
Deregulation of the miR-19b/PPP2R5E Signaling Axis Shows High Functional Impact in Colorectal Cancer Cells

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. MiR-19b Plays a Relevant Role in Regulating the Cell Viability of CRC Cells
2.2. Significant Relevance of miR-19b-Mediated PP2A Activity Inhibition through the Direct Negative Regulation of PPP2R5E
2.3. MiR-19b Promotes Cell Proliferation in a PPP2R5E-Dependent Manner
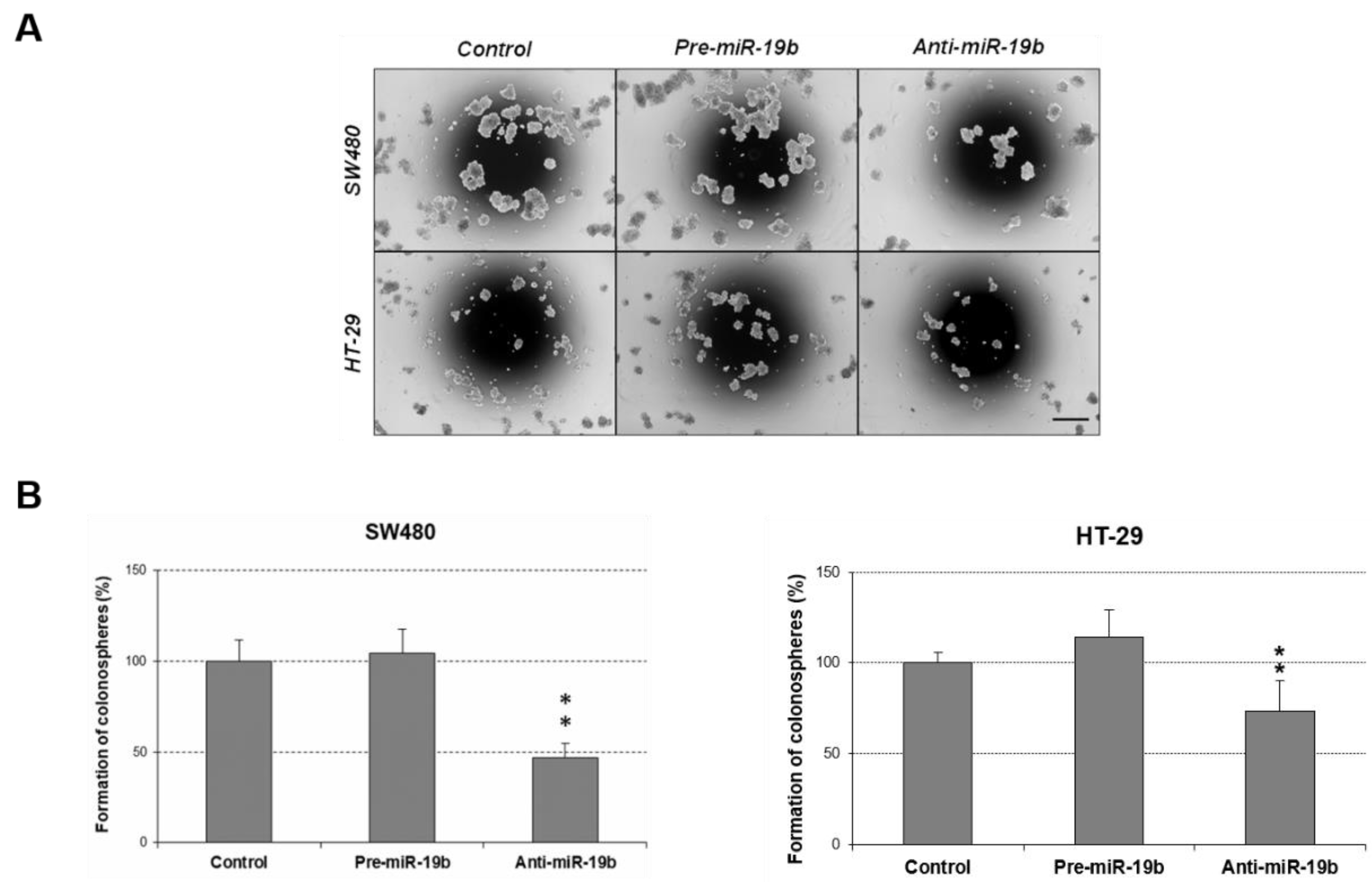
2.4. MiR-19b Regulates Cell Migration and Induces Colonosphere Formation in CRC Cells
2.5. MiR-19b/PPP2R5E Axis Regulates 5-FU Resistance in CRC Cells
3. Discussion
4. Materials and Methods
4.1. Cell Cultures and Transfection
4.2. Protein Extraction
4.3. Western Blot Analysis
4.4. Luciferase Assay
4.5. PP2A Phosphatase Activity Assay
4.6. Cell Viability Assay
4.7. Transwell Migration Assay
4.8. Colonosphere Formation
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hossain, M.S.; Karuniawati, H.; Jairoun, A.A.; Urbi, Z.; Ooi, D.J.; John, A.; Lim, Y.C.; Kaderi Kibria, K.M.; Mohiuddin, A.K.M.; Ming, L.C.; et al. Colorectal Cancer: A Review of Carcinogenesis, Global Epidemiology, Current Challenges, Risk Factors, Preventive and Treatment Strategies. Cancers 2022, 14, 1732. [Google Scholar] [CrossRef] [PubMed]
- Global Cancer Observatory. Colorectal Cancer. Available online: Https://Gco.Iarc.Fr/Today/Data/Factsheets/Cancers/10_8_9-Colorectum-Fact-Sheet.Pdf (accessed on 30 November 2022).
- Sawicki, T.; Ruszkowska, M.; Danielewicz, A.; Niedźwiedzka, E.; Arłukowicz, T.; Przybyłowicz, K.E. A Review of Colorectal Cancer in Terms of Epidemiology, Risk Factors, Development, Symptoms and Diagnosis. Cancers 2021, 13, 2025. [Google Scholar] [CrossRef] [PubMed]
- Global Cancer Observatory. Colorectal Cancer. Available online: Https://Gco.Iarc.Fr/Today/Data/Factsheets/Populations/900-World-Fact-Sheets.Pdf (accessed on 1 December 2022).
- Mattiuzzi, C.; Sanchis-Gomar, F.; Lippi, G. Concise Update on Colorectal Cancer Epidemiology. Ann. Transl. Med. 2019, 7, 609. [Google Scholar] [CrossRef] [PubMed]
- Feeney, G.; Sehgal, R.; Sheehan, M.; Hogan, A.; Regan, M.; Joyce, M.; Kerin, M. Neoadjuvant Radiotherapy for Rectal Cancer Management. World J. Gastroenterol. 2019, 25, 4850. [Google Scholar] [CrossRef] [PubMed]
- Cristóbal, I.; Santos, A.; Rubio, J.; Caramés, C.; Zazo, S.; Sanz-Álvarez, M.; Luque, M.; Madoz-Gúrpide, J.; Rojo, F.; García-Foncillas, J. Validation of MicroRNA-199b as A Promising Predictor of Outcome and Response to Neoadjuvant Treatment in Locally Advanced Rectal Cancer Patients. Cancers 2021, 13, 5003. [Google Scholar] [CrossRef] [PubMed]
- Bottarelli, L.; De Angelis, G.L.; Azzoni, C.; di Mario, F.; De Angelis, N.; Leandro, G.; Fornaroli, F.; Gaiani, F.; Negri, F. Potential Predictive Biomarkers in Locally Advanced Rectal Cancer Treated with Preoperative Chemo-Radiotherapy. Acta Bio Med. Atenei Parm. 2018, 89, 102. [Google Scholar] [CrossRef]
- Yoo, R.N.; Kim, H.J. Total Neoadjuvant Therapy in Locally Advanced Rectal Cancer: Role of Systemic Chemotherapy. Ann. Gastroenterol. Surg. 2019, 3, 356. [Google Scholar] [CrossRef]
- Peng, Y.; Croce, C.M. The Role of MicroRNAs in Human Cancer. Signal Transduct. Target. Ther. 2016, 1, 1–9. [Google Scholar] [CrossRef]
- Kong, Y.W.; Ferland-McCollough, D.; Jackson, T.J.; Bushell, M. MicroRNAs in Cancer Management. Lancet Oncol. 2012, 13, 249–258. [Google Scholar] [CrossRef]
- Strubberg, A.M.; Madison, B.B. MicroRNAs in the Etiology of Colorectal Cancer: Pathways and Clinical. Dis. Model. Mech. 2017, 10, 197. [Google Scholar] [CrossRef]
- Sur, D.; Advani, S.; Braithwaite, D. MicroRNA Panels as Diagnostic Biomarkers for Colorectal Cancer: A Systematic Review and Meta-Analysis. Front. Med. 2022, 9, 915226. [Google Scholar] [CrossRef] [PubMed]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.X.; Zhang, B.; Wei, J.; Jiang, G.Q.; Wu, Y.L.; Leng, B.J.; Xing, C.G. MiR-539 Inhibits Proliferation and Migration of Triple-Negative Breast Cancer Cells by down-Regulating LAMA4 Expression. Cancer Cell Int. 2018, 18, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hujie, G.; Zhou, S.H.; Zhang, H.; Qu, J.; Xiong, X.W.; Hujie, O.; Gong, L.C.; Yang, S.E. MicroRNA-10b Regulates Epithelial-Mesenchymal Transition by Modulating KLF4/KLF11/Smads in Hepatocellular Carcinoma. Cancer Cell Int. 2018, 18, 10. [Google Scholar] [CrossRef]
- Sohn, E.J. MicroRNA 200c-3p Regulates Autophagy via Upregulation of Endoplasmic Reticulum Stress in PC-3 Cells. Cancer Cell Int. 2018, 18, 2. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.T.; Nian, X.X.; Wang, S.Y.; Jiao, H.L.; Wang, Y.X.; Xiao, Z.Y.; Yang, R.W.; Ding, Y.Q.; Ye, Y.P.; Liao, W.T. MiR-422a Inhibits Cell Proliferation in Colorectal Cancer by Targeting AKT1 and MAPK1. Cancer Cell Int. 2017, 17, 91. [Google Scholar] [CrossRef]
- Fiori, M.E.; Villanova, L.; Barbini, C.; De Angelis, M.L.; De Maria, R. MiR-663 Sustains NSCLC by Inhibiting Mitochondrial Outer Membrane Permeabilization (MOMP) through PUMA/BBC3 and BTG2. Cell Death Dis. 2018, 9, 49. [Google Scholar] [CrossRef]
- Olive, V.; Bennett, M.J.; Walker, J.C.; Ma, C.; Jiang, I.; Cordon-Cardo, C.; Li, Q.J.; Lowe, S.W.; Hannon, G.J.; He, L. MiR-19 Is a Key Oncogenic Component of Mir-17-92. Genes Dev. 2009, 23, 2839. [Google Scholar] [CrossRef]
- Baumgartner, U.; Berger, F.; Hashemi Gheinani, A.; Burgener, S.S.; Monastyrskaya, K.; Vassella, E. MiR-19b Enhances Proliferation and Apoptosis Resistance via the EGFR Signaling Pathway by Targeting PP2A and BIM in Non-Small Cell Lung Cancer. Mol. Cancer 2018, 17, 44. [Google Scholar] [CrossRef]
- Li, J.; Yang, S.; Yan, W.; Yang, J.; Qin, Y.J.; Lin, X.L.; Xie, R.Y.; Wang, S.C.; Jin, W.; Gao, F.; et al. MicroRNA-19 Triggers Epithelial-Mesenchymal Transition of Lung Cancer Cells Accompanied by Growth Inhibition. Lab. Investig. 2015, 95, 1056–1070. [Google Scholar] [CrossRef]
- Yuan, J.; Su, Z.; Gu, W.; Shen, X.; Zhao, Q.; Shi, L.; Jin, C.; Wang, X.; Cong, H.; Ju, S. MiR-19b and MiR-20a Suppress Apoptosis, Promote Proliferation and Induce Tumorigenicity of Multiple Myeloma Cells by Targeting PTEN. Cancer Biomark. 2019, 24, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Yin, S.; Hao, Y.; Yang, J.; Zhang, H.; Sun, C.; Ma, M.; Chang, Q.; Xi, J.J. MiR-19b Promotes Tumor Growth and Metastasis via Targeting TP53. RNA 2014, 20, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Cai, J.L.; Huang, P.Z.; Kang, L.; Huang, M.J.; Wang, L.; Wang, J.P. MiR19b-3p Promotes the Growth and Metastasis of Colorectal Cancer via Directly Targeting ITGB8. Am. J. Cancer Res. 2017, 7, 1996. [Google Scholar] [PubMed]
- Jiang, T.; Ye, L.; Han, Z.; Liu, Y.; Yang, Y.; Peng, Z.; Fan, J. MiR-19b-3p Promotes Colon Cancer Proliferation and Oxaliplatin-Based Chemoresistance by Targeting SMAD4: Validation by Bioinformatics and Experimental Analyses. J. Exp. Clin. Cancer Res. 2017, 36, 131. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, K.; Tanahashi, T.; Iima, T.; Yamamoto, Y.; Akaike, Y.; Nishida, K.; Masuda, K.; Kuwano, Y.; Murakami, Y.; Fukushima, M.; et al. Role of MiR-19b and Its Target MRNAs in 5-Fluorouracil Resistance in Colon Cancer Cells. J. Gastroenterol. 2012, 47, 883–895. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Z.; Han, X.; Jiang, L.; Ge, R.; Wang, X.; Li, J. Up-Regulation of MicroRNA-19b Is Associated with Metastasis and Predicts Poor Prognosis in Patients with Colorectal Cancer. Int. J. Clin. Exp. Pathol. 2018, 11, 3952. [Google Scholar]
- Rubio, J.; Cristóbal, I.; Santos, A.; Caramés, C.; Luque, M.; Sanz-Alvarez, M.; Zazo, S.; Madoz-Gúrpide, J.; Rojo, F.; García-Foncillas, J. Low MicroRNA-19b Expression Shows a Promising Clinical Impact in Locally Advanced Rectal Cancer. Cancers 2021, 13, 1456. [Google Scholar] [CrossRef]
- Santos, A.; Cristóbal, I.; Rubio, J.; Caramés, C.; Luque, M.; Sanz-Álvarez, M.; Zazo, S.; Madoz-Gúrpide, J.; Rojo, F.; García-Foncillas, J. MicroRNA-19b Plays a Key Role in 5-Fluorouracil Resistance and Predicts Tumor Progression in Locally Advanced Rectal Cancer Patients. Int. J. Mol. Sci. 2022, 23, 12447. [Google Scholar] [CrossRef]
- Mavrakis, K.J.; Wolfe, A.L.; Oricchio, E.; Palomero, T.; de Keersmaecker, K.; McJunkin, K.; Zuber, J.; James, T.; Chang, K.; Khan, A.A.; et al. Genome-Wide RNA-Mediated Interference Screen Identifies MiR-19 Targets in Notch-Induced T-Cell Acute Lymphoblastic Leukaemia. Nat. Cell Biol. 2010, 12, 372–379. [Google Scholar] [CrossRef]
- Cristóbal, I.; Manso, R.; Rincón, R.; Caramés, C.; Senin, C.; Borrero, A.; Martínez-Useros, J.; Rodriguez, M.; Zazo, S.; Aguilera, O.; et al. PP2A Inhibition Is a Common Event in Colorectal Cancer and Its Restoration Using FTY720 Shows Promising Therapeutic Potential. Mol. Cancer Ther. 2014, 13, 938–947. [Google Scholar] [CrossRef]
- Cristóbal, I.; Rincón, R.; Manso, R.; Caramés, C.; Zazo, S.; Madoz-Gúrpide, J.; Rojo, F.; García-Foncillas, J. Deregulation of the PP2A Inhibitor SET Shows Promising Therapeutic Implications and Determines Poor Clinical Outcome in Patients with Metastatic Colorectal Cancer. Clin. Cancer Res. 2015, 21, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Cristóbal, I.; Rincón, R.; Manso, R.; Madoz-Gúrpide, J.; Caramés, C.; del Puerto-Nevado, L.; Rojo, F.; García-Foncillas, J. Hyperphosphorylation of PP2A in Colorectal Cancer and the Potential Therapeutic Value Showed by Its Forskolin-Induced Dephosphorylation and Activation. Biochim. Biophys. Acta 2014, 1842, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Cristóbal, I.; Torrejón, B.; Rubio, J.; Santos, A.; Pedregal, M.; Caramés, C.; Zazo, S.; Luque, M.; Sanz-Alvarez, M.; Madoz-Gúrpide, J.; et al. Deregulation of SET Is Associated with Tumor Progression and Predicts Adverse Outcome in Patients with Early-Stage Colorectal Cancer. J. Clin. Med. 2019, 8, 346. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.J.; Netz, U.; Hallion, J.; Bishop, C.; Stephen, V.; Burton, J.; Paas, M.; Feagins, K.; Pan, J.; Rai, S.N.; et al. Circulating Plasma MicroRNAs in Colorectal Neoplasia: A Pilot Study in Assessing Response to Therapy. Transl. Oncol. 2021, 14, 100962. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, A.; Cristóbal, I.; Caramés, C.; Luque, M.; Sanz-Álvarez, M.; Madoz-Gúrpide, J.; Rojo, F.; García-Foncillas, J. Deregulation of the miR-19b/PPP2R5E Signaling Axis Shows High Functional Impact in Colorectal Cancer Cells. Int. J. Mol. Sci. 2023, 24, 7779. https://doi.org/10.3390/ijms24097779
Santos A, Cristóbal I, Caramés C, Luque M, Sanz-Álvarez M, Madoz-Gúrpide J, Rojo F, García-Foncillas J. Deregulation of the miR-19b/PPP2R5E Signaling Axis Shows High Functional Impact in Colorectal Cancer Cells. International Journal of Molecular Sciences. 2023; 24(9):7779. https://doi.org/10.3390/ijms24097779
Chicago/Turabian StyleSantos, Andrea, Ion Cristóbal, Cristina Caramés, Melani Luque, Marta Sanz-Álvarez, Juan Madoz-Gúrpide, Federico Rojo, and Jesús García-Foncillas. 2023. "Deregulation of the miR-19b/PPP2R5E Signaling Axis Shows High Functional Impact in Colorectal Cancer Cells" International Journal of Molecular Sciences 24, no. 9: 7779. https://doi.org/10.3390/ijms24097779
APA StyleSantos, A., Cristóbal, I., Caramés, C., Luque, M., Sanz-Álvarez, M., Madoz-Gúrpide, J., Rojo, F., & García-Foncillas, J. (2023). Deregulation of the miR-19b/PPP2R5E Signaling Axis Shows High Functional Impact in Colorectal Cancer Cells. International Journal of Molecular Sciences, 24(9), 7779. https://doi.org/10.3390/ijms24097779