
Lavandula austroapennina (Lamiaceae): Getting Insights into Bioactive Polyphenols of a Rare Italian Endemic Vascular Plant


 ,
,  ,
, 
 ,
,  and
and
Abstract
1. Introduction
2. Results and Discussion
2.1. Chemical Investigation of L. austroapennina Alcoholic Extracts
2.1.1. Phenylpropenoic Acid Derivatives
2.1.2. Phenylpropanoic Acid Derivatives
2.1.3. Flavonoids
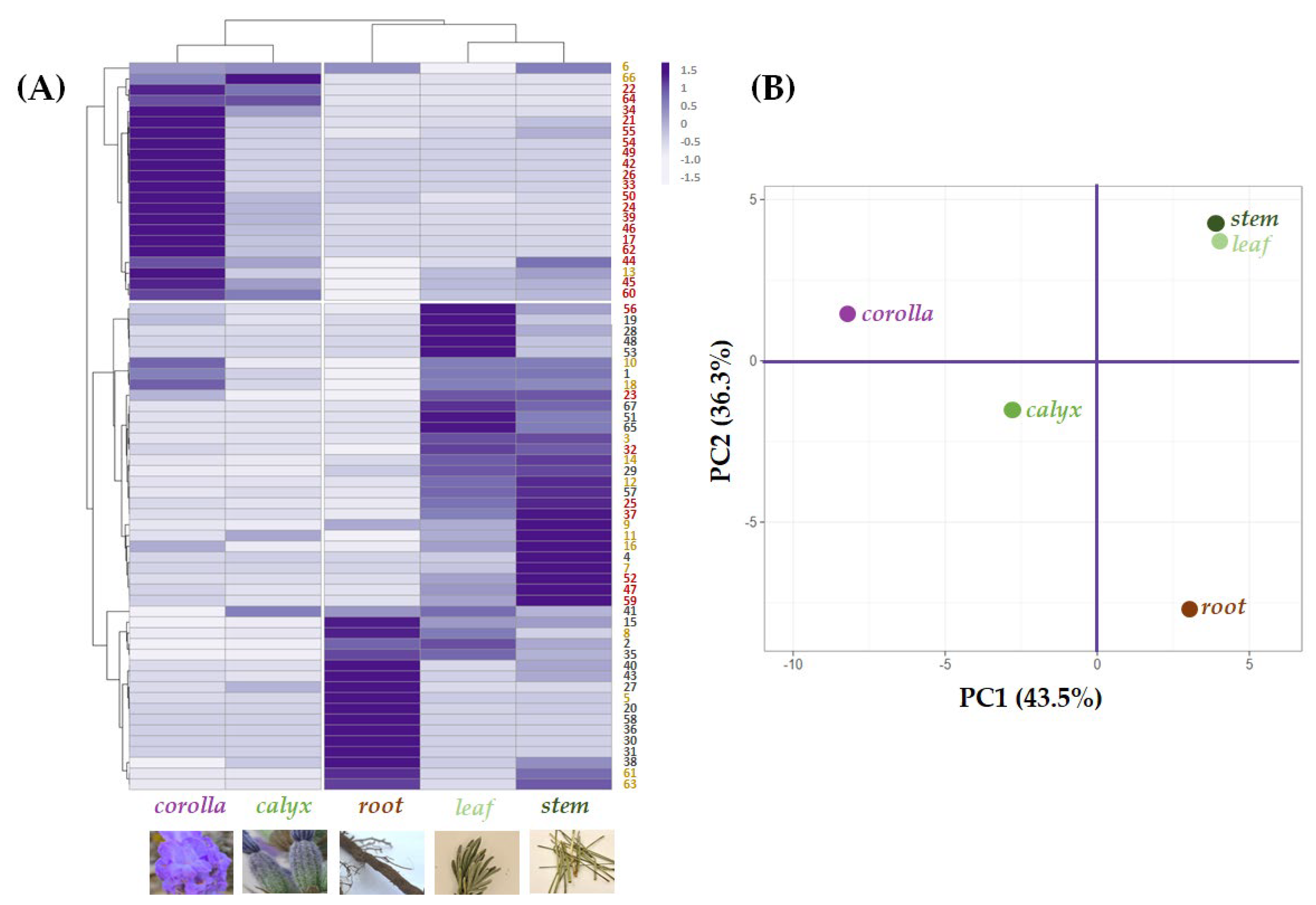
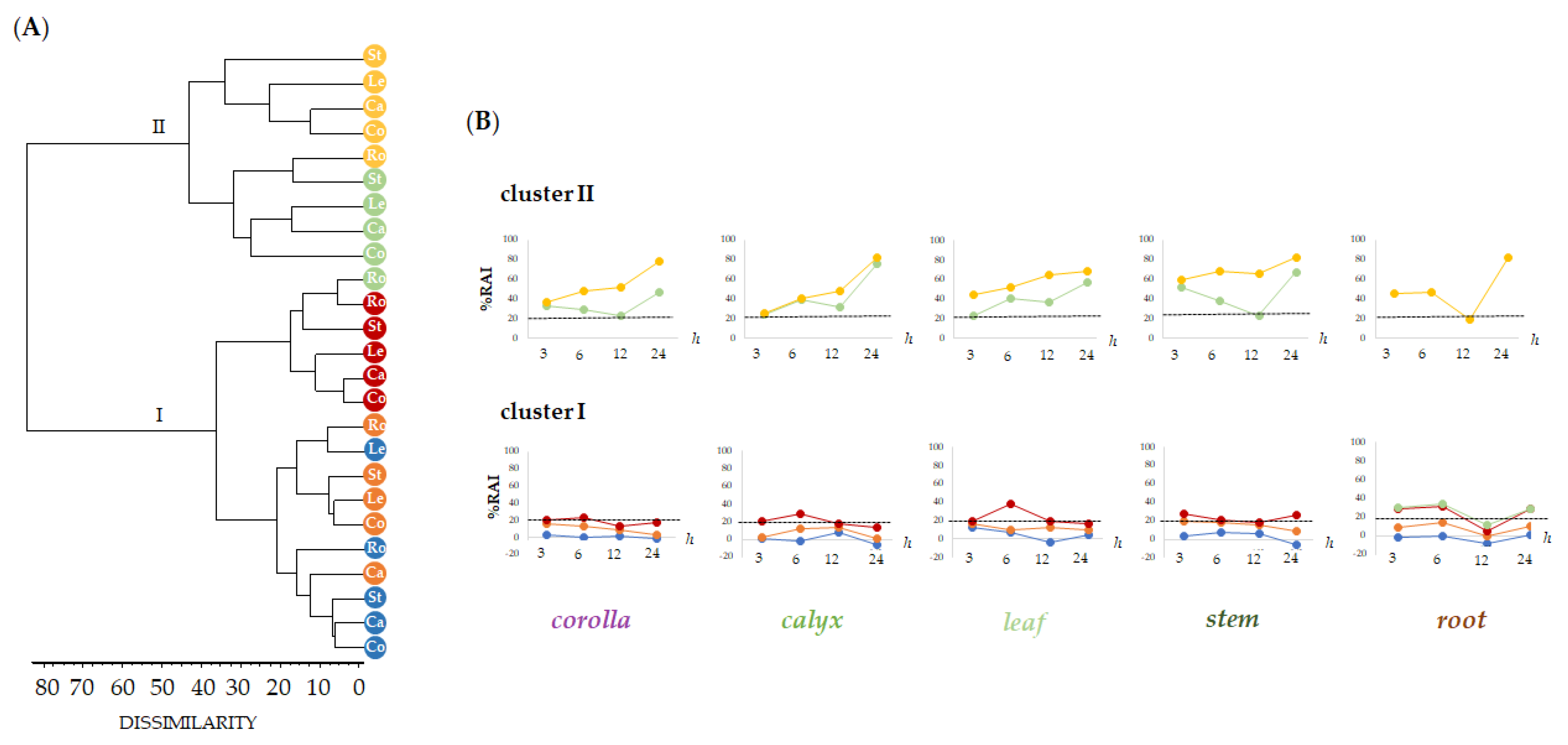
2.2. Relative Quantitation of Polyphenols in Lavandula austroapennina Organs
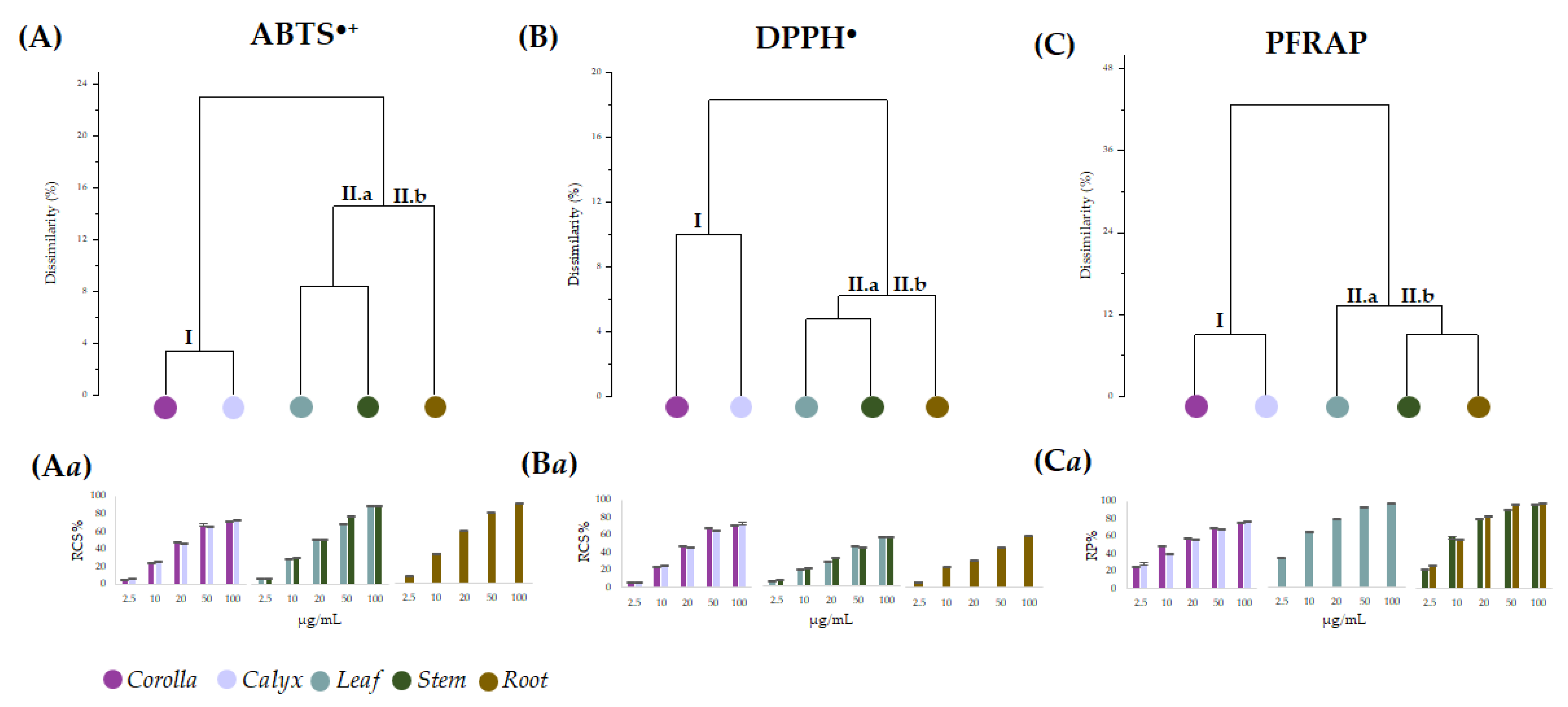
2.3. Antioxidant Activity of Lavandula austroapennina Alcoholic Extracts
2.4. Cytotoxic Screening of Alcoholic Extract from Lavandula austroapennina Organs
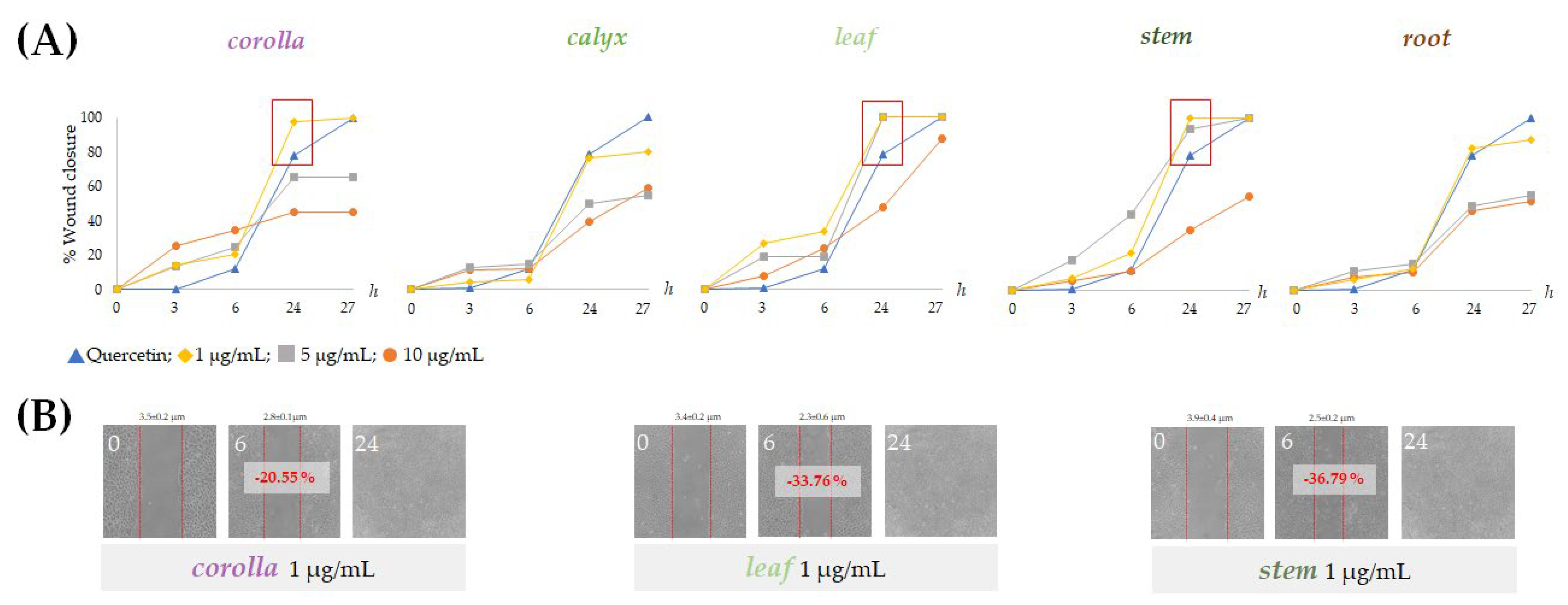
2.5. Alcoholic Extracts from Lavandula austroapennina Exert Wound-Healing Activity
3. Materials and Methods
3.1. Plant Collection and Extraction
3.2. UHPLC-ESI-QqTOF-MS and MS/MS Analyses
3.3. Antioxidant Assessment
3.4. Cell Culture and Cytotoxic Screening
3.5. Wound Scratch Assay
3.6. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aftab, T.; Rehman Hakeem, K. (Eds.) Medicinal and Aromatic Plants Healthcare and Industrial Applications; Springer Nature: Basel, Switzerland, 2021; p. 871. ISBN 978-3-030-58975-2. [Google Scholar] [CrossRef]
- Christaki, E.; Giannenas, I.; Bonos, E.; Florou-Paneri, P. Innovative Uses of Aromatic Plants as Natural Supplements in Nutrition. In Feed Additives: Aromatic Plants and Herbs in Animal Nutrition and Health; Academic Press: Cambridge, MA, USA, 2020; Chapter 2; pp. 19–34. [Google Scholar] [CrossRef]
- WHO (World Health Organization). Traditional Medicine Strategy 2014–2023; World Health Organization: Hong Kong, China, 2014; Available online: https://www.who.int/health-topics/traditional-complementary-and-integrative-medicine#tab=tab_1 (accessed on 20 January 2023).
- Allen, D.; Bilz, M.; Leaman, D.J.; Miller, R.M.; Timoshyna, A.; Window, J. European Red List of Medicinal Plants; Publications Office of the European Union: Luxemburg, 2014. [Google Scholar] [CrossRef]
- Chen, S.-L.; Yu, H.; Luo, H.-M.; Wu, Q.; Li, C.-F.; Steinmetz, A. Conservation and sustainable use of medicinal plants: Problems, progress, and prospects. Chin. Med. 2016, 11, 37. [Google Scholar] [CrossRef]
- Cocco, E.; Maccioni, D.; Sanjust, E.; Falconieri, D.; Farris, E.; Maxia, A. Ethnopharmacobotany and Diversity of Mediterranean Endemic Plants in Marmilla Subregion, Sardinia, Italy. Plants 2022, 11, 3165. [Google Scholar] [CrossRef]
- Bourgou, S.; Haj Jilani, I.B.; Karous, O.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; Greveniotis, V.; et al. Medicinal-Cosmetic Potential of the Local Endemic Plants of Crete (Greece), Northern Morocco and Tunisia: Priorities for Conservation and Sustainable Exploitation of Neglected and Underutilized Phytogenetic Resources. Biology 2021, 10, 1344. [Google Scholar] [CrossRef] [PubMed]
- Cheminal, A.; Kokkoris, I.P.; Strid, A.; Dimopoulos, P. Medicinal and Aromatic Lamiaceae Plants in Greece: Linking Diversity and Distribution Patterns with Ecosystem Services. Forests 2020, 11, 661. [Google Scholar] [CrossRef]
- Gonçalves, S.; Gomes, D.; Costa, P.; Romano, A. The phenolic content and antioxidant activity of infusions from Mediterranean medicinal plants. Ind. Crops Prod. 2013, 43, 465–471. [Google Scholar] [CrossRef]
- Lari, Z.N.; Hajimonfarednejad, M.; Riasatian, M.; Abolhassanzadeh, Z.; Iraji, A.; Vojoud, M.; Heydari, M.; Shams, M. Efficacy of inhaled Lavandula angustifolia Mill. Essential oil on sleep quality, quality of life and metabolic control in patients with diabetes mellitus type II and insomnia. J. Ethnopharmacol. 2020, 251, 112560. [Google Scholar] [CrossRef]
- Wells, R.; Truong, F.; Adal, A.M.; Sarker, L.S.; Mahmoud, S.S. Lavandula Essential Oils: A Current Review of Applications in Medicinal, Food, and Cosmetic Industries of Lavender. Nat. Prod. Commun. 2018, 13, 1403–1417. [Google Scholar] [CrossRef]
- Algieri, F.; Rodriguez-Nogales, A.; Vezza, T.; Garrido-Mesa, J.; Garrido-Mesa, N.; Utrilla, M.P.; González-Tejero, M.R.; Casares-Porcel, M.; Molero-Mesa, J.; Contreras, M.D.M.; et al. Anti-inflammatory activity of hydroalcoholic extracts of Lavandula dentata L. and Lavandula stoechas L. J. Ethnopharmacol. 2016, 190, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Dobros, N.; Zawada, K.; Paradowska, K. Phytochemical Profile and Antioxidant Activity of Lavandula angustifolia and Lavandula x intermedia Cultivars Extracted with Different Methods. Antioxidants 2022, 11, 711. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.; Gonçalves, S.; Valentão, P.; Andrade, P.B.; Almeida, C.; Nogueira, J.M.; Romano, A. Metabolic profile and biological activities of Lavandula pedunculata subsp. lusitanica (Chaytor) Franco: Studies on the essential oil and polar extracts. Food Chem. 2013, 141, 2501–2506. [Google Scholar] [CrossRef]
- Lesage-Meessen, L.; Bou, M.; Sigoillot, J.-C.; Faulds, C.B.; Lomascolo, A. Essential oils and distilled straws of lavender and lavandin: A review of current use and potential application in white biotechnology. Appl. Microbiol. Biotechnol. 2015, 99, 3375–3385. [Google Scholar] [CrossRef]
- Upson, T.; Andrews, S. The Genus Lavandula (A Botanical Magazine Monograph); Royal Botanic Gardens, Kew: Richmond, UK, 2004; ISBN 0-88192-642-6. [Google Scholar]
- Passalacqua, N.G.; Tundis, R.; Upson, T.M. A new species of Lavandula sect. Lavandula (Lamiaceae) and review of species boundaries in Lavandula angustifolia. Phytotaxa 2017, 292, 161–170. [Google Scholar] [CrossRef]
- Despinasse, Y.; Moja, S.; Soler, C.; Jullien, F.; Pasquier, B.; Bessière, J.-M.; Baudino, S.; Nicolè, F. Structure of the Chemical and Genetic Diversity of the True Lavender over Its Natural Range. Plants 2020, 9, 1640. [Google Scholar] [CrossRef]
- Orsenigo, S.; Montagnani, C.; Fenu, G.; Gargano, D.; Peruzzi, L.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Bovio, M.; et al. Red Listing plants under full national responsibility: Extinction risk and threats in the vascular flora endemic to Italy. Biol. Conserv. 2018, 224, 213–222. [Google Scholar] [CrossRef]
- Di Novella, R.; Di Novella, N.; De Martino, L.; Mancini, E.; De Feo, V. Traditional plant use in the National Park of Cilento and Vallo di Diano, Campania, Southern, Italy. J. Ethnopharmacol. 2013, 145, 328–342. [Google Scholar] [CrossRef]
- De Falco, E.; Rigano, D.; Fico, V.; Vitti, A.; Barile, G.; Pergola, M. Spontaneous Officinal Plants in the Cilento, Vallo di Diano and Alburni National Park: Tradition, Protection, Enhancement, and Recovery. Plants 2023, 12, 465. [Google Scholar] [CrossRef]
- Truzzi, E.; Chaouch, M.A.; Rossi, G.; Tagliazucchi, L.; Bertelli, D.; Benvenuti, S. Characterization and Valorization of the Agricultural Waste Obtained from Lavandula Steam Distillation for Its Reuse in the Food and Pharmaceutical Fields. Molecules 2022, 27, 1613. [Google Scholar] [CrossRef]
- Simeoni, M.C.; Pellegrini, M.; Sergi, M.; Pittia, P.; Ricci, A.; Compagnone, D. Analysis of Polyphenols in the Lamiaceae Family by Matrix Solid-Phase Dispersion Extraction Followed by Ultra-High-Performance Liquid Chromatography–Tandem Mass Spectrometry Determination. ACS Omega 2018, 3, 17610–17616. [Google Scholar] [CrossRef]
- Torras-Claveria, L.; Jauregui, O.; Bastida, J.; Codina, C.; Viladomat, F. Antioxidant Activity and Phenolic Composition of Lavandin (Lavandula x intermedia Emeric ex Loiseleur) Waste. J. Agric. Food Chem. 2007, 55, 8436–8443. [Google Scholar] [CrossRef]
- Lopes, C.L.; Pereira, E.; Soković, M.; Carvalho, A.M.; Barata, A.M.; Lopes, V.; Rocha, F.; Calhelha, R.C.; Barros, L.; Ferreira, I.C. Phenolic Composition and Bioactivity of Lavandula pedunculata (Mill.) Cav. Samples from Different Geographical Origin. Molecules 2018, 23, 1037. [Google Scholar] [CrossRef]
- Di Giacomo, S.; Percaccio, E.; Gullì, M.; Romano, A.; Vitalone, A.; Mazzanti, G.; Gaetani, S.; Di Sotto, A. Recent Advances in the Neuroprotective Properties of Ferulic Acid in Alzheimer’s Disease: A Narrative Review. Nutrients 2022, 14, 3709. [Google Scholar] [CrossRef]
- Kurkin, V.A.; Lamrini, M.; Klochkov, S.G. Lavandoside from Lavandula spica flowers. Chem. Nat. Compd. 2008, 44, 169–170. [Google Scholar] [CrossRef]
- Grayer, R.J.; Eckert, M.R.; Veitch, N.C.; Kite, G.C.; Marin, P.D.; Kokubun, T.; Simmonds, M.S.J.; Paton, A.J. The chemotaxonomic significance of two bioactive caffeic acid esters, nepetoidins A and B, in the Lamiaceae. Phytochemistry 2003, 64, 519–528. [Google Scholar] [CrossRef]
- Gericke, S.; Lübken, T.; Wolf, D.; Kaiser, M.; Hannig, C.; Speer, K. Identification of New Compounds from Sage Flowers (Salvia officinalis L.) as Markers for Quality Control and the Influence of the Manufacturing Technology on the Chemical Composition and Antibacterial Activity of Sage Flower Extracts. J. Agric. Food Chem. 2018, 66, 1843–1853. [Google Scholar] [CrossRef]
- Nigro, E.; Crescente, G.; Formato, M.; Pecoraro, M.T.; Mallardo, M.; Piccolella, S.; Daniele, A.; Pacifico, S. Hempseed Lignanamides Rich-Fraction: Chemical Investigation and Cytotoxicity towards U-87 Glioblastoma Cells. Molecules 2020, 25, 1049. [Google Scholar] [CrossRef]
- Li, X.; Yu, C.; Lu, Y.; Gu, Y.; Lu, J.; Xu, W.; Xuan, L.; Wang, Y. Pharmacokinetics, Tissue Distribution, Metabolism, and Excretion of Depside Salts from Salvia miltiorrhiza in Rats. Drug Metab. Dispos. 2007, 35, 234–239. [Google Scholar] [CrossRef]
- Du, G.; Sun, L.; Zhao, R.; Du, L.; Song, J.; Zhang, L.; He, G.; Zhang, Y.; Zhang, J. Polyphenols: Potential source of drugs for the treatment of ischaemic heart disease. Pharmacol. Ther. 2016, 162, 23–34. [Google Scholar] [CrossRef]
- Yan, X.; Zhou, T.; Tao, Y.; Wang, Q.; Liu, P.; Liu, C. Salvianolic acid B attenuates hepatocyte apoptosis by regulating mediators in death receptor and mitochondrial pathways. Exp. Biol. Med. 2010, 235, 623–632. [Google Scholar] [CrossRef]
- Tsai, M.-K.; Lin, Y.-L.; Huang, Y.-T. Effects of salvianolic acids on oxidative stress and hepatic fibrosis in rats. Toxicol. Appl. Pharmacol. 2010, 242, 155–164. [Google Scholar] [CrossRef]
- Hao, Y.; Xie, T.; Korotcov, A.; Zhou, Y.; Pang, X.; Shan, L.; Ji, H.; Sridhar, R.; Wang, P.; Califano, J.; et al. Salvianolic acid B inhibits growth of head and neck squamous cell carcinoma in vitro and in vivo via cyclooxygenase-2 and apoptotic pathways. Int. J. Cancer 2009, 124, 2200–2209. [Google Scholar] [CrossRef]
- Nicolai, M.; Pereira, P.; Vitor, R.F.; Reis, C.P.; Roberto, A.; Rijo, P. Antioxidant activity and rosmarinic acid content of ultrasound-assisted ethanolic extracts of medicinal plants. Measurement 2016, 89, 328–332. [Google Scholar] [CrossRef]
- Dahchour, A. Anxiolytic and antidepressive potentials of rosmarinic acid: A review with a focus on antioxidant and anti-inflammatory effects. Pharmacol. Res. 2022, 184, 106421. [Google Scholar] [CrossRef]
- Luo, C.; Zou, L.; Sun, H.; Peng, J.; Gao, C.; Bao, L.; Ji, R.; Jin, Y.; Sun, S. A Review of the Anti-Inflammatory Effects of Rosmarinic Acid on Inflammatory Diseases. Front. Pharmacol. 2020, 11, 153. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Imran, M.; Aslam Gondal, T.; Imran, A.; Shahbaz, M.; Muhammad Amir, R.; Wasim Sajid, M.; Batool Qaisrani, T.; Atif, M.; Hussain, G.; et al. Therapeutic Potential of Rosmarinic Acid: A Comprehensive Review. Appl. Sci. 2019, 9, 3139. [Google Scholar] [CrossRef]
- Noor, S.; Mohammad, T.; Rub, M.A.; Raza, A.; Azum, N.; Yadav, D.K.; Hassan, I.; Asiri, A.M. Biomedical features and therapeutic potential of rosmarinic acid. Arch. Pharmacal Res. 2022, 45, 205–228. [Google Scholar] [CrossRef]
- De Torre, M.P.; Cavero, R.Y.; Calvo, M.I. Anticholinesterase Activity of Selected Medicinal Plants from Navarra Region of Spain and a Detailed Phytochemical Investigation of Origanum vulgare L. ssp. vulgare. Molecules 2022, 27, 7100. [Google Scholar] [CrossRef]
- Grzegorczyk-Karolak, I.; Krzemińska, M.; Kiss, A.K.; Olszewska, M.A.; Owczarek, A. Phytochemical Profile and Antioxidant Activity of Aerial and Underground Parts of Salvia bulleyana Diels. Plants. Metabolites 2020, 10, 497. [Google Scholar] [CrossRef]
- Selenge, E.; Murata, T.; Tanaka, S.; Sasaki, K.; Batkhuu, J.; Yoshizaki, F. Monoterpene glycosides, phenylpropanoids, and acacetin glycosides from Dracocephalum foetidum. Phytochemistry 2014, 101, 91–100. [Google Scholar] [CrossRef]
- Zengin, G.; Llorent-Martínez, E.J.; Córdova, M.L.F.-D.; Bahadori, M.B.; Mocan, A.; Locatelli, M.; Aktumsek, A. Chemical composition and biological activities of extracts from three Salvia species: S. blepharochlaena, S. euphratica var. leiocalycina, and S. verticillata subsp. amasiaca. Ind. Crops Prod. 2018, 111, 11–21. [Google Scholar] [CrossRef]
- Del Valle, J.C.; Buide, M.L.; Casimiro-Soriguer, I.; Whittall, J.B.; Narbona, E. On flavonoid accumulation in different plant parts: Variation patterns among individuals and populations in the shore campion (Silene littorea). Front. Plant Sci. 2015, 6, 939. [Google Scholar] [CrossRef]
- Kachlicki, P.; Piasecka, A.; Stobiecki, M.; Marczak, Ł. Structural Characterization of Flavonoid Glycoconjugates and Their Derivatives with Mass Spectrometric Techniques. Molecules 2016, 21, 1494. [Google Scholar] [CrossRef]
- Adaszyńska-Skwirzyńska, M.; Dzięcioł, M. Comparison of phenolic acids and flavonoids contents in various cultivars and parts of common lavender (Lavandula angustifolia) derived from Poland. Nat. Prod. Res. 2017, 31, 2575–2580. [Google Scholar] [CrossRef] [PubMed]
- Panuccio, M.R.; Fazio, A.; Papalia, T.; Barreca, D. Antioxidant Properties and Flavonoid Profile in Leaves of Calabrian Lavandula multifida L., an Autochthon Plant of Mediterranean Southern Regions. Chem. Biodivers. 2016, 13, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Palacio, J.P.-D.; Díaz-Navarro, C.; Martín-Blázquez, A.; González-Olmedo, C.; Vergara-Segura, N.; Algieri, F.; Rodriguez-Nogales, A.; González-Tejero, M.R.; Casares-Porcel, M.; Molero-Mesa, J.; et al. Metabolomic analysis of Lavandula dentata L. and Lavandula stoechas L. extracts by LC-QTOF/MS experiments and multivariate analysis techniques as a chemotaxonomical tool. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2019, 154, 231–240. [Google Scholar] [CrossRef]
- Spiridon, I.; Colceru, S.; Anghel, N.; Teaca, C.A.; Bodirlau, R.; Armatu, A. Antioxidant capacity and total phenolic contents of oregano (Origanum vulgare), lavender (Lavandula angustifolia) and lemon balm (Melissa officinalis) from Romania. Nat. Prod. Res. 2011, 25, 1657–1661. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, J.; Gaspar, A.; Garrido, E.M.; Garrido, J.; Borges, F. Hydroxycinnamic Acid Antioxidants: An Electrochemical Overview. BioMed Res. Int. 2013, 2013, 251754. [Google Scholar] [CrossRef] [PubMed]
- Bors, W.; Christa, M.; Stettmaier, K.; Yinrong, L.; Yeap, F.L. Antioxidant Mechanisms of Polyphenolic Caffeic Acid Oligomers, Constituents of Salvia officinalis. Biol. Res. 2004, 37, 301–311. [Google Scholar] [CrossRef]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J. Dietary Flavonoid Aglycones and Their Glycosides: Which Show Better Biological Significance? Crit. Rev. Food Sci. Nutr. 2017, 57, 1874–1905. [Google Scholar] [CrossRef]
- Xiao, J.; Muzashvili, T.S.; Georgiev, M.I. Advances in the biotechnological glycosylation of valuable flavonoids. Biotechnol. Adv. 2014, 32, 1145–1156. [Google Scholar] [CrossRef]
- Hawrył, A.; Hawrył, M.; Waksmundzka-Hajnos, M. Liquid chromatography fingerprint analysis and antioxidant activity of selected lavender species with chemometric calculations. PLoS ONE 2019, 14, e0218974. [Google Scholar] [CrossRef]
- Gravina, C.; Fiorentino, M.; Formato, M.; Pecoraro, M.T.; Piccolella, S.; Stinca, A.; Pacifico, S.; Esposito, A. LC-HR/MS analysis of lipophilic extracts from Calendula arvensis (Vaill.) L. organs: An unexplored source in cosmeceuticals. Molecules 2022, 27, 8905. [Google Scholar] [CrossRef]
- Wang, S.-X.; Hu, L.-M.; Gao, X.-M.; Guo, H.; Fan, G.-W. Anti-inflammatory Activity of Salvianolic Acid B in Microglia Contributes to its Neuroprotective Effect. Neurochem. Res. 2010, 35, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Kotian, S.R.; Bhat, K.M.R.; Padma, D.; Pai, K.S.R. Influence of traditional medicines on the activity of keratinocytes in wound healing: An in-vitro study. Anat. Cell Biol. 2019, 52, 324–332. [Google Scholar] [CrossRef]
- Jiang, H.; Zhou, X.; Chen, L. Asiaticoside delays senescence and attenuate generation of ROS in UV-exposure cells through regulates TGF-β1/Smad pathway. Exp. Ther. Med. 2022, 24, 667. [Google Scholar] [CrossRef]
- Ruwizhi, N.; Aderibigbe, B.A. Cinnamic Acid Derivatives and Their Biological Efficacy. Int. J. Mol. Sci. 2020, 21, 5712. [Google Scholar] [CrossRef]
- Pontiki, E.; Hadjipavlou-Litina, D. Multi-Target Cinnamic Acids for Oxidative Stress and Inflammation: Design, Synthesis, Biological Evaluation and Modeling Studies. Molecules 2019, 24, 12. [Google Scholar] [CrossRef]
- Gunia-Krzyżak, A.; Słoczyńska, K.; Popiół, J.; Koczurkiewicz, P.; Marona, H.; Pękala, E. Cinnamic acid derivatives in cosmetics: Current use and future prospects. Int. J. Cosmet. Sci. 2018, 40, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.H.; Jo, Y.O.; Cho, C.-W.; Son, D.; Park, S.; Rho, J.; Choi, S.Y. Inhibitory Effects of Cinnamic Acid on Melanin Biosynthesis in Skin. Biol. Pharm. Bull. 2008, 31, 946–948. [Google Scholar] [CrossRef]
- Ghaisas, M.M.; Kshirsagar, S.B.; Sahane, R.S. Evaluation of wound healing activity of ferulic acid in diabetic rats. Int. Wound J. 2014, 11, 523–532. [Google Scholar] [CrossRef]
- Zhou, Z.; Shi, T.; Hou, J.; Li, M. Ferulic acid alleviates atopic dermatitis-like symptoms in mice via its potent anti-inflammatory effect. Immunopharmacol. Immunotoxicol. 2020, 42, 156–164. [Google Scholar] [CrossRef]
- Lim, K.-M.; Bae, S.; Koo, J.E.; Kim, E.-S.; Bae, O.-N.; Lee, J.Y. Suppression of skin inflammation in keratinocytes and acute/chronic disease models by caffeic acid phenethyl ester. Arch. Dermatol. Res. 2015, 307, 219–227. [Google Scholar] [CrossRef]
- Zhou, M.-W.; Jiang, R.-H.; Kim, K.-D.; Lee, J.-H.; Kim, C.-D.; Yin, W.-T.; Lee, J.-H. Rosmarinic acid inhibits poly(I:C)-induced inflammatory reaction of epidermal keratinocytes. Life Sci. 2016, 155, 189–194. [Google Scholar] [CrossRef]
- Koycheva, I.K.; Vasileva, L.V.; Amirova, K.M.; Marchev, A.S.; Balcheva-Sivenova, Z.P.; Georgiev, M.I. Biotechnologically Produced Lavandula angustifolia Mill. Extract Rich in Rosmarinic Acid Resolves Psoriasis-Related Inflammation Through Janus Kinase/Signal Transducer and Activator of Transcription Signaling. Front. Pharmacol. 2021, 12, 940. [Google Scholar] [CrossRef]
- Lin, J.; Lin, R.; Li, S.; Wu, H.; Ding, J.; Xiang, G.; Li, S.; Wang, Y.; Lin, D.; Gao, W.; et al. Salvianolic Acid B Promotes the Survival of Random-Pattern Skin Flaps in Rats by Inducing Autophagy. Front. Pharmacol. 2018, 9, 1178. [Google Scholar] [CrossRef]
- Guo, J.-W.; Cheng, Y.-P.; Liu, C.-Y.; Thong, H.-Y.; Huang, C.-J.; Lo, Y.; Wu, C.-Y.; Jee, S.-H. Salvianolic Acid B in Microemulsion Formulation Provided Sufficient Hydration for Dry Skin and Ameliorated the Severity of Imiquimod-induced Psoriasis-like Dermatitis in Mice. Pharmaceutics 2020, 12, 457. [Google Scholar] [CrossRef]
- Chen, Y.-S.; Lee, S.-M.; Lin, Y.-J.; Chiang, S.-H.; Lin, C.-C. Effects of Danshensu and Salvianolic Acid B from Salvia miltiorrhiza Bunge (Lamiaceae) on Cell Proliferation and Collagen and Melanin Production. Molecules 2014, 19, 2029–2041. [Google Scholar] [CrossRef]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef]
- Domaszewska-Szostek, A.; Puzianowska-Kuźnicka, M.; Kuryłowicz, A. Flavonoids in Skin Senescence Prevention and Treatment. Int. J. Mol. Sci. 2021, 22, 6814. [Google Scholar] [CrossRef]
- Leyva-López, N.; Gutierrez-Grijalva, E.P.; Ambriz-Perez, D.L.; Heredia, J.B. Flavonoids as Cytokine Modulators: A Possible Therapy for Inflammation-Related Diseases. Int. J. Mol. Sci. 2016, 17, 921. [Google Scholar] [CrossRef]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia; Edagricole: Milano, Italy, 2018; Volume 3. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia; Edagricole: Milano, Italy, 2019; Volume 4. [Google Scholar]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff [Continuously Updated]. Available online: http://sweetgum.nybg.org/ih/ (accessed on 28 February 2023).
- Formato, M.; Piccolella, S.; Zidorn, C.; Pacifico, S. UHPLC-HRMS Analysis of Fagus sylvatica (Fagaceae) Leaves: A Renewable Source of Antioxidant Polyphenols. Antioxidants 2021, 10, 1140. [Google Scholar] [CrossRef]
- Gebäck, T.; Schulz, M.M.P.; Koumoutsakos, P.; Detmar, M. TScratch: A novel and simple software tool for automated analysis of monolayer wound healing assays. Biotechniques 2009, 46, 265–274. [Google Scholar] [CrossRef]
- Grada, A.; Otero-Viñas, M.; Prieto-Castrillo, F.P.; Obagi, Z.; Falanga, V. Research Techniques Made Simple: Analysis of Collective Cell Migration Using the Wound Healing Assay. J. Investig. Dermatol. 2017, 137, e11–e16. [Google Scholar] [CrossRef]
- Podani, J. SYN-TAX 2000: Computer Programs for Data Analysis in Ecology and Systematics; User’s Manual; Scientia: Budapest, Hungary, 2001; p. 452. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Rt | Tentative Assignment | Formula | [M-H]− Found (m/z) | [M-H]− calcd. (m/z) | Error (ppm) | RDB | MS/MS Fragment Ions (m/z) |
|---|---|---|---|---|---|---|---|---|
| 3 | 2.083 | Caftaric acid | C13H12O9 | 311.0407 623.0903 a | 311.0409 | −0.5 | 8 | 179.0348; 149.0094; 135.0451 |
| 5 | 3.471 | Esculin | C15H16O9 | 339.0716 | 339.0722 | −1.6 | 8 | 177.0188; 133.0302 |
| 6 | 3.886 | p-Coumaric acid hexoside (isomer 1) | C15H18O8 | 325.0929 | 325.0929 | 0.0 | 7 | 163.0405; 119.0507 |
| 7 | 4.350 | Fertaric acid | C14H14O9 | 325.0565 | 325.0565 | 0.0 | 8 | 193.0511; 178.0267; 149.0610; 134.0378; 119.0504 |
| 8 | 4.360 | Caffeic acid hexoside (isomer 1) | C15H18O9 | 341.0872 | 341.0878 | −1.8 | 7 | 179.0356; 161.0246; 135.0457; 134.0371 |
| 9 | 5.525 | p-Coumaric acid dihexoside | C21H18O13 | 487.1467 | 487.1457 | 2.0 | 8 | 487.14621; 163.0394; 119.0501; 113.0242 |
| 10 | 6.164 | Ferulic acid hexoside (isomer 1) | C16H20O9 | 355.1032 | 355.1035 | 0.4 | 7 | 193.0508; 149.0613; 134.0377; 133.0294 |
| 11 | 6.535 | Caffeoylmalic acid (isomer 1) | C13H12O8 | 295.0462 | 295.0459 | 0.9 | 8 | 179.0348; 135.0454; 134.0373; 133.0143; 115.0042; 107.0510 |
| 12 | 6.791 | Caffeoylmalic acid (isomer 2) | C13H12O8 | 295.0463 | 295.0459 | 1.2 | 8 | 179.0349; 135.0453; 134.0370; 133.0145; 115.0043 |
| 13 | 7.659 | p-Coumaric acid hexoside (isomer 2) | C15H18O8 | 325.0931 | 325.0929 | 0.6 | 7 | 163.0400; 119.0503 |
| 14 | 7.907 | Ferulic acid dihexoside | C22H30O14 | 517.1572 | 517.1563 | 1.8 | 8 | 517.1600; 193.0510; 149.0615; 134.0372 |
| 16 | 9.347 | Caffeic acid hexoside (isomer 2) | C15H18O9 | 341.0883 | 341.0878 | 1.4 | 7 | 179.0345; 135.0450; 134.0358 |
| 18 | 10.021 | Ferulic acid hexoside (isomer 2) | C16H20O9 | 355.1038 | 355.1035 | 1.0 | 7 | 193.0501; 149.0606; 134.0373; 133.0297 |
| 61 | 19.294 | [(Z)-2-(3,5-dihydroxyphenyl)ethenyl] (E)-3-(3,4-dihydroxyphenyl)prop-2-enoate (Nepetoidin A) | C17H14O6 | 313.0720 | 313.0718 | 0.8 | 11 | 161.0246; 151.0401; 133.0296; 123.0438; 105.0345 |
| 63 | 19.770 | [(Z)-2-(3,4-dihydroxyphenyl)ethenyl] (E)-3-(3,4-dihydroxyphenyl)prop-2-enoate (Nepetoidin B) | C17H14O6 | 313.0712 | 313.0718 | −1.8 | 11 | 161.0243; 151.0400; 150.0318; 133.0292; 132.0213; 123.0450 |
| 66 | 20.861 | Tri-p-coumaroyl spermidine | C34H37N3O6 | 582.2626 | 582.2610 | 2.8 | 18 | 582.2651; 462.2058; 436.2262; 342.1446; 316.1667; 145.0299; 119.0506 |
| Peak | Rt | Tentative Assignment | Formula | [M-H]− Found (m/z) | [M-H]− calcd. (m/z) | Error (ppm) | RDB | MS/MS Fragment Ions (m/z) |
|---|---|---|---|---|---|---|---|---|
| 1 | 1.023 | 8-Hydroxydihydrocaffeic acid (danshensu) | C9H10O5 | 197.0462 | 197.0455 | 3.3 | 5 | 179.0357; 135.0450; 134.0374; 123.0449; 122.0372 |
| 2 | 1.699 | 8-Hydroxydihydrocaffeic acid hexoside | C15H20O10 | 359.0985 | 359.0984 | 0.4 | 6 | 359.0986; 197.0456; 179.0344; 135.0450; 134.0378; 123.0452 |
| 4 | 2.22 | Dihydrocaffeic acid | C9H10O4 | 181.0513 | 181.0506 | 3.7 | 5 | 163.0401; 135.0452; 134.0376; 119.0501; 117.0353; 107.0499 |
| 15 | 7.991 | Dihydroferulic acid hexoside | C16H22O9 | 357.1192 | 357.1191 | 0.3 | 6 | 195.0661; 177.0553; 151.0766; 136.0530; 121.0294 |
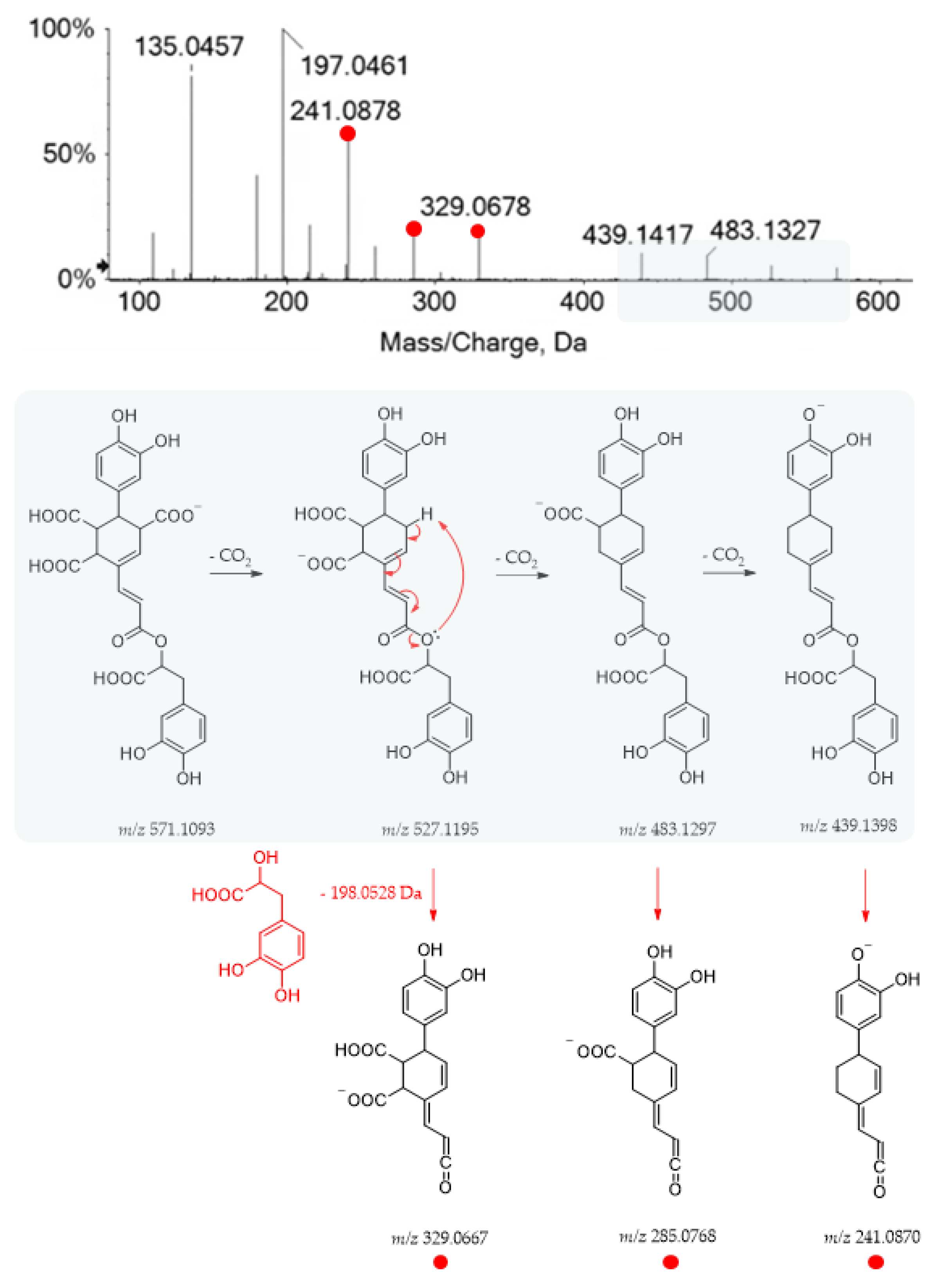
| 19 | 10.060 | Yunnaneic acid E | C27H24O14 | 571.1114 | 571.1093 | 3.6 | 16 | 527.1224; 483.1327; 439.1417; 329.0678; 285.0777; 241.0878; 215.1082; 197.0461; 179.0359; 135.0457; 109.0302 |
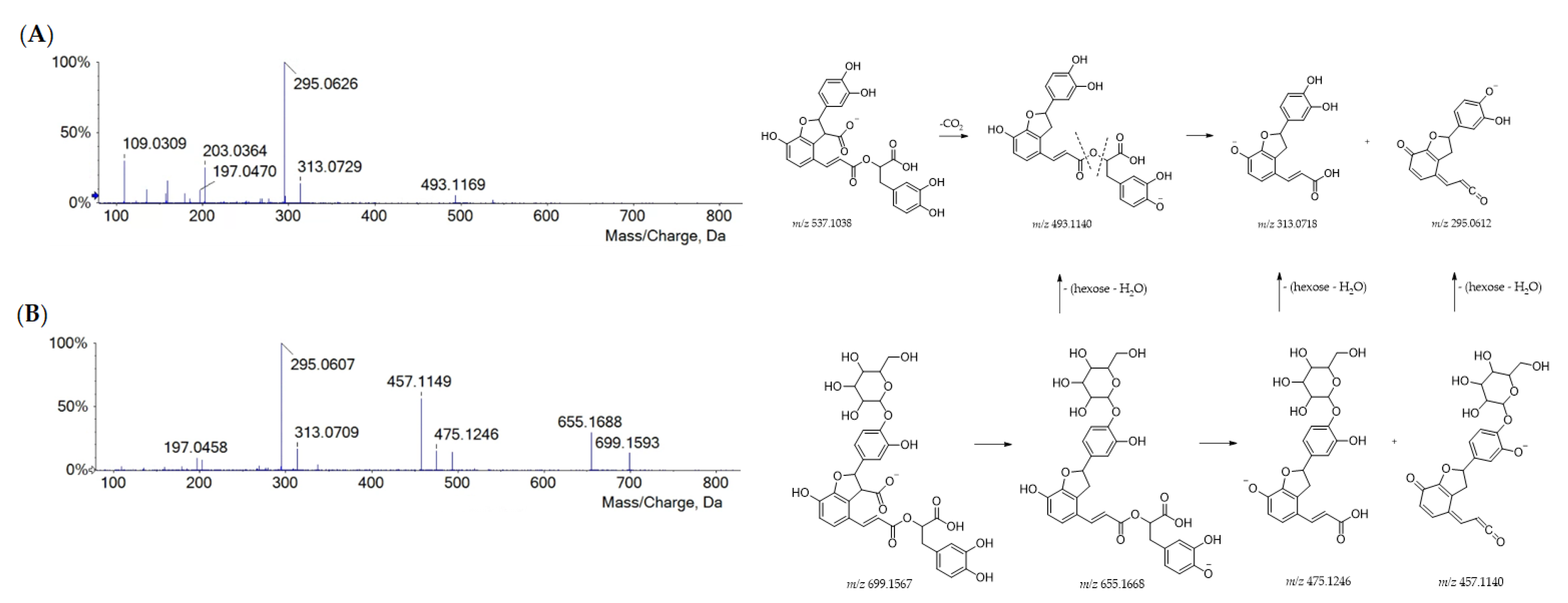
| 20 | 10.325 | Lithopermic acid A hexoside | C33H32O17 | 699.1563 | 699.1567 | −0.5 | 18 | 699.1593; 655.1688; 493.1149; 475.1246; 457.1149; 313.0709; 295.0607; 197.0458 |
| 27 | 11.488 | Lithospermic acid A | C27H22O12 | 537.1051 | 537.1039 | 2.3 | 17 | 537.1062; 493.1169; 313.0729; 295.0626; 203.0364; 197.0460; 159.0466; 109.0309 |
| 28 | 11.656 | Yunnaneic acid F (isomer 1) | C29H26O14 | 597.1257 | 597.1250 | 1.2 | 17 | 597.1256; 553.1345; 491.1338; 329.1041; 311.0900; 293.0781; 267.1002; 197.0447; 179.0330; 135.0448 |
| 29 | 11.915 | Yunnaneic acid F (isomer 2) | C29H26O14 | 597.1250 | 597.1250 | 0.0 | 17 | 597.1250; 535.1242; 481.1505; 417.0810; 399.0695; 355.0792; 311.0898; 293.0780; 267.1000; 241.1198; 197.0435; 179.0332; 135.0437 |
| 30 | 12.04 | 6-(3-(1-carboxy-2-(3,4-dihydroxyphenyl)ethoxy)-3-oxoprop-1-en-1-yl)-3-(3,4-dihydroxyphenyl)-8-hydroxy-7-oxobicyclo [2.2.2]oct-5-ene-2-carboxylic acid (isomer 1) | C27H24O12 | 539.1204 | 539.1195 | 1.7 | 16 | 539.1209; 359.0773; 297.0770; 279.0502; 271.0973; 197.0458; 179.0351; 161.0246; 135.0454; 133.0296 |
| 31 | 12.251 | 6-(3-(1-carboxy-2-(3,4-dihydroxyphenyl)ethoxy)-3-oxoprop-1-en-1-yl)-3-(3,4-dihydroxyphenyl)-8-hydroxy-7-oxobicyclo [2.2.2]oct-5-ene-2-carboxylic acid (isomer 2) | C27H24O12 | 539.1205 | 539.1195 | 1.9 | 16 | 539.1227; 359.0783; 341.0666; 315.0884; 297.0778; 271.0982; 253.0876; 135.0772; 197.0463; 179.0357; 161.0250; 135.0461 |
| 35 | 12.903 | Rosmarinic acid hexoside | C24H26O13 | 521.1322 | 521.1301 | 4.1 | 12 | 521.1333; 359.0782; 323.0775; 197.0460; 179.0353; 161.0248; 135.0454 |
| 36 | 12.979 | 6-(3-(1-carboxy-2-(3,4-dihydroxyphenyl)ethoxy)-3-oxoprop-1-en-1-yl)-3-(3,4-dihydroxyphenyl)-8-hydroxy-7-oxobicyclo [2.2.2]oct-5-ene-2-carboxylic acid (isomer 3) | C27H24O12 | 539.1216 | 539.1195 | 3.9 | 16 | 539.0781; 297.0778; 279.0665; 197.0460; 179.0355; 161.0250; 135.0455 |
| 38 | 13.293 | Rosmarinic acid isomer | C18H16O8 | 359.0775 | 359.0772 | 0.7 | 11 | 197.0456; 179.0350; 161.0248; 135.0450; 133.0296; 123.0449 |
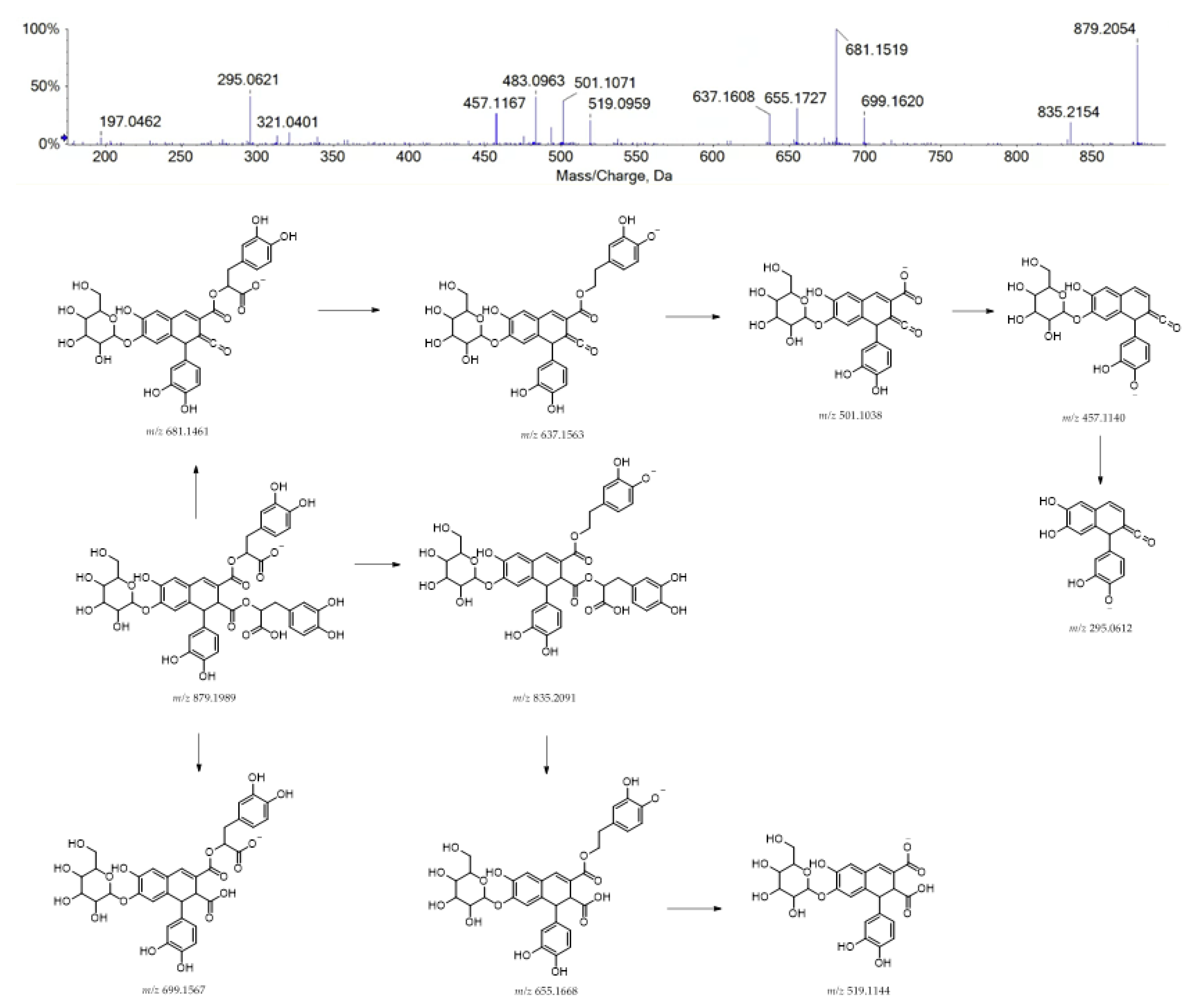
| 40 | 13.686 | Caffeic acid tetramer hexoside | C42H40O21 | 879.1991 | 879.1989 | 0.2 | 23 | 879.2054; 835.2154; 699.1620; 681.1519; 655.1727; 637.1608; 519.0959; 501.1071; 483.0963; 457.1167; 321.0401; 295.0621; 197.0462 |
| 41 | 13.963 | Rosmarinic acid | C18H16O8 | 359.0769 | 359.0772 | −1.0 | 11 | 197.0450; 179.0348; 161.0244; 135.0450; 133.0294; 123.0450 |
| 43 | 14.245 | Salvianolic acid B | C36H30O16 | 717.1487 | 717.1461 | 3.6 | 22 | 717.1494; 673.1598; 537.1062; 519.0953; 493.1158; 475.1054; 339.0511; 321.0400; 295.0609; 179.0351; 135.0453 |
| 48 | 14.447 | Yunnaneic acid E derivative 1 | C26H22O12 | 525.1054 | 525.1039 | 3.0 | 16 | 507.0953; 327.0507; 309.0395; 283.0612; 257.0803; 239.0705; 211.0758; 179.0346; 135.0452 |
| 51 | 14.867 | Yunnaneic acid F derivative | C29H24O13 | 579.1157 | 579.1144 | 2.2 | 18 | 579.1161; 491.1359; 399.0720; 293.0807; 355.0823; 311.0920; 293.0807; 267.1020; 135.0451 |
| 53 | 15.665 | Yunnaneic acid E derivative 2 | C26H20O12 | 523.0882 | 523.0882 | 0.0 | 17 | 523.0882; 505.0769; 479.0978; 325.0335; 299.0545; 255.0642; 237.0530; 211.0746; 179.0333; 135.0442 |
| 57 | 17.567 | Salvianolic acid C | C26H20O10 | 491.0997 | 491.0984 | 2.7 | 17 | 491.1012; 311.0563; 267.0661; 265.0502; 135.0454 |
| 58 | 18.267 | Sinapoyl-hexosyl-rosmarinic acid | C35H36O17 | 727.1906 | 727.1880 | 3.6 | 18.0 | 727.1934; 547.1479; 529.1383; 367.1040; 359.0785; 323.0772; 179.0355; 161.0250 |
| 65 | 20.612 | Dihydrosalvianolic acid A | C26H24O10 | 495.1314 | 495.1297 | 3.5 | 15 | 495.1329; 315.0886; 297.0775; 271.0975; 197.0451; 179.0349; 135.0455; 134.0374 |
| 67 | 21.596 | Salvianolic acid A | C26H22O10 | 493.1137 | 493.1140 | −0.7 | 16 | 493.1149; 313.0711; 269.0808; 135.0453 |
| Peak | Rt | Tentative Assignment | Formula | [M-H]− Found (m/z) | [M-H]− calcd. (m/z) | Error (ppm) | RDB | MS/MS Fragment Ions (m/z) |
|---|---|---|---|---|---|---|---|---|
| 17 | 9.450 | Dihexosyl hexuronidyl luteolin | C33H38O22 | 785.1768 | 785.1782 | −1.8 | 15 | 785.1768; 665.1341; 623.1254; 503.0831; 461.0719; 447.0917; 327.0521; 285.0405 |
| 21 | 10.507 | Apigenin di-hexuronide | C27H26O17 | 621.1097 | 621.1097 | 0.1 | 15 | 621.1160; 445.0802; 269.0463; 175.0243; 113.0244 |
| 22 | 10.668 | Hexosyl hexuronidyl apigenin | C27H28O18 | 607.1335 | 607.1305 | 5.0 | 14 | 607.1343; 431.1007; 269.0457; 113.0246 |
| 23 | 10.750 | Luteolin di-hexuronide | C27H26O16 | 637.1074 | 637.1046 | 4.3 | 15 | 637.1107; 461.0759; 285.0413 |
| 24 | 10.867 | Myricetin hexoside | C21H20O13 | 479.0848 | 479.0831 | 3.5 | 12 | 479.0841; 317.0305; 316.0214; 271.0238; 178.9977 |
| 25 | 10.988 | Luteolin hexuronyl-hexoside | C27H28O17 | 623.1272 | 623.1254 | 2.9 | 14 | 623.1269; 447.0938; 285.0395; 284.0316 |
| 26 | 11.295 | Dihexuronidyl tricetin | C27H26O19 | 653.1027 | 653.0996 | 4.8 | 15 | 653.1049; 477.0699; 343.0495; 301.0359 |
| 32 | 12.784 | Luteolin hexuronide (isomer 1) | C21H18O12 | 461.0736 | 461.0725 | 2.3 | 13 | 285.0406 |
| 33 | 12.839 | Dihexosyl dihexuronidyl tricetin | C39H46O29 | 977.2092 | 977.2052 | 4.1 | 17 | 977.2052; 815.1562; 801.1844; 639.1281; 477.0647; 301.0354 |
| 34 | 12.898 | Dihexosyl p-coumaroyl hexuronidyl tricetin | C42H44O25 | 947.2121 | 947.2099 | 2.3 | 21 | 947.2099; 785.1555; 771.1813; 609.1244; 463.0855; 301.0354 |
| 37 | 12.991 | Luteolin hexoside | C21H20O11 | 447.0931 | 447.0933 | −0.4 | 12 | 447.0940; 285.0401; 284.0324 |
| 39 | 13.482 | Malonylhexosyl hexuronidyl apigenin | C30H30O19 | 693.1338 | 693.1309 | 4.3 | 16 | 649.1464; 607.1354; 473.1118; 431.1012; 269.0450; 113.0240 |
| 42 | 13.966 | Kaempferol hexoside | C21H20O11 | 447.0954 | 447.0933 | 4.7 | 12 | 447.0951; 327.0487; 285.0405; 284.0337; 255.0302; 227.0357; 151.0024 |
| 44 | 14.321 | Apigenin hexuronide | C21H18O11 | 445.0794 | 445.0776 | −1.7 | 13 | 269.0455; 113.0245 |
| 45 | 14.376 | Apigenin hexoside | C21H20O10 | 431.0984 | 431.0984 | 0.1 | 12 | 269.0454; 113.0249 |
| 46 | 14.42 | Dihexosyl p-coumaroyl hexuronidyl luteolin | C42H44O24 | 931.2188 | 931.2150 | 4.1 | 21 | 931.2150; 769.1641; 593.1320; 447.0951; 285.0416 |
| 47 | 14.446 | Luteolin hexuronide (isomer 2) | C21H18O12 | 461.0730 | 461.0725 | 1.0 | 13 | 285.0400 |
| 49 | 14.616 | Trihexosyl p-coumaroyl luteolin | C39H50O25 | 917.2375 | 917.2357 | 1.9 | 20 | 917.2357; 755.1865; 609.1611; 593.1303; 489.1149; 447.0961; 325.0911; 285.0405 |
| 50 | 14.745 | Tricetin | C15H10O7 | 301.0352 | 301.0354 | −0.6 | 11 | 301.0352; 255.0312; 239.0331; 215.0394; 191.0345; 149.0246 |
| 52 | 15.394 | Luteolin malonyl-hexoside | C24H22O14 | 533.0947 | 533.0937 | 1.9 | 14 | 489.1065; 285.0402; 284.0324 |
| 54 | 15.924 | Dihexosyl p-coumaroyl tricetin | C36H36O19 | 771.1814 | 771.1778 | 4.7 | 19 | 771.1778; 609.1245; 463.0889; 301.0354; 300.0283 |
| 55 | 16.848 | Apigenin acetyl-hexoside (isomer 1) | C23H22O11 | 473.1100 | 473.1089 | 2.3 | 13 | 473.1115; 413.0882; 269.0444; 268.0375 |
| 56 | 17.018 | Luteolin | C15H10O5 | 285.0404 | 285.0405 | −0.2 | 11 | 285.0408; 175.0393; 151.0022; 133.0288 |
| 59 | 18.777 | Apigenin acetyl-hexoside (isomer 2) | C23H22O11 | 473.1098 | 473.1089 | 1.8 | 13 | 473.1106; 269.0441; 268.0369; 239.0312 |
| 60 | 19.033 | Apigenin | C15H10O5 | 269.0458 | 269.0455 | 0.9 | 11 | 269.0454; 151.0028; 117.0341 |
| 62 | 19.468 | Tricetin p-coumaroyl hexoside | C30H26O14 | 609.1263 | 609.1250 | 2.2 | 18 | 609.1263; 301.0354 |
| 64 | 20.339 | Apigenin p-coumaroyl hexoside | C30H26O12 | 577.1371 | 577.1352 | 3.3 | 18 | 577.1352; 269.0440 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gravina, C.; Formato, M.; Piccolella, S.; Fiorentino, M.; Stinca, A.; Pacifico, S.; Esposito, A. Lavandula austroapennina (Lamiaceae): Getting Insights into Bioactive Polyphenols of a Rare Italian Endemic Vascular Plant. Int. J. Mol. Sci. 2023, 24, 8038. https://doi.org/10.3390/ijms24098038
Gravina C, Formato M, Piccolella S, Fiorentino M, Stinca A, Pacifico S, Esposito A. Lavandula austroapennina (Lamiaceae): Getting Insights into Bioactive Polyphenols of a Rare Italian Endemic Vascular Plant. International Journal of Molecular Sciences. 2023; 24(9):8038. https://doi.org/10.3390/ijms24098038
Chicago/Turabian StyleGravina, Claudia, Marialuisa Formato, Simona Piccolella, Marika Fiorentino, Adriano Stinca, Severina Pacifico, and Assunta Esposito. 2023. "Lavandula austroapennina (Lamiaceae): Getting Insights into Bioactive Polyphenols of a Rare Italian Endemic Vascular Plant" International Journal of Molecular Sciences 24, no. 9: 8038. https://doi.org/10.3390/ijms24098038
APA StyleGravina, C., Formato, M., Piccolella, S., Fiorentino, M., Stinca, A., Pacifico, S., & Esposito, A. (2023). Lavandula austroapennina (Lamiaceae): Getting Insights into Bioactive Polyphenols of a Rare Italian Endemic Vascular Plant. International Journal of Molecular Sciences, 24(9), 8038. https://doi.org/10.3390/ijms24098038