
Efficient Isolation and Expansion of Limbal Melanocytes for Tissue Engineering



Abstract
1. Introduction
2. Experimental Design
2.1. Materials and Reagents
2.1.1. Glassware and Plasticware
- ➢
- Micropipette tips (0.5–20 µL, 100–200 µL, 1000 µL) (Greiner Bio-One, Frickenhausen, Germany);
- ➢
- A 12-well plate (Corning, Costar®, Kaiserslautern, Germany, catalog number: 3513);
- ➢
- A 60 mm cell culture dish (Corning, Falcon®, catalog number: 353004);
- ➢
- A 100 mm cell culture dish (Corning, Falcon®, catalog number: 353003);
- ➢
- Syringe filter 0.2 μm (VWR, Radnor, PA, USA, catalog number: 28145-501);
- ➢
- Disposable scalpel blades, No. 10 (pfm Medical ag, Feather®, catalog number: 201000010);
- ➢
- Serological pipettes (5 mL, 10 mL) (Corning, Stripette™);
- ➢
- Conical tubes (15 mL) (Greiner Bio-One, catalog number: 188271);
- ➢
- Conical tubes (50 mL) (Greiner Bio-One, catalog number: 227261);
- ➢
- T75 flasks (Corning, catalog number: CLS430641);
- ➢
- Reversible cell strainers (Stem Cell Technologies, Köln, Germany, catalog number: 27215);
- ➢
- Cell filter 20 µm (Cell Trics™, Sysmex Partec GmbH, Norderstedt, Germany, catalog number: 04-004-2325);
- ➢
- FACS tubes (5 mL polystyrene round-bottom tube, Falcon, catalog number: 352058).
2.1.2. Antibodies
- ➢
- PE mouse anti-human CD117 (BD Biosciences, Heidelberg, Germany, catalog number: 561682);
- ➢
- APC mouse anti-human CD90 (BD Biosciences, catalog number: 55986);
- ➢
- Mouse IgG2a, k isotype-APC (eBioscience, Thermo Scientific, Dreieich, Germany, catalog number: 17-4724-81);
- ➢
- Mouse IgG2a, k isotype-PE (Biolegend, Amsterdam, The Netherlands, catalog number: 400212);
- ➢
- Mouse anti-human cytokeratin pan AE1/AE3 (DAKO, Hamburg, Germany, catalog number: M3515);
- ➢
- Rat anti-human vimentin (R&D Systems, Wiesbaden, Germany, catalog number: MAB2105);
- ➢
- Rabbit anti-human Melan-A (Abcam, Cambridge, UK, catalog number: ab210546);
- ➢
- Rabbit anti-human Sox10 (Abcam, catalog number: ab155279);
- ➢
- Mouse anti-human HMB-45 (Abcam, catalog number: ab787);
- ➢
- Mouse anti-human TRP1 (Abcam, catalog number: ab235447).
2.1.3. Culture Media, Reagents, and Buffers
- ➢
- For washing tissues and cells, Dulbecco’s Phosphate Buffered Saline (DPBS) (no calcium, no magnesium) (Thermo Fisher Scientific, Dreieich, Germany, Gibco™, catalog number: 14190094) was used;
- ➢
- Dulbecco’s Modified Eagle Medium (DMEM) high glucose (Thermo Fisher Scientific, Gibco™, catalog number: 11960044) and fetal bovine serum (FBS) (Thermo Scientific, Gibco™, catalog number: 10082147) were used to inhibit trypsin action;
- ➢
- To propagate LMs, we recommend using CnT-40 melanocyte proliferation medium (CELLnTEC, Bern, Switzerland);
- ➢
- For successful expansion of LMs, we used iMATRIX 511 (Nippi, Tokyo, Japan, catalog number: 892012) to coat the cultureware;
- ➢
- Penicillin–streptomycin 100× (Sigma-Aldrich, Taufkirchen, Germany, catalog number: P4333);
- ➢
- EDTA 0.5 M (Invitrogen, Karlsruhe, Germany, catalog number: AM9260G) was used in FACS buffer to reduce cell clump formation;
- ➢
- As a disinfectant, 70% isopropyl alcohol was used;
- ➢
- 4′,6-diamidino-2-phenylindole (DAPI) (Sigma-Aldrich, catalog number: MBD0015) was used during FACS to differentiate live and dead cells. You may experiment with any viability indicator that works well with the conjugated antibodies used in sorting.
2.1.4. Cell and Tissue Dissociation Reagents
- ➢
- Collagenase A (Sigma-Aldrich, Roche Diagnostics, Basel, Switzerland, catalog number: 10103578001) was used to digest the limbal segments;
- ➢
- Trypsin–EDTA 0.25% (Thermo Fisher Scientific, Gibco®, catalog number: 25200056) was used to dissociate limbal clusters into single cells as well as passaging of limbal melanocytes.
2.2. Equipment Required
- ➢
- Pipettes: micropipettes of all sizes (P20, P200, P1000; Eppendorf Research plus) and pipette aids (BRAND, accu-jet® pro) were required;
- ➢
- Centrifuge: the centrifuge must be capable of reaching speeds of at least 400× g with rotors to accommodate 15 mL and 50 mL centrifuge tubes and FACS tubes;
- ➢
- Cell counter: In this study, we used hemocytometer (MARIENFELD, Lauda-Königshofen, Germany, catalog number: 0640130). You may use any kind of cell counter to count the cell numbers;
- ➢
- Surgical instruments: to dissect the tissues, forceps (Dumont, Hausen ob Verena, Germany, 5-Dumoxel®-H) were required;
- ➢
- Aseptic environment: A biosafety cabinet (Thermo Scientific™, Type S2020 1.2) was required to create an aseptic environment for cell isolation and culturing. All the steps need to be performed under a biosafety cabinet;
- ➢
- CO2 incubator: for optimal cell culturing, CO2 incubator (Thermo Scientific™, Heracell™ 240i) was required to maintain 37 °C and 5% CO2;
- ➢
- Microscope: to visualize cell phases, contrast-inverted microscope with a camera (ZEISS, Oberkochen, Germany, Objectives 4×, 10×, 20×) was required;
- ➢
- Refrigerator and freezer: a refrigerator at 2–8 °C and freezer at −20 °C were required for storage of media and reagents;
- ➢
- Flow sorter: In this study, we used FACS Aria II sorter (BD Biosciences) to sort the cells. You may use any other cell sorter to sort the cells using this protocol;
- ➢
- Water bath: a water bath (GFL®, Karlsbad, Germany, catalog number: 1013) was required to warm the reagents.
2.3. Software Required
- ➢
- CapturePro 2.10.0.1 (JENOPTIC Optical systems GmbH; Villingen-Schwenningen, Germany);
- ➢
- FACSDiva v8.0.1 software (BD Pharmingen, BD Biosciences, Heidelberg, Germany);
- ➢
- FlowJo v10.2 software (Tree Star, Ashland, OR, USA).
2.4. Preparation of Solutions
3. Procedure
- A.
- Tissue
- 1.
- Organ-cultured corneoscleral tissues (age of 64.5 ± 12.2 years; culture duration 23.0 ± 3.5 days; postmortem time of 32.66 ± 15.8 h) were obtained from the cornea bank with appropriate research consent and ethical approval [11]. Tissues that are not suitable for transplantation due to low corneal endothelial cell density (<2200 cells/mm2) or the presence of neurological disorders or malignancies in the donors can be used. Donor cornea remnants after corneal endothelial transplant preparation are also a valuable source if appropriate research consent has been obtained.
- B.
- Dissection of limbus
- 2.
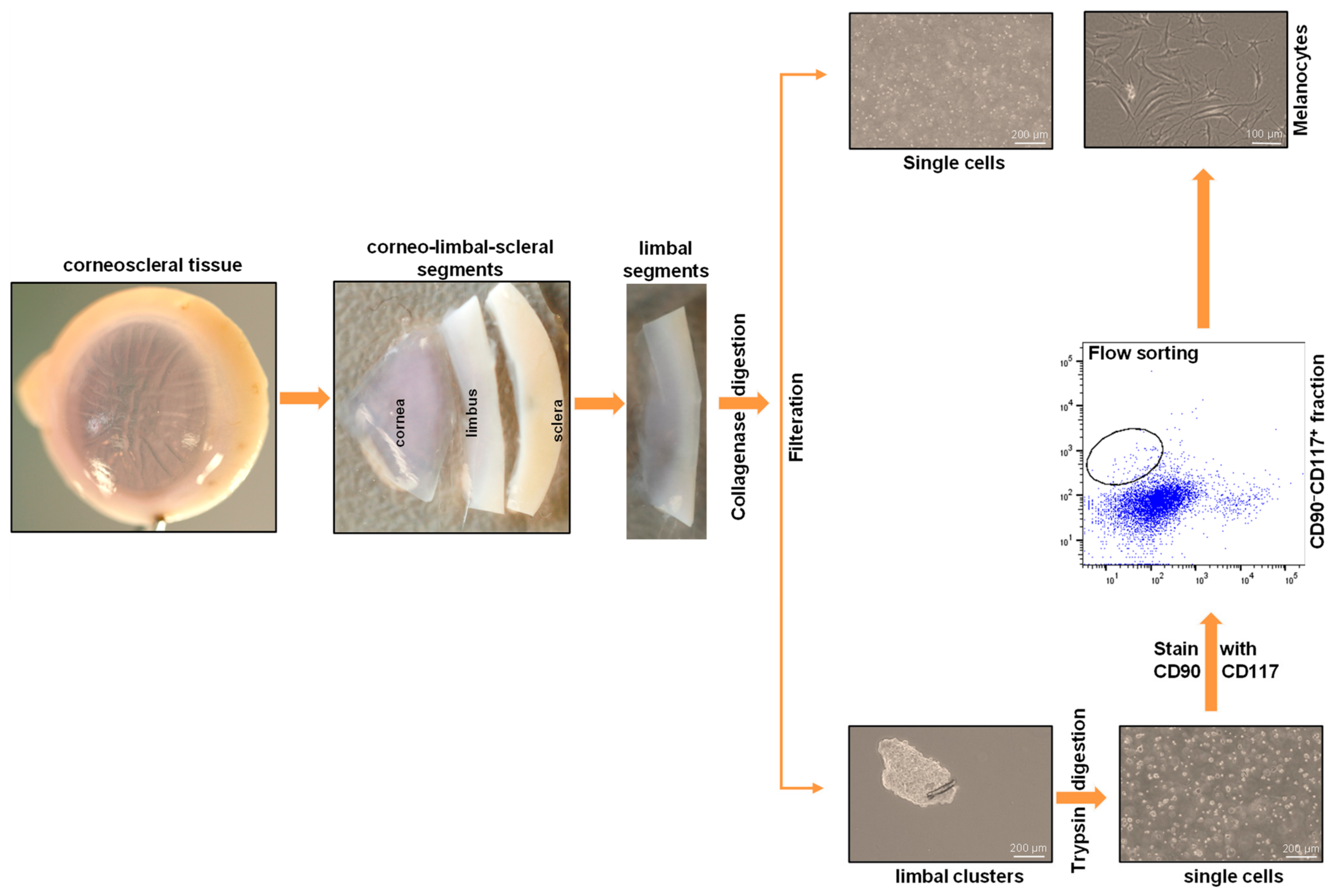
- Place the organ-cultured corneoscleral tissue in a 60 mm culture dish, and wash twice with DPBS. Using a scalpel blade and forceps, scrape the posterior side (endothelial side) of the tissue to remove unwanted tissue, such as trabecular meshwork and iris/ciliary tissue;
- 3.
- Place the tissue in a new 60 mm culture dish containing 1× DPBS, and cut the tissue into four equal quarters (Figure 2A) of corneoscleral segments (Figure 2B) using a scalpel blade and forceps;Note: to obtain adequate LMs for downstream applications, use 4–6 corneoscleral tissues.
- 4.
- Make incisions of corneoscleral segments at 1 mm before and beyond the anatomical limbus to obtain limbal segments (Figure 2C).
- C.
- Preparation of limbal cell suspension
- 5.
- Place the limbal segments in a 60 mm dish with 5 mL of collagenase A (2 mg/mL), and cut them into smaller pieces (each limbal segment into 2–3 pieces) with a scalpel blade. Incubate overnight at 37 °C with 5% CO2 to digest the stromal collagen and obtain limbal cell clusters;
- 6.
- After overnight incubation (~16–18 h), triturate the suspension with an up-and-down motion using a 1 mL pipette (P1000), and examine for the presence of cell clusters and single cells under the microscope (Figure 2D);Note: If incomplete digestion of limbal segments occurs after overnight incubation trituration, re-incubate for an additional 2 h in the same solution at 37 °C with 5% to ensure complete digestion. In contrast, prolonged digestion of tissue (more than 20 h) may have a negative impact on cell viability and quality.
- 7.
- To separate limbal cell clusters from single cells, filter the tissue digest using 20 µm cell filters that allow single cells to flow through while retaining the clusters;
- 8.
- To remove any remaining single cells, wash the filters twice with DPBS;
- 9.
- Place the strainer in the opposite direction on a 60 mm dish. Add 5 mL of 0.25% trypsin–EDTA to flush clusters (Figure 2E) into a Petri dish, and incubate at 37 °C (incubator) for 10–15 min to dissociate the clusters into single cells;Note: In place of 20 µm cell filters, 37 µm reversible cell strainers can be used. Do not allow the filter to dry as this might cause cell death. The filtered single-cell suspension can be discarded or used for other purposes, such as obtaining limbal fibroblasts.
- 10.
- After incubation, triturate the cell suspension with an up-and-down motion using a 1 mL pipette. Observe the cell suspension under the microscope to ensure the complete dissociation of clusters into single cells (Figure 2F). Add 5 mL of pre-warmed DMEM (37 °C in water bath) containing 10% FBS to inhibit trypsin digestion;Note: If clusters are not completely dissociated after 15 min of incubation and trituration, re-incubate for an additional 5 min in the same solution at 37 °C with 5% CO2 to achieve complete dissociation. Prolonged incubation of clusters in trypsin–EDTA might adversely affect cell viability and quality.
- 11.
- Filter the cell suspension using a 40 µm cell strainer into a 15 mL Falcon tube to remove any cell clumps. Wash the cell strainer with DPBS twice;
- 12.
- Centrifuge at 200× g for 5 min at room temperature;
- 13.
- After centrifugation, remove the supernatant carefully, and resuspend the cell pellet in 200 µL of ice-cold FACS buffer (see Section 2.4) by pipetting up and down using a P200 pipette.
- D.
- Fluorescence-activated cell sorting (FACS)
- 14.
- Transfer the cell suspension to FACS tubes (100 µL/tube). Add a mouse APC-conjugated anti-human CD90 antibody (5 µL/106 cells) and PE-conjugated anti-human CD117 antibody (5 µL/106 cells) to one tube and an IgG2a-Isotype APC and an IgG2a-Isotype PE to another tube at 4 °C;Note: The cell density should be <1 × 107 cells/mL. If the cell number is high, adjust the volumes and concentration of antibodies accordingly.
- 15.
- Incubate the cells on ice for 45 min, agitating every 15 min
- 16.
- After incubation, add 1 mL of FACS buffer to each FACS tube, and centrifuge for 5 min at 400× g at 4 °C. Repeat the washing process twice;
- 17.
- After washing, add 500 µL of FACS buffer containing DAPI (1:5000) to the cell pellet to exclude dead cells, and proceed to flow sorting using a FACS Aria II sorter [11];
- 18.
- Set up the FACS Aria II machine according to manufacturer’s instructions;
- 19.
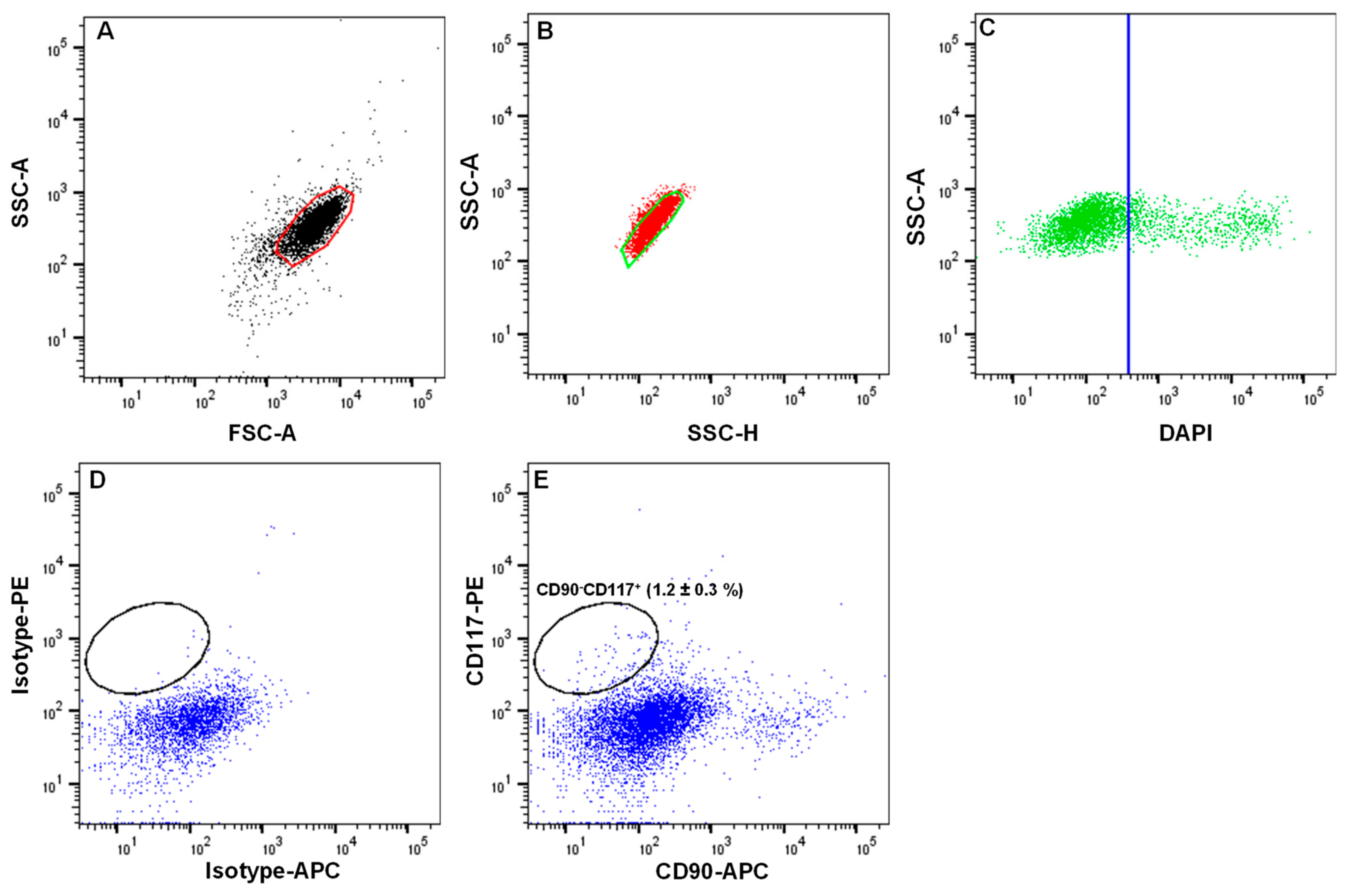
- Analyze the limbal cell suspensions. Set the gate on forward scatter (FSC-A) and side scatter (SSC-A) to select cells of interest based on size and granularity (Figure 3A). To remove doublets or clumps and enrich single cells, employ side scatter area vs. width (Figure 3B), followed by dead cell exclusion based on DAPI (Figure 3C);
- 20.
- Then, depending on the isotype controls, adjust the gates to select CD90− CD117+ (Figure 3D);Note: a positive region for each antibody staining needs to be defined, and this is defined as a region that contains almost no cells (e.g., <0.3%) when stained with the corresponding isotype control antibody.
- 21.
- Sort the CD90− CD117+ cells into 2 mL of FACS buffer in FACS tubes.Note: the CD90− CD117− populations mainly contain LEPCs and can be used to enrich LEPCs using cell type-specific media, whereas the CD90+ population containing LMSCs can be used for the expansion of LMSCs in cell type-specific media [11].
- E.
- Expansion of limbal melanocytes
- 22.
- Seed all the sorted CD90− CD117+ LMs into a well of a 12-well plate coated with i-Matrix-511 (see Section 2.4, laminin-coated plates);Note: the number of CD90− CD117+ cells per limbus varies from sample to sample.
- 23.
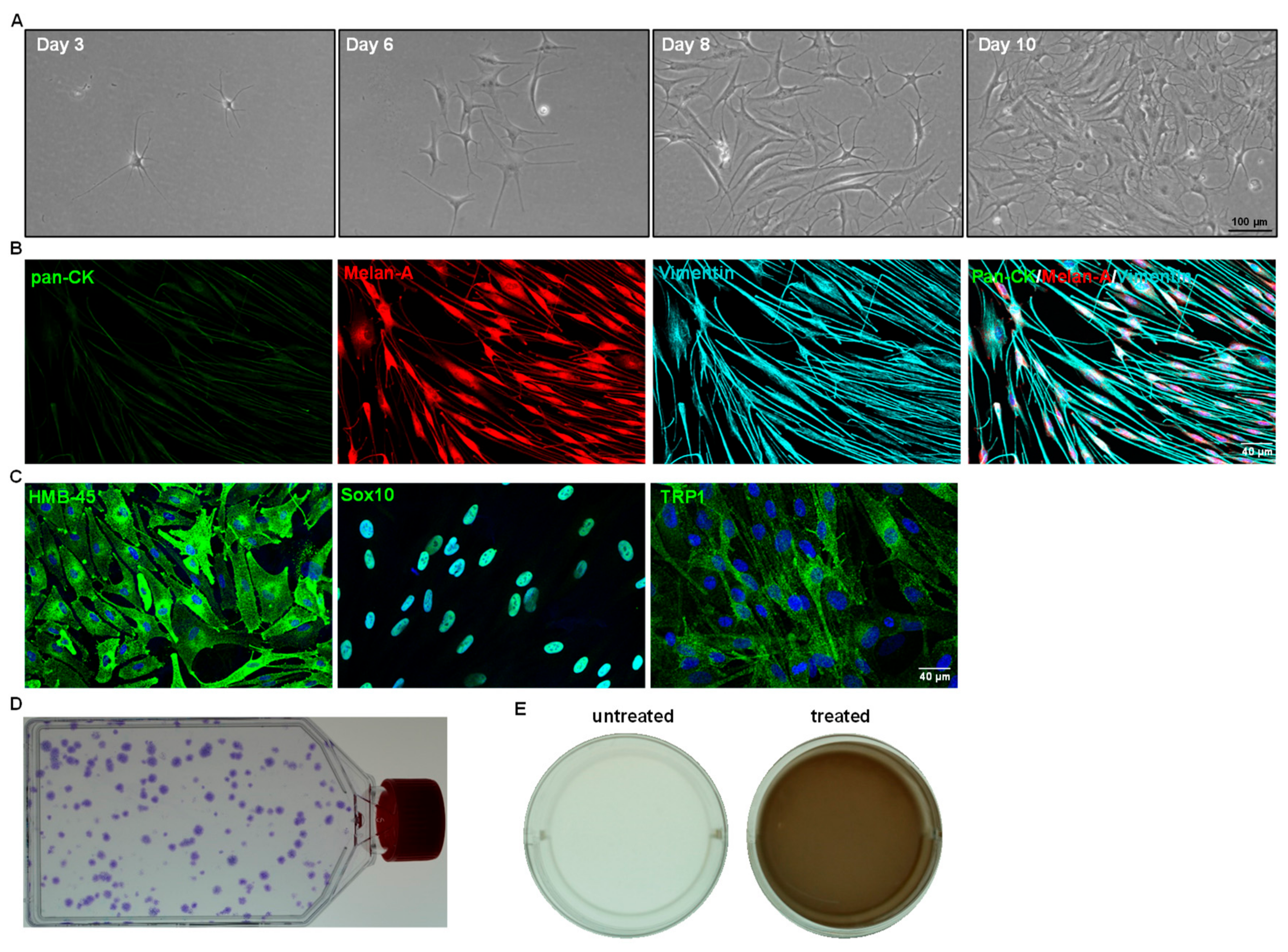
- Cultivate the LMs at 37 °C with 5% CO2 in CnT-40 medium (2 mL/well) to expand LMs. Change media every 2 days. Visualize the morphology of LMs using phase-contrast microscopy.
- F.
- Sub-cultivation of limbal melanocytes
- 24.
- Remove the media from the culture vessel at 70 to 80% confluency;
- 25.
- Wash the cells using DPBS, and add 1 mL (per well 12-well plate) of trypsin–EDTA (0.25%; pre-warmed at 37 °C in a water bath). Incubate at 37 °C with 5% CO2 for 5 min;
- 26.
- After incubation, add 1 mL of DMEM containing 10% FBS to inhibit trypsin action, and mix well;
- 27.
- Transfer the cell suspension to a 15 mL tube, and centrifuge at 200× g for 5 min. Remove the supernatant carefully, resuspend the cell pellet in CnT-40 medium and count the total cell number using a hemocytometer;Note: after 10 to 15 days of culture (roughly 70 to 80% confluency), the number of cells obtained from the well of a 12-well culture plate ranges from 40 to 60,000 cells.
- 28.
- Seed the cells in T25 flask (2 × 103 cells/cm2, i.e., 50,000 cells/T25 flask) coated with i-Matrix-511 in CnT-40 medium, and change the media every other day;Note: Over-confluence (more than 80%) and prolonged trypsin digestion (more than 5 min) adversely affect cell viability and the quality of cells during sub-culturing. Always passage cells at 70 to 80% confluence. Avoid prolonged incubations in trypsin. We recommend the seeding density of 2 × 103 cells/cm2 for expansion of LMs. If the obtained cell number is lower than 50,000 cells, we still recommend seeding all the cells in T25 flask.
- 29.
- After 10 to 14 days, repeat the steps from 24 to 27. Use the cells for the application of choice or sub-culturing. For subculturing, seed all the cells in T75 flask coated with i-Matrix-511 (see Section 2.4) in CnT-40 medium, and change the media every other day. Incubate at 37 °C with 5% CO2 for 5 min;
- 30.
- Passage the cells (2 × 103 cells/cm2) until the desired number of LMs is achieved for application of choice. Melanocytes take 10 to 14 days to reach confluence (with seeding density of 2 × 103 cells/cm2). The culturing and passaging of LMs can be carried out for 24 months without any significant changes in cell proliferation or behavior;
- 31.
4. Expected Results
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shortt, A.J.; Secker, G.A.; Munro, P.M.; Khaw, P.T.; Tuft, S.J.; Daniels, J.T. Characterization of the Limbal Epithelial Stem Cell Niche: Novel Imaging Techniques Permit In Vivo Observation and Targeted Biopsy of Limbal Epithelial Stem Cells. Stem Cells 2007, 25, 1402–1409. [Google Scholar] [CrossRef] [PubMed]
- Ordonez, P.; Di Girolamo, N. Limbal Epithelial Stem Cells: Role of the Niche Microenvironment. Stem Cells 2012, 30, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Polisetti, N.; Zenkel, M.; Menzel-Severing, J.; Kruse, F.E.; Schlötzer-Schrehardt, U. Cell Adhesion Molecules and Stem Cell-Niche-Interactions in the Limbal Stem Cell Niche. Stem Cells 2016, 34, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Mei, H.; Gonzalez, S.; Deng, S.X. Extracellular Matrix Is an Important Component of Limbal Stem Cell Niche. J. Funct. Biomater. 2012, 3, 879–894. [Google Scholar] [CrossRef]
- Higa, K.; Shimmura, S.; Miyashita, H.; Kato, N.; Ogawa, Y.; Kawakita, T.; Shimazaki, J.; Tsubota, K. N-Cadherin in the Maintenance of Human Corneal Limbal Epithelial Progenitor Cells In Vitro. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4640–4645. [Google Scholar] [CrossRef]
- Liu, L.; Nielsen, F.M.; Emmersen, J.; Bath, C.; Østergaard Hjortdal, J.; Riis, S.; Fink, T.; Pennisi, C.P.; Zachar, V. Pigmentation Is Associated with Stemness Hierarchy of Progenitor Cells within Cultured Limbal Epithelial Cells. Stem Cells 2018, 36, 1411–1420. [Google Scholar] [CrossRef]
- Dziasko, M.A.; Tuft, S.J.; Daniels, J.T. Limbal Melanocytes Support Limbal Epithelial Stem Cells in 2D and 3D Microenvironments. Exp. Eye Res. 2015, 138, 70–79. [Google Scholar] [CrossRef]
- Dziasko, M.A.; Armer, H.E.; Levis, H.J.; Shortt, A.J.; Tuft, S.; Daniels, J.T. Localisation of Epithelial Cells Capable of Holoclone Formation In Vitro and Direct Interaction with Stromal Cells in the Native Human Limbal Crypt. PLoS ONE 2014, 9, e94283. [Google Scholar] [CrossRef]
- Polisetti, N.; Gießl, A.; Zenkel, M.; Heger, L.; Dudziak, D.; Naschberger, E.; Stich, L.; Steinkasserer, A.; Kruse, F.E.; Schlötzer-Schrehardt, U. Melanocytes as Emerging Key Players in Niche Regulation of Limbal Epithelial Stem Cells. Ocul. Surf. 2021, 22, 172–189. [Google Scholar] [CrossRef]
- Polisetti, N.; Gießl, A.; Li, S.; Sorokin, L.; Kruse, F.E.; Schlötzer-Schrehardt, U. Laminin-511-E8 Promotes Efficient In Vitro Expansion of Human Limbal Melanocytes. Sci. Rep. 2020, 10, 11074. [Google Scholar] [CrossRef]
- Polisetti, N.; Sharaf, L.; Schlötzer-Schrehardt, U.; Schlunck, G.; Reinhard, T. Efficient Isolation and Functional Characterization of Niche Cells from Human Corneal Limbus. Int. J. Mol. Sci. 2022, 23, 2750. [Google Scholar] [CrossRef]
- Rama, P.; Ferrari, G.; Pellegrini, G. Cultivated Limbal Epithelial Transplantation. Curr. Opin. Ophthalmol. 2017, 28, 387–389. [Google Scholar] [CrossRef]
- Ghareeb, A.E.; Lako, M.; Figueiredo, F.C. Recent Advances in Stem Cell Therapy for Limbal Stem Cell Deficiency: A Narrative Review. Ophthalmol. Ther. 2020, 9, 809–831. [Google Scholar] [CrossRef]
- Willemsen, M.; Luiten, R.M.; Teunissen, M.B.M. Instant Isolation of Highly Purified Human Melanocytes from Freshly Prepared Epidermal Cell Suspensions. Pigment. Cell Melanoma Res. 2020, 33, 763–766. [Google Scholar] [CrossRef]
- Hayashi, R.; Yamato, M.; Sugiyama, H.; Sumide, T.; Yang, J.; Okano, T.; Tano, Y.; Nishida, K. N-Cadherin Is Expressed by Putative Stem/Progenitor Cells and Melanocytes in the Human Limbal Epithelial Stem Cell Niche. Stem Cells 2007, 25, 289–296. [Google Scholar] [CrossRef]
- Polisetti, N.; Schlötzer-Schrehardt, U.; Reinhard, T.; Schlunck, G. Isolation and Enrichment of Melanocytes from Human Corneal Limbus Using CD117 (c-Kit) as Selection Marker. Sci. Rep. 2020, 10, 17588. [Google Scholar] [CrossRef]
- Moraes, D.A.; Sibov, T.T.; Pavon, L.F.; Alvim, P.Q.; Bonadio, R.S.; Da Silva, J.R.; Pic-Taylor, A.; Toledo, O.A.; Marti, L.C.; Azevedo, R.B.; et al. A Reduction in CD90 (THY-1) Expression Results in Increased Differentiation of Mesenchymal Stromal Cells. Stem Cell Res. Ther. 2016, 7, 97. [Google Scholar] [CrossRef]
- Polisetty, N.; Fatima, A.; Madhira, S.L.; Sangwan, V.S.; Vemuganti, G.K. Mesenchymal Cells from Limbal Stroma of Human Eye. Mol. Vis. 2008, 14, 431–442. [Google Scholar]
- Li, S.; Zenkel, M.; Kruse, F.E.; Gießl, A.; Schlötzer-Schrehardt, U. Identification, Isolation, and Characterization of Melanocyte Precursor Cells in the Human Limbal Stroma. Int. J. Mol. Sci. 2022, 23, 3756. [Google Scholar] [CrossRef]
- Michalak-Mićka, K.; Büchler, V.L.; Zapiórkowska-Blumer, N.; Biedermann, T.; Klar, A.S. Characterization of a Melanocyte Progenitor Population in Human Interfollicular Epidermis. Cell Rep. 2022, 38, 110419. [Google Scholar] [CrossRef]
- Polisetti, N.; Schlunck, G.; Reinhard, T.; Kruse, F.E.; Schlötzer-Schrehardt, U. Isolation and Ex Vivo Expansion of Human Limbal Epithelial Progenitor Cells. Bio Protoc. 2020, 10, e3754. [Google Scholar] [CrossRef] [PubMed]
- Polisetti, N.; Sharaf, L.; Reinhard, T.; Schlunck, G. Isolation and Ex Vivo Expansion of Limbal Mesenchymal Stromal Cells. Bio Protoc. 2022, 12, e4471. [Google Scholar] [CrossRef] [PubMed]
- Polisetti, N.; Sharaf, L.; Martin, G.; Schlunck, G.; Reinhard, T. P-Cadherin Is Expressed by Epithelial Progenitor Cells and Melanocytes in the Human Corneal Limbus. Cells 2022, 11, 1975. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Problem Encountered | Explanations | Solutions |
|---|---|---|
| Limbal tissue is not digested completely or at all | Collagenase A might have degraded | Prepare fresh collagenase Ensure that the stock solution and collagenase A powder are stored properly |
| Incomplete digestion | Extend the incubation time but not over 20 h A higher concentration of collagenase (2%) can be used to digest limbal tissue | |
| Limbal clusters are not or only partially digested | Trypsin–EDTA might have degraded | Use fresh trypsin–EDTA Ensure proper storage of stock solution |
| Incomplete digestion | Prolong the incubation time but do not exceed 20 min | |
| Yield of sorted LMs is low | Sorted LMs might have been lost in the collection tubes | Use low-absorbing tubes |
| Quality of donor tissue | Obtain only healthy limbal tissues from younger donors (set a cutoff age of 70 y) if possible. The younger the donors, the higher the number of melanocytes | |
| A prolonged period of tissue digestion caused cell death | Do not exceed 20 h digestion in collagenase A | |
| LMs did not attach/grow | Laminin-coated plates dried | Ensure the laminin plates do not dry while coating the plates Always use freshly coated plates. If prepared in advance, keep at 4 °C for no longer than one week |
| Culture medium is not in optimal condition | Always use CnT-40 fresh medium. Do not use medium kept for more than a month at 4 °C | |
| Stromal contamination | Gating strategy is not accurate | Set the gates according to the corresponding isotype controls (0.3%) |
| Microbial contamination | Donor corneal tissue is infected | If the cornea looks cloudy/hazy, do not process |
| Contamination during cell processing | Ensure that surgical instruments are autoclaved, and all materials and reagents used are sterile Add antibiotics to the FACS buffer |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polisetti, N.; Reinhard, T.; Schlunck, G. Efficient Isolation and Expansion of Limbal Melanocytes for Tissue Engineering. Int. J. Mol. Sci. 2023, 24, 7827. https://doi.org/10.3390/ijms24097827
Polisetti N, Reinhard T, Schlunck G. Efficient Isolation and Expansion of Limbal Melanocytes for Tissue Engineering. International Journal of Molecular Sciences. 2023; 24(9):7827. https://doi.org/10.3390/ijms24097827
Chicago/Turabian StylePolisetti, Naresh, Thomas Reinhard, and Günther Schlunck. 2023. "Efficient Isolation and Expansion of Limbal Melanocytes for Tissue Engineering" International Journal of Molecular Sciences 24, no. 9: 7827. https://doi.org/10.3390/ijms24097827
APA StylePolisetti, N., Reinhard, T., & Schlunck, G. (2023). Efficient Isolation and Expansion of Limbal Melanocytes for Tissue Engineering. International Journal of Molecular Sciences, 24(9), 7827. https://doi.org/10.3390/ijms24097827