

From Pathogenesis to Intervention: The Importance of the Microbiome in Oral Mucositis
 ,
, 
Abstract
1. Introduction
2. The Oral Microbiome and Its Influence on Oral Health
3. The Oral Microbiome and Oral Mucositis: Cause or Consequence?
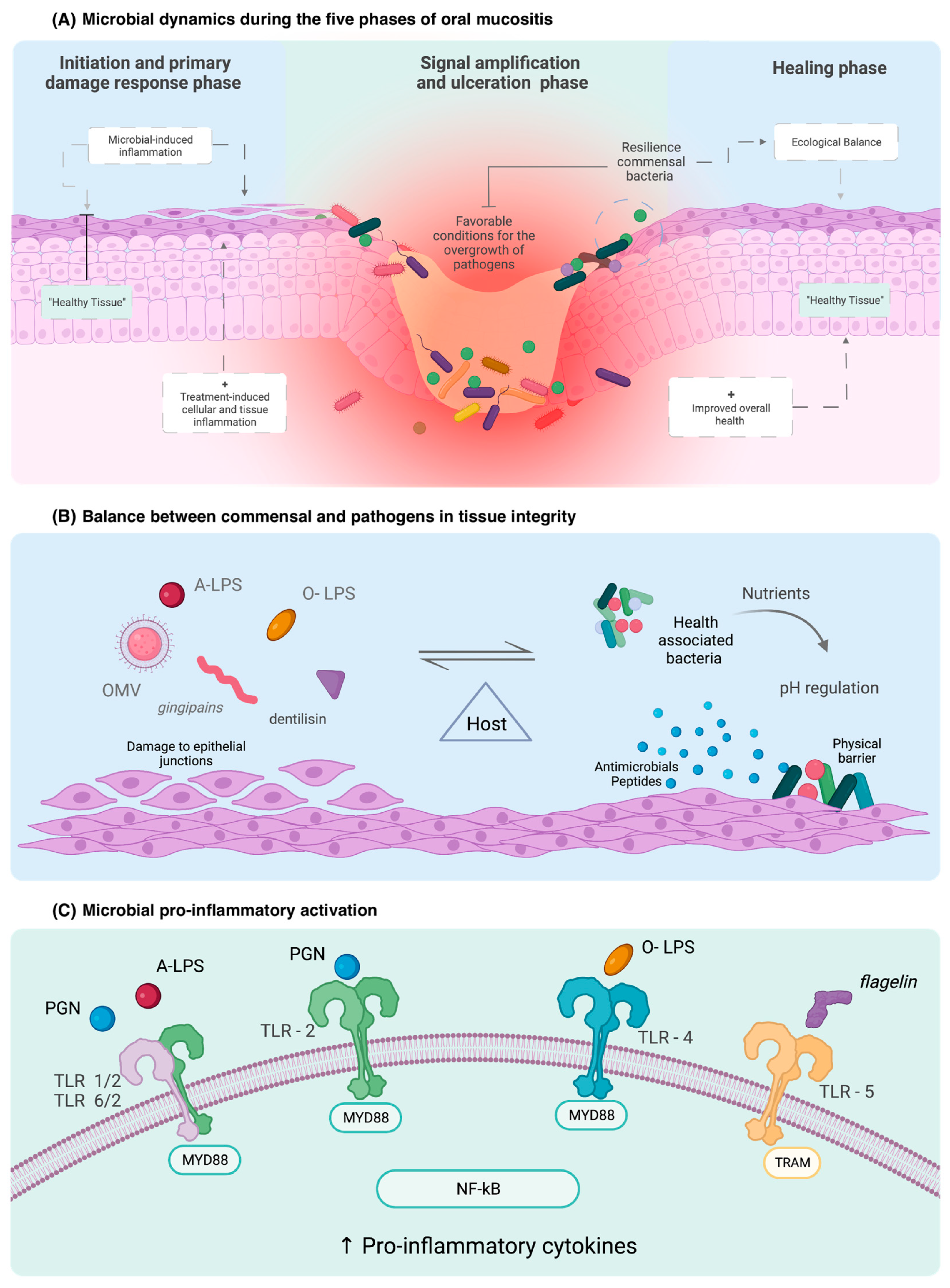
3.1. Dynamic Changes in the Oral Microbiome in the Setting of Oral Mucositis

{kind=link}
| Author | Study Population (n), Treatment | Sample Collection Site | Time Point (s) | Key Findings |
|---|---|---|---|---|
| Zhu et al., 2017 [34] | NPC (n = 41), CRT | Retropharyngeal | During RT (10, 20, 30, 40, 50, 60 and 70 Gy) | Developed ≥ grade 2 OM During severe OM: ↑ Actinobacillus, Mannheimia and Streptobacillus During mild OM: ↑ Enhydrobacter, Schwartzia, Pseudoramibacter, Treponema |
| Vesty et al., 2020 [31] | HNC (n = 19), RT | Unstimulated whole saliva Oral mucosa | Pre-RT and during RT (0–20 Gy, 21–40 Gy and 41–60 Gy) | Developed ≥ grade 2 OM Baseline: ↑ Capnocytophaga, Neisseria, Olsenella, Parviomonas, Tannerella |
| Hou et al., 2018 [30] | HNC (n = 19), CRT | Oropharyngeal mucosa | Pre-RT and during RT (10, 20, 30, 40, 50, 60, and 70 Gy) | Developed ≥ grade 2 OM OM onset: ↑ Prevotella, Fusobacterium, Treponema, Porphyromonas |
| Reyes-Gibby et al., 2020 [35] | HNC (n = 57), CRT | Buccal mucosa | Pre-RT, OM onset and OM ulceration | Onset of OM non-ulcerated ↑ Prevotella, Fusobacterium, Streptococcus Development of severe OM Baseline: ↑ Cardiobacterium, Granulicatella During OM: ↑ Megasphaera, Cardiobacterium |
| Al-Qadami et al., 2023 [36] | HNC (n = 20), CRT | Stool | Pre-treatment | Developed ≥ grade 3 OM Baseline: ↑ Eubacterium, Victivallis, Ruminococcus Developed grade 0–1 OM Baseline: ↑ Alistipes |
| Hong B-Y et al., 2019 [27] | Solid tumours * (n = 49), 5-FU or doxorubicin QT | Unstimulated saliva Oral mucosa | Pre-QT, 3 days, 9 days, 14 days after QT infusion | Developed ≥ grade 2 OM During OM: ↑ Fusobacterium, uncultured Clostridiales, Prevotella, Treponema |
| Laheij et al., 2019 [33] | MM (n = 51), auto-HSCT | Oral cavity rinse | Pre auto-HSCT, 0–4 days, 1 w, 4 w and 3 m after SC infusion | Developed ≥ grade 2 OM Baseline: ↑ Veillonella, Enterococcus, Streptococcus, Staphylococcus, Fusobacterium, Prevotella No OM development Baseline: ↑ Streptococcus, Actinomyces |
| Shouval et al., 2020 [37] | Haematological diseases (n = 604), allo-HSCT | Saliva | Pre allo-HSCT and 1w, 2 w, 3 w, 4 w, 5 w after SC infusion | Developed ≥ grade 0–1 Baseline: ↑ Aggregatibacter, TG5, Lactobacillus, Butyrivibrio, Treponema, Schwartzia, Paludibacter Developed ≥ grade 2–3 Baseline: ↑ Kingella, Atopobium, Haemophilus, Fusobacterium, Corynebacterium, Actinomyces, Cardiobacterium Developed ≥ grade 0–1 During OM: ↑ Filifactor, Selenomonas, Brachymonas, Eikenella, Treponema, TG5 Developed ≥ grade 2–3 During OM: ↑ Methylobacterium |
| Bruno et al., 2022 [38] | Haematological diseases (n = 30), allo-HSCT | Buccal mucosa | Pre allo-HSCT, OM ulceration onset and OM healed | Developed ≥ grade 2 OM OM onset: ↑ Mycoplasma and Lactobacillus Post OM: ↑ Staphylococcus, Treponema 2, Enterococcus, Lactobacillus |
| Laheij et al., 2022 [28] | Haematological diseases (n = 50), allo-HSCT | Oral cavity rinse | Pre-SCT and 3 m, 6 m, 12 m and 18 m after SC infusion | Developed ≥ grade 2 OM OM onset: ↑ Mycobacterium, Staphylococcus, Enterococcus |
3.2. Evidence of Causal Involvement of the Oral Microbiome in Oral Mucositis
3.3. Mechanisms by Which the Microbiome Contributes to Oral Mucositis
4. Clinical Applications of the Microbiome in OM
4.1. Microbial Fingerprints as Biomarkers for Oral Mucositis
4.2. Microbial Therapeutics for OM Prevention and Treatment
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sonis, S.T. The pathobiology of mucositis. Nat. Rev. Cancer 2004, 4, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Villa, A.; Sonis, S.T. Pharmacotherapy for the management of cancer regimen-related oral mucositis. Expert Opin. Pharmacother. 2016, 17, 1801–1807. [Google Scholar] [CrossRef] [PubMed]
- Sonis, S.T.; Elting, L.S.; Keefe, D.; Peterson, D.E.; Schubert, M.; Hauer-Jensen, M.; Bekele, B.N.; Raber-Durlacher, J.; Donnelly, J.P.; Rubenstein, E.B. Perspectives on cancer therapy-induced mucosal injury: Pathogenesis, measurement, epidemiology, and consequences for patients. Cancer 2004, 100, 1995–2025. [Google Scholar] [CrossRef] [PubMed]
- Elad, S.; Cheng, K.K.F.; Lalla, R.v.; Yarom, N.; Hong, C.; Logan, R.M.; Bowen, J.; Gibson, R.; Saunders, D.P.; Zadik, Y.; et al. MASCC/ISOO clinical practice guidelines for the management of mucositis secondary to cancer therapy. Cancer 2020, 126, 4423–4431. [Google Scholar] [CrossRef]
- Cinausero, M.; Aprile, G.; Ermacora, P.; Basile, D.; Vitale, M.G.; Fanotto, V.; Parisi, G.; Calvetti, L.; Sonis, S.T. New Frontiers in the Pathobiology and Treatment of Cancer Regimen-Related Mucosal Injury. Front. Pharmacol. 2017, 8, 354. [Google Scholar] [CrossRef]
- Wardill, H.R.; Sonis, S.T.; Blijlevens, N.M.A.; van Sebille, Y.Z.A.; Ciorba, M.A.; Loeffen, E.A.H.; Cheng, K.K.F.; Bossi, P.; Porcello, L.; Castillo, D.A.; et al. Prediction of mucositis risk secondary to cancer therapy: A systematic review of current evidence and call to action. Support. Care Cancer 2020, 28, 5059–5073. [Google Scholar] [CrossRef]
- Deo, P.N.; Deshmukh, R. Oral microbiome: Unveiling the fundamentals. J. Oral Maxillofac. Pathol. 2019, 23, 122–128. [Google Scholar] [CrossRef]
- Vasconcelos, R.M.; Sanfilippo, N.; Paster, B.J.; Kerr, A.R.; Li, Y.; Ramalho, L.; Queiroz, E.L.; Smith, B.; Sonis, S.T.; Corby, P.M. Host-Microbiome Cross-talk in Oral Mucositis. J. Dent. Res. 2016, 95, 725–733. [Google Scholar] [CrossRef]
- Maki, K.A.; Kazmi, N.; Barb, J.J.; Ames, N. The Oral and Gut Bacterial Microbiomes: Similarities, Differences, and Connections. Biol. Res. Nurs. 2021, 23, 7–20. [Google Scholar] [CrossRef]
- Kilian, M.; Chapple, I.L.C.; Hannig, M.; Marsh, P.D.; Meuric, V.; Pedersen, A.M.L.; Tonetti, M.S.; Wade, W.G.; Zaura, E. The oral microbiome—An update for oral healthcare professionals. Br. Dent. J. 2016, 221, 657–666. [Google Scholar] [CrossRef]
- Arweiler, N.B.; Netuschil, L. The Oral Microbiota. Adv. Exp. Med. Biol. 2016, 902, 45–60. [Google Scholar] [PubMed]
- Caselli, E.; Fabbri, C.; D’Accolti, M.; Soffritti, I.; Bassi, C.; Mazzacane, S.; Franchi, M. Defining the oral microbiome by whole-genome sequencing and resistome analysis: The complexity of the healthy picture. BMC Microbiol. 2020, 20, 120. [Google Scholar] [CrossRef] [PubMed]
- Willis, J.R.; Gabaldón, T. The Human Oral Microbiome in Health and Disease: From Sequences to Ecosystems. Microorganisms 2020, 8, 308. [Google Scholar] [CrossRef]
- Belibasakis, G.N.; Bostanci, N.; Marsh, P.D.; Zaura, E. Applications of the oral microbiome in personalized dentistry. Arch. Oral Biol. 2019, 104, 7–12. [Google Scholar] [CrossRef]
- Mark Welch, J.L.; Ramírez-Puebla, S.T.; Borisy, G.G. Oral Microbiome Geography: Micron-Scale Habitat and Niche. Cell Host Microbe 2020, 28, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Radaic, A.; Ganther, S.; Kamarajan, P.; Grandis, J.; Yom, S.S.; Kapila, Y.L. Paradigm shift in the pathogenesis and treatment of oral cancer and other cancers focused on the oralome and antimicrobial-based therapeutics. Periodontology 2000 2021, 87, 76–93. [Google Scholar] [CrossRef]
- Şenel, S. An Overview of Physical, Microbiological and Immune Barriers of Oral Mucosa. Int. J. Mol. Sci. 2021, 22, 7821. [Google Scholar] [CrossRef]
- Shang, L.; Deng, D.; Buskermolen, J.K.; Janus, M.M.; Krom, B.P.; Roffel, S.; Waaijman, T.; van Loveren, C.; Crielaard, W.; Gibbs, S. Multi-species oral biofilm promotes reconstructed human gingiva epithelial barrier function. Sci. Rep. 2018, 8, 16061. [Google Scholar] [CrossRef]
- Wade, W.G. The oral microbiome in health and disease. Pharmacol. Res. 2013, 69, 137–143. [Google Scholar] [CrossRef]
- Rosier, B.T.; Moya-Gonzalvez, E.M.; Corell-Escuin, P.; Mira, A. Isolation and Characterization of Nitrate-Reducing Bacteria as Potential Probiotics for Oral and Systemic Health. Front. Microbiol. 2020, 11, 555465. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Carlström, M.; Weitzberg, E. Metabolic Effects of Dietary Nitrate in Health and Disease. Cell Metab. 2018, 28, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Enersen, M.; Nakano, K.; Amano, A. Porphyromonas gingivalis fimbriae. J. Oral Microbiol. 2013, 5, 20265. [Google Scholar] [CrossRef] [PubMed]
- Groeger, S.; Meyle, J. Oral Mucosal Epithelial Cells. Front. Immunol. 2019, 10, 208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, D.; Liu, S.; Zhang, S.; Pan, Y. The Role of Porphyromonas gingivalis Outer Membrane Vesicles in Periodontal Disease and Related Systemic Diseases. Front. Cell. Infect. Microbiol. 2020, 10, 585917. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Naito, M.; Omori, K.; Ono, S.; Nakayama, K.; Ohara, N. Porphyromonas gingivalis Gingipains Induce Cyclooxygenase-2 Expression and Prostaglandin E(2) Production via ERK1/2-Activated AP-1 (c-Jun/c-Fos) and IKK/NF-κB p65 Cascades. J. Immunol. 2022, 208, 1146–1154. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.A.; Garrett, W.S. Fusobacterium nucleatum—Symbiont, opportunist and oncobacterium. Nat. Rev. Microbiol. 2019, 17, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Hong, B.-Y.; Sobue, T.; Choquette, L.; Dupuy, A.K.; Thompson, A.; Burleson, J.A.; Salner, A.L.; Schauer, P.K.; Joshi, P.; Fox, E.; et al. Chemotherapy-induced oral mucositis is associated with detrimental bacterial dysbiosis. Microbiome 2019, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Laheij, A.M.G.A.; Rozema, F.R.; Brennan, M.T.; von Bültzingslöwen, I.; van Leeuwen, S.J.M.; Potting, C.; Huysmans, M.-C.D.N.J.M.; Hazenberg, M.D.; Brandt, B.W.; Zaura, E.; et al. Long-Term Analysis of Resilience of the Oral Microbiome in Allogeneic Stem Cell Transplant Recipients. Microorganisms 2022, 10, 734. [Google Scholar] [CrossRef]
- Mojdami, Z.D.; Barbour, A.; Oveisi, M.; Sun, C.; Fine, N.; Saha, S.; Marks, C.; Elebyary, O.; Watson, E.; Tenenbaum, H.; et al. The Effect of Intensity-Modulated Radiotherapy to the Head and Neck Region on the Oral Innate Immune Response and Oral Microbiome: A Prospective Cohort Study of Head and Neck Tumour Patients. Int. J. Mol. Sci. 2022, 23, 9594. [Google Scholar] [CrossRef]
- Hou, J.; Zheng, H.M.; Li, P.; Liu, H.Y.; Zhou, H.W.; Yang, X.J. Distinct shifts in the oral microbiota are associated with the progression and aggravation of mucositis during radiotherapy. Radiother. Oncol. 2018, 129, 44–51. [Google Scholar] [CrossRef]
- Vesty, A.; Gear, K.; Biswas, K.; Mackenzie, B.W.; Taylor, M.W.; Douglas, R.G. Oral microbial influences on oral mucositis during radiotherapy treatment of head and neck cancer. Support. Care Cancer 2020, 28, 2683–2691. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Toyosaki, M.; Matsui, K.; Machida, S.; Kikkawa, E.; Ota, Y.; Kaneko, A.; Ogawa, Y.; Ando, K.; Onizuka, M. An analysis of oral microbial flora by T-RFLP in patients undergoing hematopoietic stem cell transplantation. Int. J. Hematol. 2020, 112, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Laheij, A.M.G.A.; Raber-Durlacher, J.E.; Koppelmans, R.G.A.; Huysmans, M.-C.D.N.J.M.; Potting, C.; van Leeuwen, S.J.M.; Hazenberg, M.D.; Brennan, M.T.; von Bültzingslöwen, I.; Johansson, J.-E.; et al. Microbial changes in relation to oral mucositis in autologous hematopoietic stem cell transplantation recipients. Sci. Rep. 2019, 9, 16929. [Google Scholar] [CrossRef]
- Zhu, X.-X.; Yang, X.-J.; Chao, Y.-L.; Zheng, H.-M.; Sheng, H.-F.; Liu, H.-Y.; He, Y.; Zhou, H.-W. The Potential Effect of Oral Microbiota in the Prediction of Mucositis During Radiotherapy for Nasopharyngeal Carcinoma. EBioMedicine 2017, 18, 23–31. [Google Scholar] [CrossRef]
- Reyes-Gibby, C.C.; Wang, J.; Zhang, L.; Peterson, C.B.; Do, K.-A.; Jenq, R.R.; Shelburne, S.; Shah, D.P.; Chambers, M.S.; Hanna, E.Y.; et al. Oral microbiome and onset of oral mucositis in patients with squamous cell carcinoma of the head and neck. Cancer 2020, 126, 5124–5136. [Google Scholar] [CrossRef] [PubMed]
- Al-Qadami, G.; Bowen, J.; van Sebille, Y.; Secombe, K.; Dorraki, M.; Verjans, J.; Wardill, H.; Le, H. Baseline gut microbiota composition is associated with oral mucositis and tumour recurrence in patients with head and neck cancer: A pilot study. Support. Care Cancer 2023, 31, 98. [Google Scholar] [CrossRef] [PubMed]
- Shouval, R.; Eshel, A.; Dubovski, B.; Kuperman, A.A.; Danylesko, I.; Fein, J.A.; Fried, S.; Geva, M.; Kouniavski, E.; Neuman, H.; et al. Patterns of salivary microbiota injury and oral mucositis in recipients of allogeneic hematopoietic stem cell transplantation. Blood Adv. 2020, 4, 2912–2917. [Google Scholar] [CrossRef]
- Bruno, J.S.; Heidrich, V.; Knebel, F.H.; de Molla, V.C.; Parahyba, C.J.; Miranda-Silva, W.; Asprino, P.F.; Tucunduva, L.; Rocha, V.; Novis, Y.; et al. Commensal oral microbiota impacts ulcerative oral mucositis clinical course in allogeneic stem cell transplant recipients. Sci. Rep. 2022, 12, 17527. [Google Scholar] [CrossRef]
- Gupta, N.; Quah, S.Y.; Yeo, J.F.; Ferreira, J.; Tan, K.S.; Hong, C.H.L. Role of oral flora in chemotherapy-induced oral mucositis in vivo. Arch. Oral Biol. 2019, 101, 51–56. [Google Scholar] [CrossRef]
- Al-Qadami, G.; van Sebille, Y.; Bowen, J.; Wardill, H. Oral-Gut Microbiome Axis in the Pathogenesis of Cancer Treatment-Induced Oral Mucositis. Front. Oral Health 2022, 3, 881949. [Google Scholar] [CrossRef]
- Gheorghe, C.E.; Ritz, N.L.; Martin, J.A.; Wardill, H.R.; Cryan, J.F.; Clarke, G. Investigating causality with fecal microbiota transplantation in rodents: Applications, recommendations and pitfalls. Gut Microbes 2021, 13, 1941711. [Google Scholar] [CrossRef] [PubMed]
- Al-Qadami, G.; Verma, G.; van Sebille, Y.; Le, H.; Hewson, I.; Bateman, E.; Wardill, H.; Bowen, J. Antibiotic-Induced Gut Microbiota Depletion Accelerates the Recovery of Radiation-Induced Oral Mucositis in Rats. Int. J. Radiat. Oncol. Biol. Phys. 2022, 113, 845–858. [Google Scholar] [CrossRef]
- Wardill, H.R.; van der Aa, S.A.R.; da Silva Ferreira, A.R.; Havinga, R.; Tissing, W.J.E.; Harmsen, H.J.M. Antibiotic-induced disruption of the microbiome exacerbates chemotherapy-induced diarrhoea and can be mitigated with autologous faecal microbiota transplantation. Eur. J. Cancer 2021, 153, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Ma, Y.; Peng, X.; Qiu, W.; Kong, L.; Ren, B.; Li, M.; Cheng, G.; Zhou, X.; Cheng, L. Antibiotic-induced dysbiosis of the rat oral and gut microbiota and resistance to Salmonella. Arch. Oral Biol. 2020, 114, 104730. [Google Scholar] [CrossRef]
- Stokman, M.A.; Spijkervet, F.K.L.; Burlage, F.R.; Dijkstra, P.U.; Manson, W.L.; de Vries, E.G.E.; Roodenburg, J.L.N. Oral mucositis and selective elimination of oral flora in head and neck cancer patients receiving radiotherapy: A double-blind randomised clinical trial. Br. J. Cancer 2003, 88, 1012–1016. [Google Scholar] [CrossRef]
- Gupta, N.; Ferreira, J.; Hong, C.H.L.; Tan, K.S. Lactobacillus reuteri DSM 17938 and ATCC PTA 5289 ameliorates chemotherapy-induced oral mucositis. Sci. Rep. 2020, 10, 16189. [Google Scholar] [CrossRef]
- Xia, C.; Jiang, C.; Li, W.; Wei, J.; Hong, H.; Li, J.; Feng, L.; Wei, H.; Xin, H.; Chen, T. A Phase II Randomized Clinical Trial and Mechanistic Studies Using Improved Probiotics to Prevent Oral Mucositis Induced by Concurrent Radiotherapy and Chemotherapy in Nasopharyngeal Carcinoma. Front. Immunol. 2021, 12, 618150. [Google Scholar] [CrossRef]
- Xiao, H.; Fan, Y.; Li, Y.; Dong, J.; Zhang, S.; Wang, B.; Liu, J.; Liu, X.; Fan, S.; Guan, J.; et al. Oral microbiota transplantation fights against head and neck radiotherapy-induced oral mucositis in mice. Comput. Struct. Biotechnol. J. 2021, 19, 5898–5910. [Google Scholar] [CrossRef]
- Pouncey, A.L.; Scott, A.J.; Alexander, J.L.; Marchesi, J.; Kinross, J. Gut microbiota, chemotherapy and the host: The influence of the gut microbiota on cancer treatment. Ecancermedicalscience 2018, 12, 868. [Google Scholar] [CrossRef]
- Alexander, J.L.; Wilson, I.D.; Teare, J.; Marchesi, J.R.; Nicholson, J.K.; Kinross, J.M. Gut microbiota modulation of chemotherapy efficacy and toxicity. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 356–365. [Google Scholar] [CrossRef]
- Sonis, S.T. The Chicken or the Egg? Changes in Oral Microbiota as Cause or Consequence of Mucositis During Radiation Therapy. EBioMedicine 2017, 18, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Pedroso, S.H.S.P.; Vieira, A.T.; Bastos, R.W.; Oliveira, J.S.; Cartelle, C.T.; Arantes, R.M.E.; Soares, P.M.G.; Generoso, S.v.; Cardoso, V.N.; Teixeira, M.M.; et al. Evaluation of mucositis induced by irinotecan after microbial colonization in germ-free mice. Microbiology 2015, 161, 1950–1960. [Google Scholar] [CrossRef] [PubMed]
- Wardill, H.R.; Gibson, R.J.; van Sebille, Y.Z.A.; Secombe, K.R.; Coller, J.K.; White, I.A.; Manavis, J.; Hutchinson, M.R.; Staikopoulos, V.; Logan, R.M.; et al. Irinotecan-Induced Gastrointestinal Dysfunction and Pain Are Mediated by Common TLR4-Dependent Mechanisms. Mol. Cancer Ther. 2016, 15, 1376–1386. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Hao, S.; Wang, J.; Zou, J.; Wang, Y. Roles of Toll-Like Receptors in Radiotherapy- and Chemotherapy-Induced Oral Mucositis: A Concise Review. Front. Cell. Infect. Microbiol. 2022, 12, 831387. [Google Scholar] [CrossRef]
- Moutsopoulos, N.M.; Konkel, J.E. Tissue-Specific Immunity at the Oral Mucosal Barrier. Trends Immunol. 2018, 39, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Suárez, L.J.; Arboleda, S.; Angelov, N.; Arce, R.M. Oral versus Gastrointestinal Mucosal Immune Niches in Homeostasis and Allostasis. Front. Immunol. 2021, 12, 705206. [Google Scholar] [CrossRef]
- Sonis, S.T. Pathobiology of oral mucositis: Novel insights and opportunities. J. Support. Oncol. 2007, 5, 3–11. [Google Scholar]
- Li, D.; Wu, M. Pattern recognition receptors in health and diseases. Signal Transduct. Target Ther. 2021, 6, 291. [Google Scholar] [CrossRef]
- Takeuchi, H.; Sasaki, N.; Yamaga, S.; Kuboniwa, M.; Matsusaki, M.; Amano, A. Porphyromonas gingivalis induces penetration of lipopolysaccharide and peptidoglycan through the gingival epithelium via degradation of junctional adhesion molecule 1. PLoS Pathog 2019, 15, e1008124. [Google Scholar] [CrossRef]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef]
- Feuillet, V.; Medjane, S.; Mondor, I.; Demaria, O.; Pagni, P.P.; Galán, J.E.; Flavell, R.A.; Alexopoulou, L. Involvement of Toll-like receptor 5 in the recognition of flagellated bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 12487–12492. [Google Scholar] [CrossRef]
- Hall, M.W.; Singh, N.; Ng, K.F.; Lam, D.K.; Goldberg, M.B.; Tenenbaum, H.C.; Neufeld, J.D.; Beiko, R.G.; Senadheera, D.B. Inter-personal diversity and temporal dynamics of dental, tongue, and salivary microbiota in the healthy oral cavity. NPJ Biofilms Microbiomes 2017, 3, 2. [Google Scholar] [CrossRef]
- Huang, K.; Gao, X.; Wu, L.; Yan, B.; Wang, Z.; Zhang, X.; Peng, L.; Yu, J.; Sun, G.; Yang, Y. Salivary Microbiota for Gastric Cancer Prediction: An Exploratory Study. Front. Cell. Infect. Microbiol. 2021, 11, 640309. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Alekseyenko, A.v.; Wu, J.; Peters, B.A.; Jacobs, E.J.; Gapstur, S.M.; Purdue, M.P.; Abnet, C.C.; Stolzenberg-Solomon, R.; Miller, G.; et al. Human oral microbiome and prospective risk for pancreatic cancer: A population-based nested case-control study. Gut 2018, 67, 120–127. [Google Scholar] [CrossRef]
- Flemer, B.; Warren, R.D.; Barrett, M.P.; Cisek, K.; Das, A.; Jeffery, I.B.; Hurley, E.; O’Riordain, M.; Shanahan, F.; O’Toole, P.W. The oral microbiota in colorectal cancer is distinctive and predictive. Gut 2018, 67, 1454–1463. [Google Scholar] [CrossRef] [PubMed]
- Grier, A.; Myers, J.A.; O’Connor, T.G.; Quivey, R.G.; Gill, S.R.; Kopycka-Kedzierawski, D.T. Oral Microbiota Composition Predicts Early Childhood Caries Onset. J. Dent. Res. 2021, 100, 599–607. [Google Scholar] [CrossRef]
- Lim, Y.; Fukuma, N.; Totsika, M.; Kenny, L.; Morrison, M.; Punyadeera, C. The Performance of an Oral Microbiome Biomarker Panel in Predicting Oral Cavity and Oropharyngeal Cancers. Front. Cell. Infect. Microbiol. 2018, 8, 267. [Google Scholar] [CrossRef]
- Jiang, R.; Liu, Y.; Zhang, H.; Chen, Y.; Liu, T.; Zeng, J.; Nie, E.; Chen, S.; Tan, J. Distinctive microbiota of delayed healing of oral mucositis after radiotherapy of nasopharyngeal carcinoma. Front. Cell. Infect. Microbiol. 2022, 12, 1070322. [Google Scholar] [CrossRef]
- Zaura, E.; Brandt, B.W.; Prodan, A.; Teixeira de Mattos, M.J.; Imangaliyev, S.; Kool, J.; Buijs, M.J.; Jagers, F.L.; Hennequin-Hoenderdos, N.L.; Slot, D.E.; et al. On the ecosystemic network of saliva in healthy young adults. ISME J. 2017, 11, 1218–1231. [Google Scholar] [CrossRef]
- Zaura, E. A Commentary on the Potential Use of Oral Microbiome in Prediction, Diagnosis or Prognostics of a Distant Pathology. Dent. J. 2022, 10, 156. [Google Scholar] [CrossRef]
- Veziant, J.; Villéger, R.; Barnich, N.; Bonnet, M. Gut Microbiota as Potential Biomarker and/or Therapeutic Target to Improve the Management of Cancer: Focus on Colibactin-Producing Escherichia coli in Colorectal Cancer. Cancers 2021, 13, 2215. [Google Scholar] [CrossRef] [PubMed]
- Metwaly, A.; Reitmeier, S.; Haller, D. Microbiome risk profiles as biomarkers for inflammatory and metabolic disorders. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 383–397. [Google Scholar] [CrossRef]
- Brookes, Z.L.S.; Bescos, R.; Belfield, L.A.; Ali, K.; Roberts, A. Current uses of chlorhexidine for management of oral disease: A narrative review. J. Dent. 2020, 103, 103497. [Google Scholar] [CrossRef]
- Laheij, A.M.G.A.; de Soet, J.J. Can the oral microflora affect oral ulcerative mucositis? Curr. Opin. Support. Palliat. Care 2014, 8, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-F.; Liu, S.-P.; Muo, C.-H.; Chen, H.-J.; Liao, Y.-F.; Chiu, K.-M.; Chen, C.-C.; Tsai, C.-H.; Hong, H.-H.; Chang, C.-T. The impact of timing and modalities of dental prophylaxis on the risk of 5-fluorouracil-related oral mucositis in patients with head and neck cancer: A nationwide population-based cohort study. Support. Care Cancer 2021, 29, 3163–3171. [Google Scholar] [CrossRef] [PubMed]
- Saunders, D.P.; Rouleau, T.; Cheng, K.; Yarom, N.; Kandwal, A.; Joy, J.; Bektas Kayhan, K.; van de Wetering, M.; Brito-Dellan, N.; Kataoka, T.; et al. Systematic review of antimicrobials, mucosal coating agents, anesthetics, and analgesics for the management of oral mucositis in cancer patients and clinical practice guidelines. Support. Care Cancer 2020, 28, 2473–2484. [Google Scholar] [CrossRef]
- Hong, C.H.L.; Gueiros, L.A.; Fulton, J.S.; Cheng, K.K.F.; Kandwal, A.; Galiti, D.; Fall-Dickson, J.M.; Johansen, J.; Ameringer, S.; Kataoka, T.; et al. Systematic review of basic oral care for the management of oral mucositis in cancer patients and clinical practice guidelines. Support. Care Cancer 2019, 27, 3949–3967. [Google Scholar] [CrossRef]
- Chatzigiannidou, I.; Teughels, W.; van de Wiele, T.; Boon, N. Oral biofilms exposure to chlorhexidine results in altered microbial composition and metabolic profile. NPJ Biofilms Microbiomes 2020, 6, 13. [Google Scholar] [CrossRef]
- Bescos, R.; Ashworth, A.; Cutler, C.; Brookes, Z.L.; Belfield, L.; Rodiles, A.; Casas-Agustench, P.; Farnham, G.; Liddle, L.; Burleigh, M.; et al. Effects of Chlorhexidine mouthwash on the oral microbiome. Sci. Rep. 2020, 10, 5254. [Google Scholar] [CrossRef]
- Wong, A.C.; Levy, M. New Approaches to Microbiome-Based Therapies. mSystems 2019, 4, e00122-19. [Google Scholar] [CrossRef]
- Butera, A.; Pascadopoli, M.; Pellegrini, M.; Gallo, S.; Zampetti, P.; Scribante, A. Oral Microbiota in Patients with Peri-Implant Disease: A Narrative Review. Appl. Sci. 2022, 12, 3250. [Google Scholar] [CrossRef]
- Butera, A.; Folini, E.; Cosola, S.; Russo, G.; Scribante, A.; Gallo, S.; Stablum, G.; Menchini Fabris, G.B.; Covani, U.; Genovesi, A. Evaluation of the Efficacy of Probiotics Domiciliary Protocols for the Management of Periodontal Disease, in Adjunction of Non-Surgical Periodontal Therapy (NSPT): A Systematic Literature Review. Appl. Sci. 2023, 13, 663. [Google Scholar] [CrossRef]
- Scribante, A.; Gallo, S.; Pascadopoli, M.; Soleo, R.; Di Fonso, F.; Politi, L.; Venugopal, A.; Marya, A.; Butera, A. Management of Periodontal Disease with Adjunctive Therapy with Ozone and Photobiomodulation (PBM): A Randomized Clinical Trial. Photonics 2022, 9, 138. [Google Scholar] [CrossRef]
- Jiang, C.; Wang, H.; Xia, C.; Dong, Q.; Chen, E.; Qiu, Y.; Su, Y.; Xie, H.; Zeng, L.; Kuang, J.; et al. A randomized, double-blind, placebo-controlled trial of probiotics to reduce the severity of oral mucositis induced by chemoradiotherapy for patients with nasopharyngeal carcinoma. Cancer 2019, 125, 1081–1090. [Google Scholar] [CrossRef]
- Mirza, M.A.; Aruna, D.; Irukulla, M. Efficacy of Bacillus clausii UBBC—07 spores in the amelioration of oral mucositis in head and neck cancer patients undergoing radiation therapy. Cancer Treat Res. Commun. 2022, 31, 100523. [Google Scholar] [CrossRef]
- de Sanctis, V.; Belgioia, L.; Cante, D.; la Porta, M.R.; Caspiani, O.; Guarnaccia, R.; Argenone, A.; Muto, P.; Musio, D.; de Felice, F.; et al. Lactobacillus brevis CD2 for Prevention of Oral Mucositis in Patients with Head and Neck Tumors: A Multicentric Randomized Study. Anticancer Res. 2019, 39, 1935–1942. [Google Scholar] [CrossRef]
- Liu, Y.-C.; Wu, C.-R.; Huang, T.-W. Preventive Effect of Probiotics on Oral Mucositis Induced by Cancer Treatment: A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2022, 23, 13268. [Google Scholar] [CrossRef]
- López-López, A.; Camelo-Castillo, A.; Ferrer, M.D.; Simon-Soro, Á.; Mira, A. Health-Associated Niche Inhabitants as Oral Probiotics: The Case of Streptococcus dentisani. Front. Microbiol. 2017, 8, 379. [Google Scholar] [CrossRef]
- Ferrer, M.D.; López-López, A.; Nicolescu, T.; Perez-Vilaplana, S.; Boix-Amorós, A.; Dzidic, M.; Garcia, S.; Artacho, A.; Llena, C.; Mira, A. Topic Application of the Probiotic Streptococcus dentisani Improves Clinical and Microbiological Parameters Associated With Oral Health. Front. Cell. Infect. Microbiol. 2020, 10, 465. [Google Scholar] [CrossRef]
- Cosseau, C.; Devine, D.A.; Dullaghan, E.; Gardy, J.L.; Chikatamarla, A.; Gellatly, S.; Yu, L.L.; Pistolic, J.; Falsafi, R.; Tagg, J.; et al. The commensal Streptococcus salivarius K12 downregulates the innate immune responses of human epithelial cells and promotes host-microbe homeostasis. Infect. Immun. 2008, 76, 4163–4175. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Zhang, H.; Zheng, X.; Wang, J.; Jia, X.; Peng, X.; Xie, Q.; Zou, J.; Zheng, L.; et al. Probiotic Streptococcus salivarius K12 Alleviates Radiation-Induced Oral Mucositis in Mice. Front. Immunol. 2021, 12, 684824. [Google Scholar] [CrossRef]
- Nath, S.; Zilm, P.; Jamieson, L.; Kapellas, K.; Goswami, N.; Ketagoda, K.; Weyrich, L.S. Development and characterization of an oral microbiome transplant among Australians for the treatment of dental caries and periodontal disease: A study protocol. PLoS ONE 2021, 16, e0260433. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]

| Exogenous Ligands (Responsible for the Recognition) | Toll-like Receptor | Intracellular Adaptor Proteins | Signalling Pathways |
|---|---|---|---|
| Peptidoglycan (PGN) (G+) Glycolipid LAM (Mycoplasma) Peptideo PSMs (Staphylococcus) | 2 | myD88 | Inflammatory cytokines |
| O-LPS (G-) Lipoteichoic acid (G+) | 4 | TIRAP → myD88 or TRAM | Inflammatory cytokines or Inflammatory cytokines IFN |
| Flagelin (Treponema) | 5 | myD88 | Inflammatory cytokines |
| A-LPS (Porphyromonas) Peptidoglycan (PGN) (G+) Glycolipid LAM (Mycoplasma) Peptideo PSMs (Staphylococcus) | 1/2 * | myD88 | Inflammatory cytokines |
| 2/6 * | myD88 | Inflammatory cytokines |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruno, J.S.; Al-Qadami, G.H.; Laheij, A.M.G.A.; Bossi, P.; Fregnani, E.R.; Wardill, H.R. From Pathogenesis to Intervention: The Importance of the Microbiome in Oral Mucositis. Int. J. Mol. Sci. 2023, 24, 8274. https://doi.org/10.3390/ijms24098274
Bruno JS, Al-Qadami GH, Laheij AMGA, Bossi P, Fregnani ER, Wardill HR. From Pathogenesis to Intervention: The Importance of the Microbiome in Oral Mucositis. International Journal of Molecular Sciences. 2023; 24(9):8274. https://doi.org/10.3390/ijms24098274
Chicago/Turabian StyleBruno, Julia S., Ghanyah H. Al-Qadami, Alexa M. G. A. Laheij, Paolo Bossi, Eduardo R. Fregnani, and Hannah R. Wardill. 2023. "From Pathogenesis to Intervention: The Importance of the Microbiome in Oral Mucositis" International Journal of Molecular Sciences 24, no. 9: 8274. https://doi.org/10.3390/ijms24098274
APA StyleBruno, J. S., Al-Qadami, G. H., Laheij, A. M. G. A., Bossi, P., Fregnani, E. R., & Wardill, H. R. (2023). From Pathogenesis to Intervention: The Importance of the Microbiome in Oral Mucositis. International Journal of Molecular Sciences, 24(9), 8274. https://doi.org/10.3390/ijms24098274