossRef]
- Lee, G.; Grovey, B.; Furnish, T.; Wallace, M. Medical Cannabis for Neuropathic Pain. Curr. Pain. Headache Rep. 2018, 22, 8. [Google Scholar] [CrossRef]
- Berman, J.S.; Symonds, C.; Birch, R. Efficacy of Two Cannabis Based Medicinal Extracts for Relief of Central Neuropathic Pain from Brachial Plexus Avulsion: Results of a Randomised Controlled Trial. Pain 2004, 112, 299–306. [Google Scholar] [CrossRef]
- Luongo, L.; Starowicz, K.; Maione, S.; Di Marzo, V. Allodynia Lowering Induced by Cannabinoids and Endocannabinoids (ALICE). Pharmacol. Res. 2017, 119, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Nagarkatti, P.; Pandey, R.; Rieder, S.A.; Hegde, V.L.; Nagarkatti, M. Cannabinoids as Novel Anti-Inflammatory Drugs. Future Med. Chem. 2009, 1, 1333–1349. [Google Scholar] [CrossRef] [PubMed]
- Wilsey, B.; Marcotte, T.; Tsodikov, A.; Millman, J.; Bentley, H.; Gouaux, B.; Fishman, S. A Randomized, Placebo-Controlled, Crossover Trial of Cannabis Cigarettes in Neuropathic Pain. J. Pain. 2008, 9, 506–521. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.J.; Toperoff, W.; Vaida, F.; van den Brande, G.; Gonzales, J.; Gouaux, B.; Bentley, H.; Atkinson, J.H. Smoked Medicinal Cannabis for Neuropathic Pain in HIV: A Randomized, Crossover Clinical Trial. Neuropsychopharmacology 2009, 34, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Ware, M.A.; Wang, T.; Shapiro, S.; Robinson, A.; Ducruet, T.; Huynh, T.; Gamsa, A.; Bennett, G.J.; Collet, J.-P. Smoked Cannabis for Chronic Neuropathic Pain: A Randomized Controlled Trial. Cmaj 2010, 182, E694–E701. [Google Scholar] [CrossRef] [PubMed]
- Wallace, M.S.; Marcotte, T.D.; Umlauf, A.; Gouaux, B.; Atkinson, J.H. Efficacy of Inhaled Cannabis on Painful Diabetic Neuropathy. J. Pain 2015, 16, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Wallace, M.S.; Marcotte, T.D.; Atkinson, J.H.; Padovano, H.T.; Bonn-Miller, M. A Secondary Analysis from a Randomized Trial on the Effect of Plasma Tetrahydrocannabinol Levels on Pain Reduction in Painful Diabetic Peripheral Neuropathy. J. Pain 2020, 21, 1175–1186. [Google Scholar] [CrossRef]
- Andreae, M.H.; Carter, G.M.; Shaparin, N.; Suslov, K.; Ellis, R.J.; Ware, M.A.; Abrams, D.I.; Prasad, H.; Wilsey, B.; Indyk, D.; et al. Inhaled Cannabis for Chronic Neuropathic Pain: A Meta-Analysis of Individual Patient Data. J. Pain 2015, 16, 1221–1232. [Google Scholar] [CrossRef]
- Stockings, E.; Campbell, G.; Hall, W.D.; Nielsen, S.; Zagic, D.; Rahman, R.; Murnion, B.; Farrell, M.; Weier, M.; Degenhardt, L. Cannabis and Cannabinoids for the Treatment of People with Chronic Noncancer Pain Conditions: A Systematic Review and Meta-Analysis of Controlled and Observational Studies. Pain 2018, 159, 1932–1954. [Google Scholar] [CrossRef]
- Solmi, M.; De Toffol, M.; Kim, J.Y.; Choi, M.J.; Stubbs, B.; Thompson, T.; Firth, J.; Miola, A.; Croatto, G.; Baggio, F.; et al. Balancing Risks and Benefits of Cannabis Use: Umbrella Review of Meta-Analyses of Randomised Controlled Trials and Observational Studies. BMJ 2023, 382, e072348. [Google Scholar] [CrossRef] [PubMed]
- Sokolaj, E.; Assareh, N.; Anderson, K.; Aubrey, K.R.; Vaughan, C.W. Cannabis Constituents for Chronic Neuropathic Pain; Reconciling the Clinical and Animal Evidence. J. Neurochem. 2023. [Google Scholar] [CrossRef] [PubMed]
- Gilhus, N.E.; Tzartos, S.; Evoli, A.; Palace, J.; Burns, T.M.; Verschuuren, J.J.G.M. Myasthenia Gravis. Nat. Rev. Dis. Primers 2019, 5, 30. [Google Scholar] [CrossRef] [PubMed]
- Farmakidis, C.; Pasnoor, M.; Dimachkie, M.M.; Barohn, R.J. Treatment of Myasthenia Gravis. Neurol. Clin. 2018, 36, 311–337. [Google Scholar] [CrossRef] [PubMed]
- Van der Kloot, W. Anandamide, a Naturally-Occurring Agonist of the Cannabinoid Receptor, Blocks Adenylate Cyclase at the Frog Neuromuscular Junction. Brain Res. 1994, 649, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Tarasova, E.O.; Khotkina, N.A.; Bogacheva, P.O.; Chernyshev, K.A.; Gaydukov, A.E.; Balezina, O.P. Noncanonical Potentiation of Evoked Quantal Release of Acetylcholine by Cannabinoids Anandamide and 2-Arachidonoylglycerol in Mouse Motor Synapses. Biochem. Moscow Suppl. Ser. A 2021, 15, 395–405. [Google Scholar] [CrossRef]
- Newman, Z.; Malik, P.; Wu, T.-Y.; Ochoa, C.; Watsa, N.; Lindgren, C. Endocannabinoids Mediate Muscarine-Induced Synaptic Depression at the Vertebrate Neuromuscular Junction. Eur. J. Neurosci. 2007, 25, 1619–1630. [Google Scholar] [CrossRef]
- Sánchez-Pastor, E.; Trujillo, X.; Huerta, M.; Andrade, F. Effects of Cannabinoids on Synaptic Transmission in the Frog Neuromuscular Junction. J. Pharmacol. Exp. Ther. 2007, 321, 439–445. [Google Scholar] [CrossRef]
- Silveira, P.E.; Silveira, N.A.; de Cássia Morini, V.; Kushmerick, C.; Naves, L.A. Opposing Effects of Cannabinoids and Vanilloids on Evoked Quantal Release at the Frog Neuromuscular Junction. Neurosci. Lett. 2010, 473, 97–101. [Google Scholar] [CrossRef]
- Turkanis, S.A.; Karler, R. Effects of Delta-9-Tetrahydrocannabinol, 11 -Hydroxy-Delta-9-Tetrahydrocannabinol and Cannabidiol on Neuromuscular Transmission in the Frog. Neuropharmacology 1986, 25, 1273–1278. [Google Scholar] [CrossRef]
- Kumbaraci, N.M.; Nastuk, W.L. Effects of Delta 9-Tetrahydrocannabinol on Excitable Membranes and Neuromuscular Transmission. Mol. Pharmacol. 1980, 17, 344–349. [Google Scholar]
- Ge, D.; Odierna, G.L.; Phillips, W.D. Influence of Cannabinoids upon Nerve-Evoked Skeletal Muscle Contraction. Neurosci. Lett. 2020, 725, 134900. [Google Scholar] [CrossRef] [PubMed]
- Puopolo, T.; Liu, C.; Ma, H.; Seeram, N.P. Inhibitory Effects of Cannabinoids on Acetylcholinesterase and Butyrylcholinesterase Enzyme Activities. Med. Cannabis Cannabinoids 2022, 5, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Oláh, T.; Bodnár, D.; Tóth, A.; Vincze, J.; Fodor, J.; Reischl, B.; Kovács, A.; Ruzsnavszky, O.; Dienes, B.; Szentesi, P.; et al. Cannabinoid Signalling Inhibits Sarcoplasmic Ca2+ Release and Regulates Excitation-Contraction Coupling in Mammalian Skeletal Muscle. J. Physiol. 2016, 594, 7381–7398. [Google Scholar] [CrossRef] [PubMed]
- Singlár, Z.; Ganbat, N.; Szentesi, P.; Osgonsandag, N.; Szabó, L.; Telek, A.; Fodor, J.; Dienes, B.; Gönczi, M.; Csernoch, L.; et al. Genetic Manipulation of CB1 Cannabinoid Receptors Reveals a Role in Maintaining Proper Skeletal Muscle Morphology and Function in Mice. Int. J. Mol. Sci. 2022, 23, 15653. [Google Scholar] [CrossRef] [PubMed]
- Mathis, S.; Tazir, M.; Magy, L.; Duval, F.; Le Masson, G.; Duchesne, M.; Couratier, P.; Ghorab, K.; Solé, G.; Lacoste, I.; et al. History and Current Difficulties in Classifying Inherited Myopathies and Muscular Dystrophies. J. Neurol. Sci. 2018, 384, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Mirouse, V. Evolution and Developmental Functions of the Dystrophin-Associated Protein Complex: Beyond the Idea of a Muscle-Specific Cell Adhesion Complex. Front. Cell Dev. Biol. 2023, 11, 1182524. [Google Scholar] [CrossRef]
- Cyrulnik, S.E.; Fee, R.J.; Batchelder, A.; Kiefel, J.; Goldstein, E.; Hinton, V.J. Cognitive and Adaptive Deficits in Young Children with Duchenne Muscular Dystrophy (DMD). J. Int. Neuropsychol. Soc. 2008, 14, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Adorisio, R.; Mencarelli, E.; Cantarutti, N.; Calvieri, C.; Amato, L.; Cicenia, M.; Silvetti, M.; D’Amico, A.; Grandinetti, M.; Drago, F.; et al. Duchenne Dilated Cardiomyopathy: Cardiac Management from Prevention to Advanced Cardiovascular Therapies. J. Clin. Med. 2020, 9, 3186. [Google Scholar] [CrossRef]
- Himič, V.; Davies, K.E. Evaluating the Potential of Novel Genetic Approaches for the Treatment of Duchenne Muscular Dystrophy. Eur. J. Hum. Genet. 2021, 29, 1369. [Google Scholar] [CrossRef]
- von Maltzahn, J.; Jones, A.E.; Parks, R.J.; Rudnicki, M.A. Pax7 Is Critical for the Normal Function of Satellite Cells in Adult Skeletal Muscle. Proc. Natl. Acad. Sci. USA 2013, 110, 16474–16479. [Google Scholar] [CrossRef] [PubMed]
- Iannotti, F.A.; Pagano, E.; Guardiola, O.; Adinolfi, S.; Saccone, V.; Consalvi, S.; Piscitelli, F.; Gazzerro, E.; Busetto, G.; Carrella, D.; et al. Genetic and Pharmacological Regulation of the Endocannabinoid CB1 Receptor in Duchenne Muscular Dystrophy. Nat. Commun. 2018, 9, 3950. [Google Scholar] [CrossRef] [PubMed]
- Iannotti, F.A.; Silvestri, C.; Mazzarella, E.; Martella, A.; Calvigioni, D.; Piscitelli, F.; Ambrosino, P.; Petrosino, S.; Czifra, G.; Bíró, T.; et al. The Endocannabinoid 2-AG Controls Skeletal Muscle Cell Differentiation via CB1 Receptor-Dependent Inhibition of Kv7 Channels. Proc. Natl. Acad. Sci. USA 2014, 111, E2472–E2481. [Google Scholar] [CrossRef] [PubMed]
- DiMario, J.X.; Uzman, A.; Strohman, R.C. Fiber Regeneration Is Not Persistent in Dystrophic (MDX) Mouse Skeletal Muscle. Dev. Biol. 1991, 148, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Nathan, P.J.; O’Neill, B.V.; Napolitano, A.; Bullmore, E.T. Neuropsychiatric Adverse Effects of Centrally Acting Antiobesity Drugs. CNS Neurosci. Ther. 2011, 17, 490–505. [Google Scholar] [CrossRef] [PubMed]
- Kalkan, H.; Pagano, E.; Paris, D.; Panza, E.; Cuozzo, M.; Moriello, C.; Piscitelli, F.; Abolghasemi, A.; Gazzerro, E.; Silvestri, C.; et al. Targeting Gut Dysbiosis against Inflammation and Impaired Autophagy in Duchenne Muscular Dystrophy. EMBO Mol. Med. 2023, 15, e16225. [Google Scholar] [CrossRef] [PubMed]
- Iannotti, F.A.; Pagano, E.; Moriello, A.S.; Alvino, F.G.; Sorrentino, N.C.; D’Orsi, L.; Gazzerro, E.; Capasso, R.; De Leonibus, E.; De Petrocellis, L.; et al. Effects of Non-Euphoric Plant Cannabinoids on Muscle Quality and Performance of Dystrophic Mdx Mice. Br. J. Pharmacol. 2019, 176, 1568–1584. [Google Scholar] [CrossRef]
- Argenziano, M.; Tortora, C.; Bellini, G.; Di Paola, A.; Punzo, F.; Rossi, F. The Endocannabinoid System in Pediatric Inflammatory and Immune Diseases. Int. J. Mol. Sci. 2019, 20, 5875. [Google Scholar] [CrossRef]
- Mack, A.; Joy, J. Marijuana and pain. In Marijuana as Medicine? The Science Beyond the Controversy; National Academies Press: Washington, DC, USA, 2000. [Google Scholar]
- Ramaekers, J.G.; Kauert, G.; van Ruitenbeek, P.; Theunissen, E.L.; Schneider, E.; Moeller, M.R. High-Potency Marijuana Impairs Executive Function and Inhibitory Motor Control. Neuropsychopharmacology 2006, 31, 2296–2303. [Google Scholar] [CrossRef]
- Freeman, T.P.; Hall, W. Benefits and Risks of Cannabinoids. BMJ 2023, 382, 2113. [Google Scholar] [CrossRef]
- Russo, E.B. Current Therapeutic Cannabis Controversies and Clinical Trial Design Issues. Front. Pharmacol. 2016, 7, 309. [Google Scholar] [CrossRef]
- Lopera, V.; Rodríguez, A.; Amariles, P. Clinical Relevance of Drug Interactions with Cannabis: A Systematic Review. J. Clin. Med. 2022, 11, 1154. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iannotti, F.A. Cannabinoids, Endocannabinoids, and Synthetic Cannabimimetic Molecules in Neuromuscular Disorders. Int. J. Mol. Sci. 2024, 25, 238. https://doi.org/10.3390/ijms25010238
Iannotti FA. Cannabinoids, Endocannabinoids, and Synthetic Cannabimimetic Molecules in Neuromuscular Disorders. International Journal of Molecular Sciences. 2024; 25(1):238. https://doi.org/10.3390/ijms25010238
Chicago/Turabian StyleIannotti, Fabio Arturo. 2024. "Cannabinoids, Endocannabinoids, and Synthetic Cannabimimetic Molecules in Neuromuscular Disorders" International Journal of Molecular Sciences 25, no. 1: 238. https://doi.org/10.3390/ijms25010238
APA StyleIannotti, F. A. (2024). Cannabinoids, Endocannabinoids, and Synthetic Cannabimimetic Molecules in Neuromuscular Disorders. International Journal of Molecular Sciences, 25(1), 238. https://doi.org/10.3390/ijms25010238