
ELMO1 Deficiency Reduces Neutrophil Chemotaxis in Murine Peritonitis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
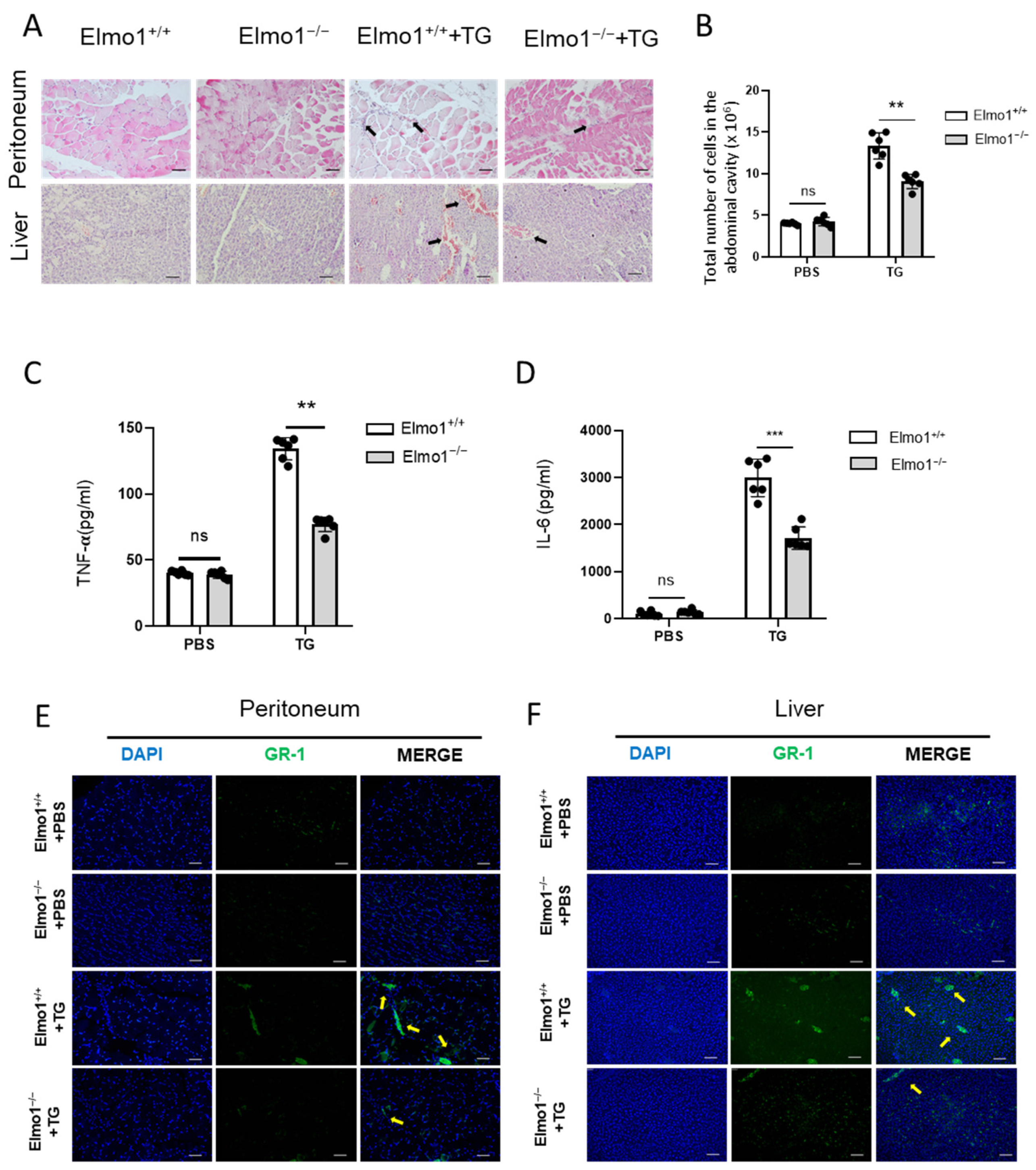
2.1. ELMO1 Plays a Role in Acute Peritonitis by Regulating the Infiltration of Neutrophils
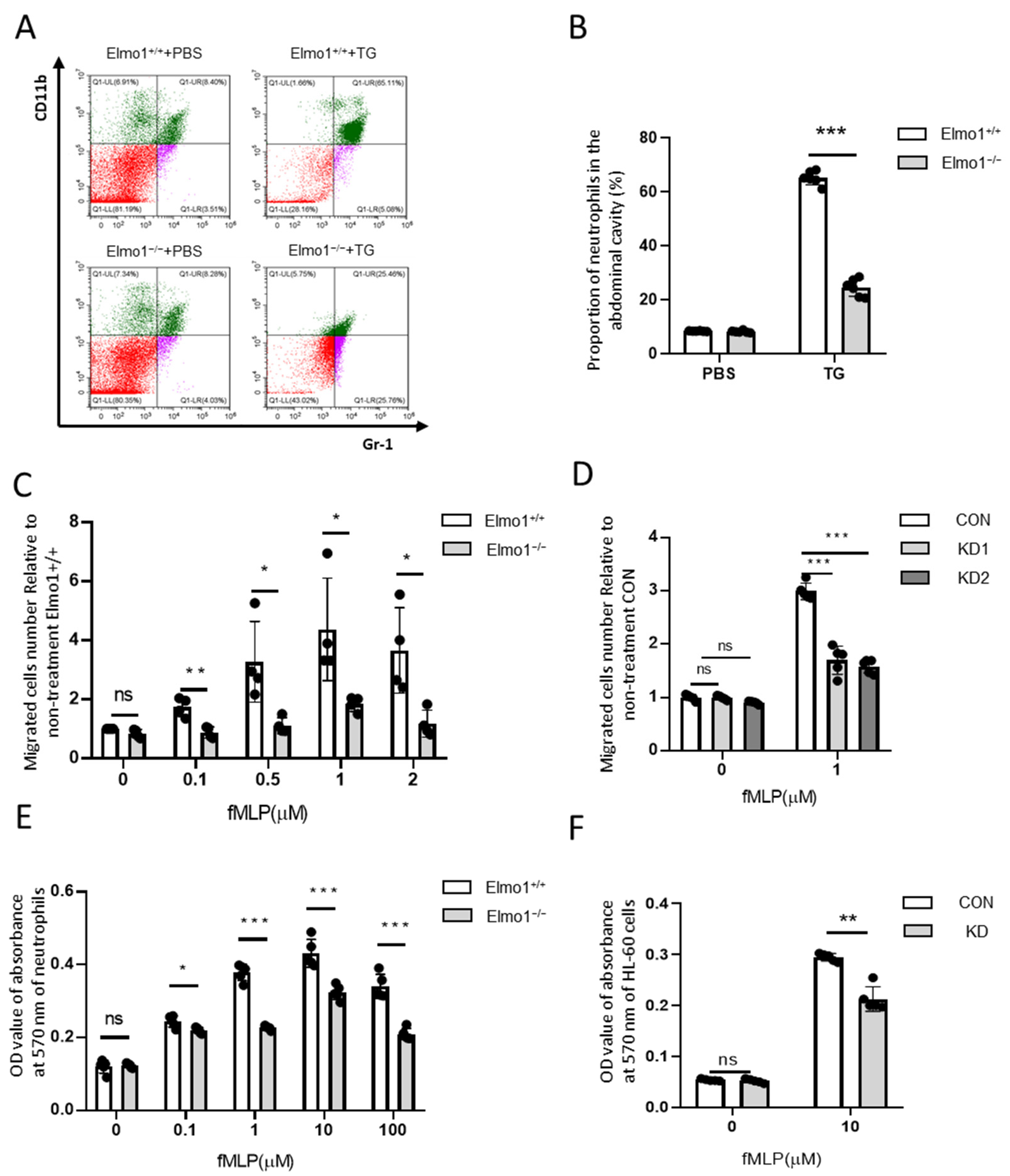
2.2. Elmo1 Deficiency Reduces the Chemotaxis and Adhesion of Neutrophils
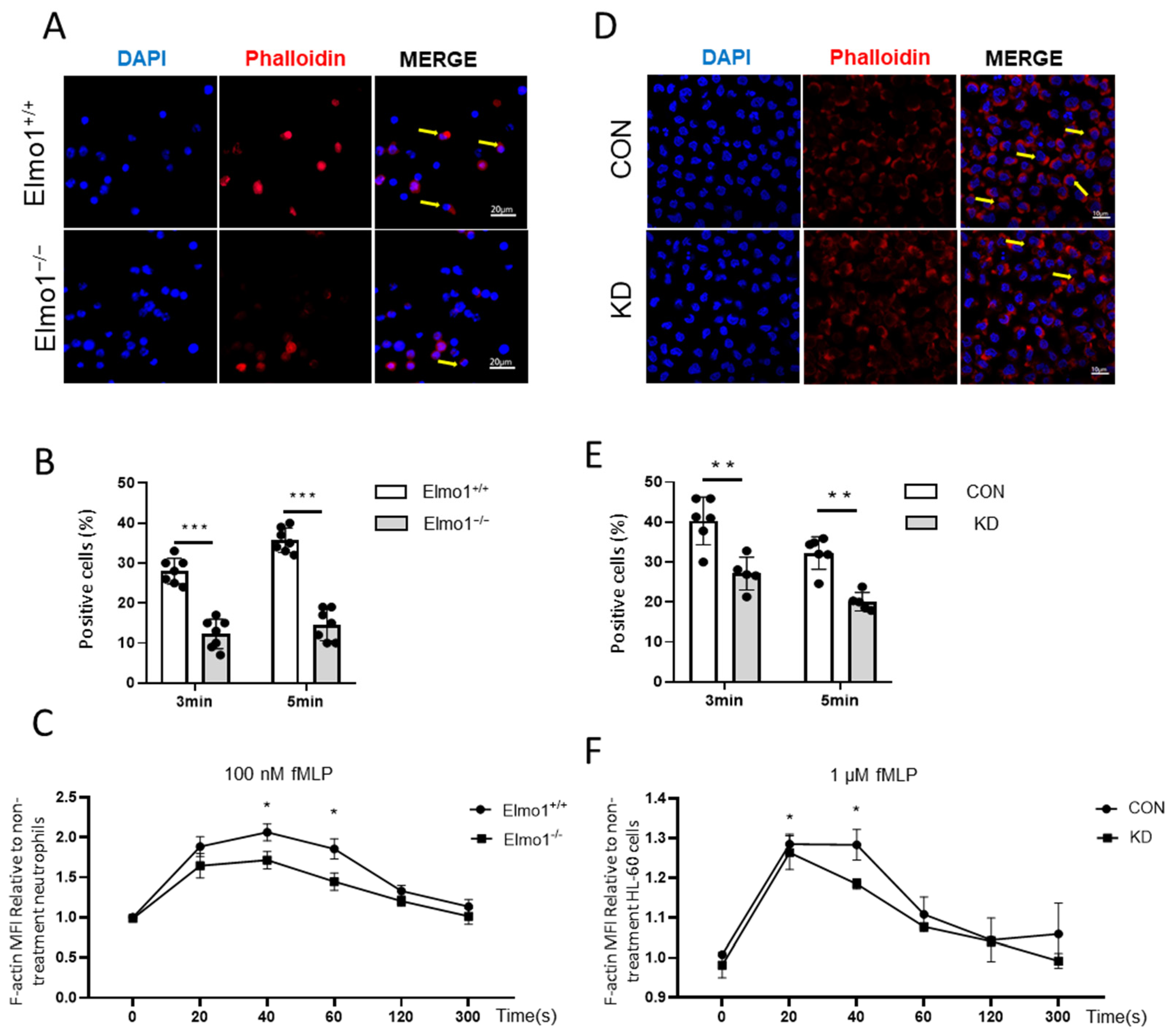
2.3. Deficiency of Elmo1 Down-Regulates fMLP-Induced Actin Polymerization
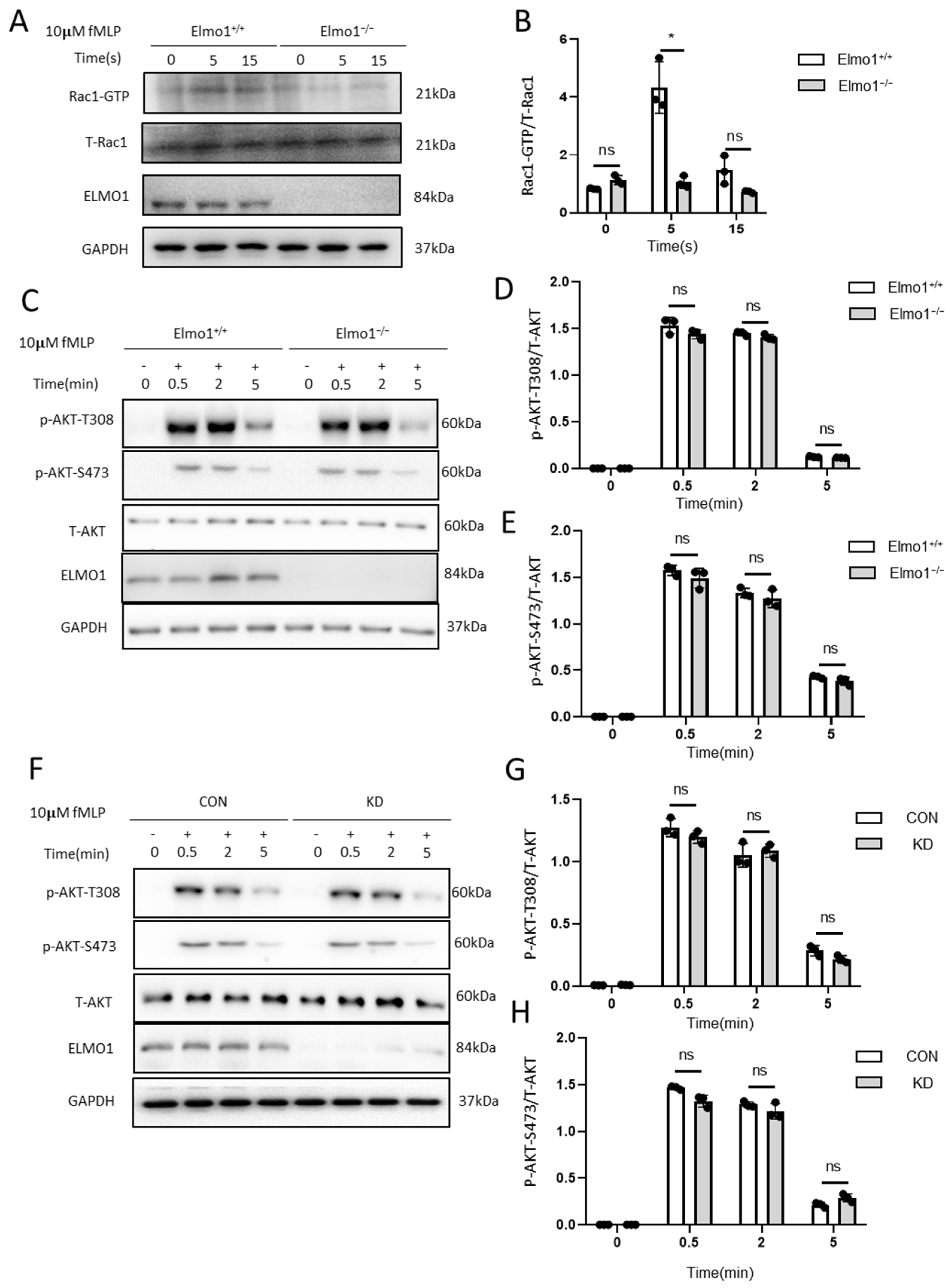
2.4. ELMO1 Regulates fMLP-Triggered Rac Activation Independent of PI3K and mTORC2 Signaling Pathways
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Primary Neutrophil Isolation and Identification
4.3. Cell Culture, Infection, and Differentiation
4.4. H&E, Immunofluorescence, and Phalloidin Staining Assay
4.5. Generation of the Peritonitis Mouse Model
4.6. Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. Adhesion Assay
4.8. Transwell Migration Assay
4.9. Rac Activity Assay
4.10. Western Blot Analysis
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Niccoli, G.; Montone, R.A.; Sabato, V.; Crea, F. Role of Allergic Inflammatory Cells in Coronary Artery Disease. Circulation 2018, 138, 1736–1748. [Google Scholar] [CrossRef] [PubMed]
- Konig, M.F.; Abusleme, L.; Reinholdt, J.; Palmer, R.J.; Teles, R.P.; Sampson, K.; Rosen, A.; Nigrovic, P.A.; Sokolove, J.; Giles, J.T.; et al. Aggregatibacter actinomycetemcomitans-induced hypercitrullination links periodontal infection to autoimmunity in rheumatoid arthritis. Sci. Transl. Med. 2016, 8, 369ra176. [Google Scholar] [CrossRef] [PubMed]
- Venet, F.; Monneret, G. Advances in the understanding and treatment of sepsis-induced immunosuppression. Nat. Rev. Nephrol. 2018, 14, 121–137. [Google Scholar] [CrossRef] [PubMed]
- Margraf, A.; Lowell, C.A.; Zarbock, A. Neutrophils in acute inflammation: Current concepts and translational implications. Blood 2022, 139, 2130–2144. [Google Scholar] [CrossRef]
- Petri, B.; Sanz, M.J. Neutrophil chemotaxis. Cell Tissue Res. 2018, 371, 425–436. [Google Scholar] [CrossRef]
- Michael, M.; Vermeren, S. A neutrophil-centric view of chemotaxis. Essays Biochem. 2019, 63, 607–618. [Google Scholar] [CrossRef]
- Chen, K.; Bao, Z.; Tang, P.; Gong, W.; Yoshimura, T.; Wang, J.M. Chemokines in homeostasis and diseases. Cell Mol. Immunol 2018, 15, 324–334. [Google Scholar] [CrossRef]
- Mafi, A.; Kim, S.K.; Goddard, W.A., 3rd. The mechanism for ligand activation of the GPCR-G protein complex. Proc. Natl. Acad. Sci. USA 2022, 119, e2110085119. [Google Scholar] [CrossRef]
- Surve, C.R.; To, J.Y.; Malik, S.; Kim, M.; Smrcka, A.V. Dynamic regulation of neutrophil polarity and migration by the heterotrimeric G protein subunits Galphai-GTP and Gbetagamma. Sci. Signal. 2016, 9, ra22. [Google Scholar] [CrossRef]
- Khater, M.; Wei, Z.; Xu, X.; Huang, W.; Lokeshwar, B.L.; Lambert, N.A.; Wu, G. G protein betagamma translocation to the Golgi apparatus activates MAPK via p110gamma-p101 heterodimers. J. Biol. Chem 2021, 296, 100325. [Google Scholar] [CrossRef]
- Liu, L.; Das, S.; Losert, W.; Parent, C.A. mTORC2 regulates neutrophil chemotaxis in a cAMP- and RhoA-dependent fashion. Dev. Cell 2010, 19, 845–857. [Google Scholar] [CrossRef]
- Takai, Y.; Sasaki, T.; Matozaki, T. Small GTP-binding proteins. Physiol. Rev. 2001, 81, 153–208. [Google Scholar] [CrossRef]
- Filippi, M.D.; Szczur, K.; Harris, C.E.; Berclaz, P.Y. Rho GTPase Rac1 is critical for neutrophil migration into the lung. Blood 2007, 109, 1257–1264. [Google Scholar] [CrossRef]
- Roberts, A.W.; Kim, C.; Zhen, L.; Lowe, J.B.; Kapur, R.; Petryniak, B.; Spaetti, A.; Pollock, J.D.; Borneo, J.B.; Bradford, G.B.; et al. Deficiency of the hematopoietic cell-specific Rho family GTPase Rac2 is characterized by abnormalities in neutrophil function and host defense. Immunity 1999, 10, 183–196. [Google Scholar] [CrossRef]
- Tuosto, L.; Capuano, C.; Muscolini, M.; Santoni, A.; Galandrini, R. The multifaceted role of PIP2 in leukocyte biology. Cell Mol. Life Sci. 2015, 72, 4461–4474. [Google Scholar] [CrossRef]
- Pare, G.; Vitry, J.; Merchant, M.L.; Vaillancourt, M.; Murru, A.; Shen, Y.; Elowe, S.; Lahoud, M.H.; Naccache, P.H.; McLeish, K.R.; et al. The Inhibitory Receptor CLEC12A Regulates PI3K-Akt Signaling to Inhibit Neutrophil Activation and Cytokine Release. Front. Immunol. 2021, 12, 650808. [Google Scholar] [CrossRef]
- Heit, B.; Liu, L.; Colarusso, P.; Puri, K.D.; Kubes, P. PI3K accelerates, but is not required for, neutrophil chemotaxis to fMLP. J. Cell Sci. 2008, 121, 205–214. [Google Scholar] [CrossRef]
- Ballesteros-Alvarez, J.; Andersen, J.K. mTORC2: The other mTOR in autophagy regulation. Aging Cell 2021, 20, e13431. [Google Scholar] [CrossRef]
- Nadolni, W.; Immler, R.; Hoelting, K.; Fraticelli, M.; Ripphahn, M.; Rothmiller, S.; Matsushita, M.; Boekhoff, I.; Gudermann, T.; Sperandio, M.; et al. TRPM7 Kinase Is Essential for Neutrophil Recruitment and Function via Regulation of Akt/mTOR Signaling. Front. Immunol. 2020, 11, 606893. [Google Scholar] [CrossRef]
- Gambardella, L.; Vermeren, S. Molecular players in neutrophil chemotaxis—Focus on PI3K and small GTPases. J. Leukoc. Biol. 2013, 94, 603–612. [Google Scholar] [CrossRef]
- Liang, X.; Hou, Y.; Han, L.; Yu, S.; Zhang, Y.; Cao, X.; Yan, J. ELMO1 Regulates RANKL-Stimulated Differentiation and Bone Resorption of Osteoclasts. Front. Cell Dev. Biol. 2021, 9, 702916. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, K.S.; Lorenz, U. Engulfment of apoptotic cells: Signals for a good meal. Nat. Rev. Immunol. 2007, 7, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Franke, K.; Otto, W.; Johannes, S.; Baumgart, J.; Nitsch, R.; Schumacher, S. miR-124-regulated RhoG reduces neuronal process complexity via ELMO/Dock180/Rac1 and Cdc42 signalling. EMBO J. 2012, 31, 2908–2921. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Mihaylov, V.; Xu, X.; Brzostowski, J.A.; Li, H.; Liu, L.; Veenstra, T.D.; Parent, C.A.; Jin, T. A Gbetagamma effector, ElmoE, transduces GPCR signaling to the actin network during chemotaxis. Dev. Cell 2012, 22, 92–103. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, X.; Pan, M.; Jin, T. ELMO1 Directly Interacts with Gbetagamma Subunit to Transduce GPCR Signaling to Rac1 Activation in Chemotaxis. J. Cancer 2016, 7, 973–983. [Google Scholar] [CrossRef]
- Li, H.; Yang, L.; Fu, H.; Yan, J.; Wang, Y.; Guo, H.; Hao, X.; Xu, X.; Jin, T.; Zhang, N. Association between Galphai2 and ELMO1/Dock180 connects chemokine signalling with Rac activation and metastasis. Nat. Commun. 2013, 4, 1706. [Google Scholar] [CrossRef]
- Arandjelovic, S.; Perry, J.S.A.; Lucas, C.D.; Penberthy, K.K.; Kim, T.H.; Zhou, M.; Rosen, D.A.; Chuang, T.Y.; Bettina, A.M.; Shankman, L.S.; et al. A noncanonical role for the engulfment gene ELMO1 in neutrophils that promotes inflammatory arthritis. Nat. Immunol. 2019, 20, 141–151. [Google Scholar] [CrossRef]
- Arandjelovic, S.; Perry, J.S.A.; Zhou, M.; Ceroi, A.; Smirnov, I.; Walk, S.F.; Shankman, L.S.; Cambre, I.; Onengut-Gumuscu, S.; Elewaut, D.; et al. ELMO1 signaling is a promoter of osteoclast function and bone loss. Nat. Commun. 2021, 12, 4974. [Google Scholar] [CrossRef]
- Jarzynka, M.J.; Hu, B.; Hui, K.M.; Bar-Joseph, I.; Gu, W.; Hirose, T.; Haney, L.B.; Ravichandran, K.S.; Nishikawa, R.; Cheng, S.Y. ELMO1 and Dock180, a bipartite Rac1 guanine nucleotide exchange factor, promote human glioma cell invasion. Cancer Res. 2007, 67, 7203–7211. [Google Scholar] [CrossRef]
- Xie, Z.; Yu, W.; Zheng, G.; Li, J.; Cen, S.; Ye, G.; Li, Z.; Liu, W.; Li, M.; Lin, J.; et al. TNF-alpha-mediated m(6)A modification of ELMO1 triggers directional migration of mesenchymal stem cell in ankylosing spondylitis. Nat. Commun. 2021, 12, 5373. [Google Scholar] [CrossRef]
- De Pascalis, C.; Etienne-Manneville, S. Single and collective cell migration: The mechanics of adhesions. Mol. Biol. Cell 2017, 28, 1833–1846. [Google Scholar] [CrossRef]
- Szczur, K.; Xu, H.; Atkinson, S.; Zheng, Y.; Filippi, M.D. Rho GTPase CDC42 regulates directionality and random movement via distinct MAPK pathways in neutrophils. Blood 2006, 108, 4205–4213. [Google Scholar] [CrossRef]
- Ambruso, D.R.; Knall, C.; Abell, A.N.; Panepinto, J.; Kurkchubasche, A.; Thurman, G.; Gonzalez-Aller, C.; Hiester, A.; deBoer, M.; Harbeck, R.J.; et al. Human neutrophil immunodeficiency syndrome is associated with an inhibitory Rac2 mutation. Proc. Natl. Acad. Sci. USA 2000, 97, 4654–4659. [Google Scholar] [CrossRef]
- Csepanyi-Komi, R.; Sirokmany, G.; Geiszt, M.; Ligeti, E. ARHGAP25, a novel Rac GTPase-activating protein, regulates phagocytosis in human neutrophilic granulocytes. Blood 2012, 119, 573–582. [Google Scholar] [CrossRef]
- Trepat, X.; Chen, Z.; Jacobson, K. Cell migration. Compr. Physiol. 2012, 2, 2369–2392. [Google Scholar] [CrossRef]
- Senoo, H.; Kamimura, Y.; Kimura, R.; Nakajima, A.; Sawai, S.; Sesaki, H.; Iijima, M. Phosphorylated Rho-GDP directly activates mTORC2 kinase towards AKT through dimerization with Ras-GTP to regulate cell migration. Nat. Cell Biol. 2019, 21, 867–878. [Google Scholar] [CrossRef]
- Bressenot, A.; Salleron, J.; Bastien, C.; Danese, S.; Boulagnon-Rombi, C.; Peyrin-Biroulet, L. Comparing histological activity indexes in UC. Gut 2015, 64, 1412–1418. [Google Scholar] [CrossRef]
- Resende, C.B.; Borges, I.; Goncalves, W.A.; Carneiro, R.; Rezende, B.M.; Pinho, V.; Nobre, V.; Teixeira, M.M. Neutrophil activity in sepsis: A systematic review. Braz. J. Med. Biol. Res. 2020, 53, e7851. [Google Scholar] [CrossRef]
- Fukui, S.; Fukui, S.; Van Bruggen, S.; Shi, L.; Sheehy, C.E.; Chu, L.; Wagner, D.D. NLRP3 inflammasome activation in neutrophils directs early inflammatory response in murine peritonitis. Sci. Rep. 2022, 12, 21313. [Google Scholar] [CrossRef]
- Lajqi, T.; Braun, M.; Kranig, S.A.; Frommhold, D.; Poschl, J.; Hudalla, H. LPS Induces Opposing Memory-like Inflammatory Responses in Mouse Bone Marrow Neutrophils. Int. J. Mol. Sci. 2021, 22, 9803. [Google Scholar] [CrossRef]
- Lajqi, T.; Frommhold, D.; Braun, M.; Alexander Kranig, S.; Poschl, J.; Hudalla, H. Gram-positive Staphylococcus aureus LTA promotes distinct memory-like effects in murine bone marrow neutrophils. Cell Immunol. 2022, 376, 104535. [Google Scholar] [CrossRef] [PubMed]
- Lajqi, T.; Kostlin-Gille, N.; Hillmer, S.; Braun, M.; Kranig, S.A.; Dietz, S.; Krause, C.; Ruhle, J.; Frommhold, D.; Poschl, J.; et al. Gut Microbiota-Derived Small Extracellular Vesicles Endorse Memory-like Inflammatory Responses in Murine Neutrophils. Biomedicines 2022, 10, 442. [Google Scholar] [CrossRef] [PubMed]
- Eckhardt, S.G.; Dai, A.; Davidson, K.K.; Forseth, B.J.; Wahl, G.M.; Von Hoff, D.D. Induction of differentiation in HL60 cells by the reduction of extrachromosomally amplified c-myc. Proc. Natl. Acad. Sci. USA 1994, 91, 6674–6678. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Zhang, S.; Xu, R.; Chu, H.; Biswas, S.; Yu, S.; Miao, D.; Li, W.; Li, S.; Brown, A.J.; et al. Elevated HB-EGF expression in neural stem cells causes middle age obesity by suppressing Hypocretin/Orexin expression. FASEB J. 2021, 35, e21345. [Google Scholar] [CrossRef]
- Smith, M.L.; Olson, T.S.; Ley, K. CXCR2- and E-selectin-induced neutrophil arrest during inflammation in vivo. J. Exp. Med. 2004, 200, 935–939. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.; Geng, X.; Liu, H.; Zhang, Y.; Cao, X.; Li, B.; Yan, J. ELMO1 Deficiency Reduces Neutrophil Chemotaxis in Murine Peritonitis. Int. J. Mol. Sci. 2023, 24, 8103. https://doi.org/10.3390/ijms24098103
Yu S, Geng X, Liu H, Zhang Y, Cao X, Li B, Yan J. ELMO1 Deficiency Reduces Neutrophil Chemotaxis in Murine Peritonitis. International Journal of Molecular Sciences. 2023; 24(9):8103. https://doi.org/10.3390/ijms24098103
Chicago/Turabian StyleYu, Shuxiang, Xiaoke Geng, Huibing Liu, Yunyun Zhang, Xiumei Cao, Baojie Li, and Jianshe Yan. 2023. "ELMO1 Deficiency Reduces Neutrophil Chemotaxis in Murine Peritonitis" International Journal of Molecular Sciences 24, no. 9: 8103. https://doi.org/10.3390/ijms24098103
APA StyleYu, S., Geng, X., Liu, H., Zhang, Y., Cao, X., Li, B., & Yan, J. (2023). ELMO1 Deficiency Reduces Neutrophil Chemotaxis in Murine Peritonitis. International Journal of Molecular Sciences, 24(9), 8103. https://doi.org/10.3390/ijms24098103