

β-Cyclodextrin Polymer-Based Fluorescence Enhancement Strategy via Host–Guest Interaction for Sensitive Assay of SARS-CoV-2


Abstract
1. Introduction
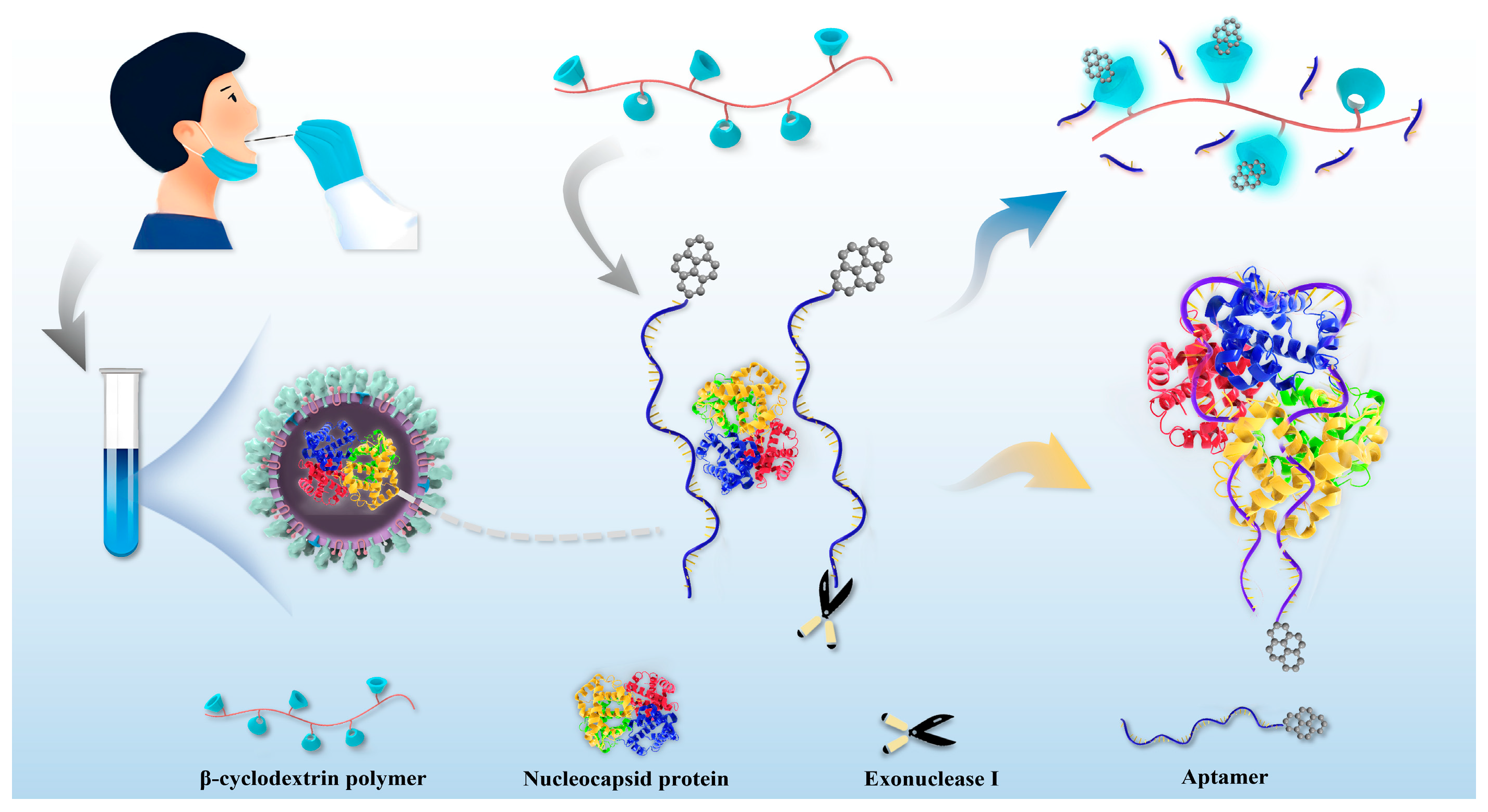
2. Results and Discussion
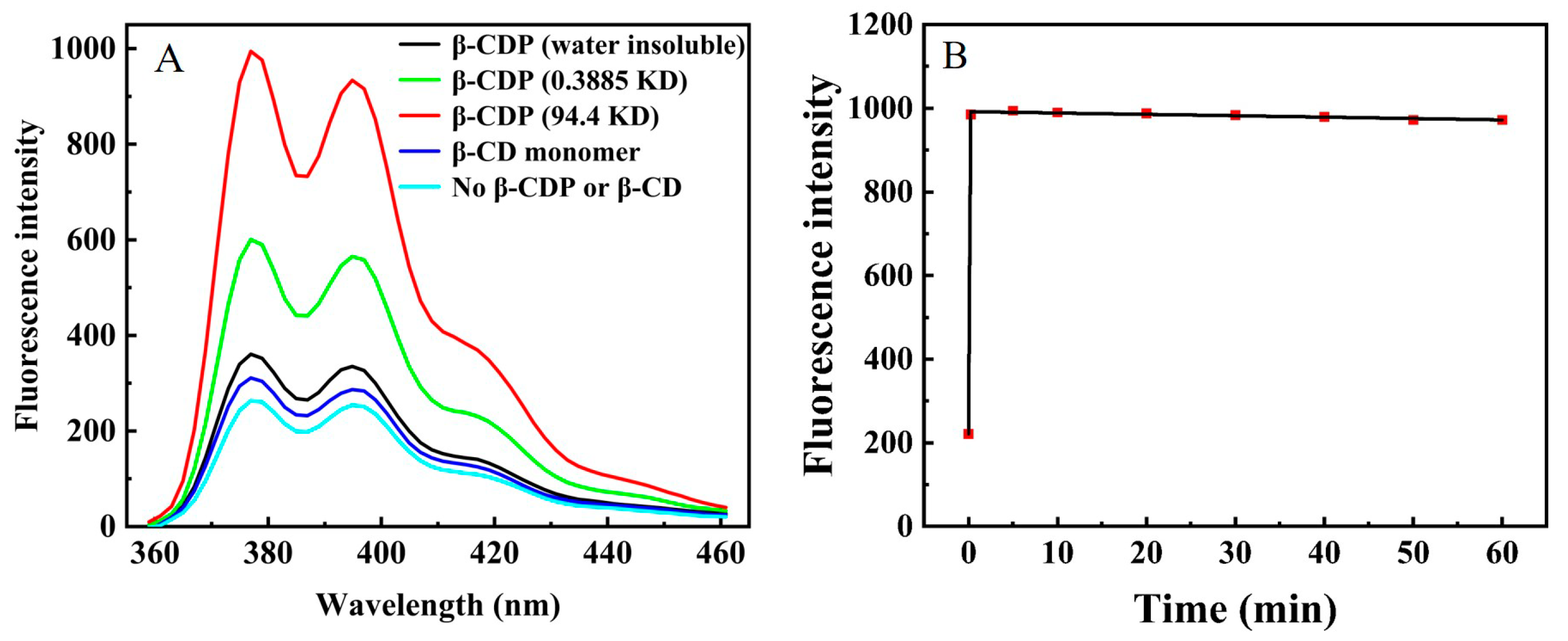
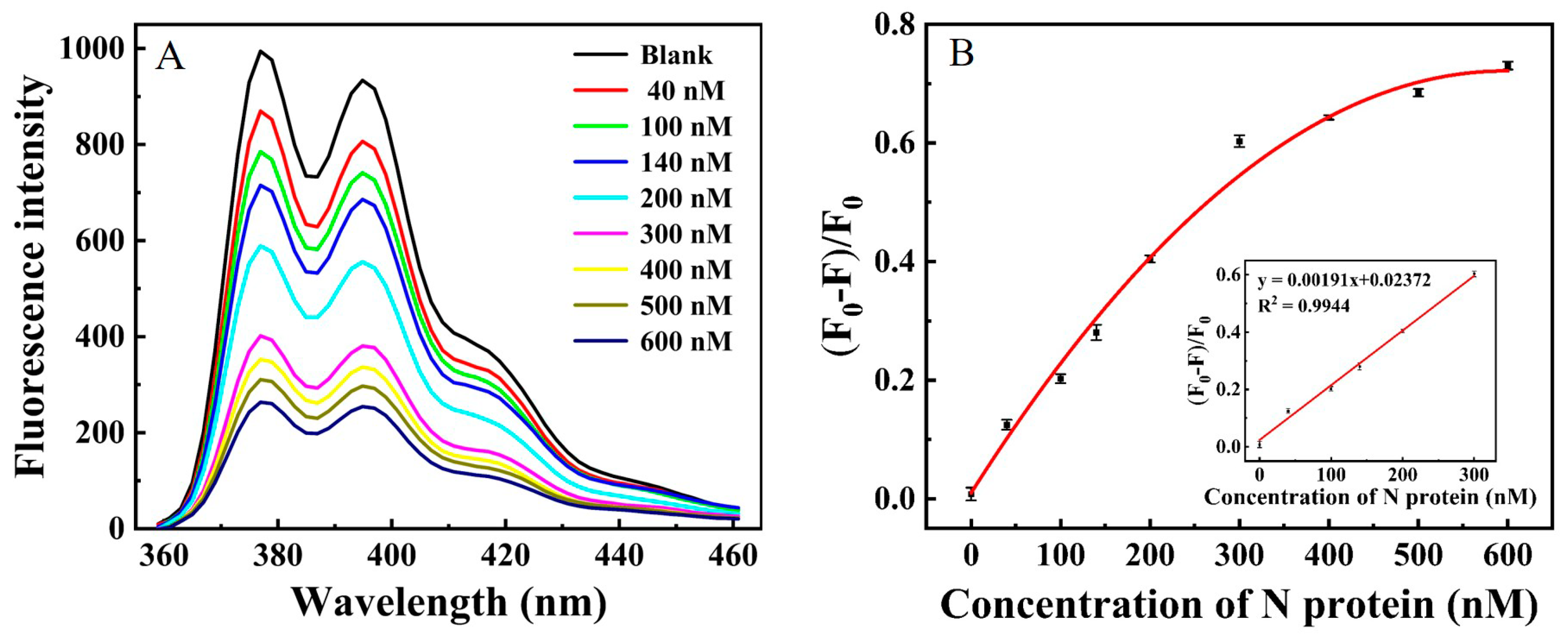
2.1. The Fluorescence Enhancement of β-Cyclodextrin Polymer
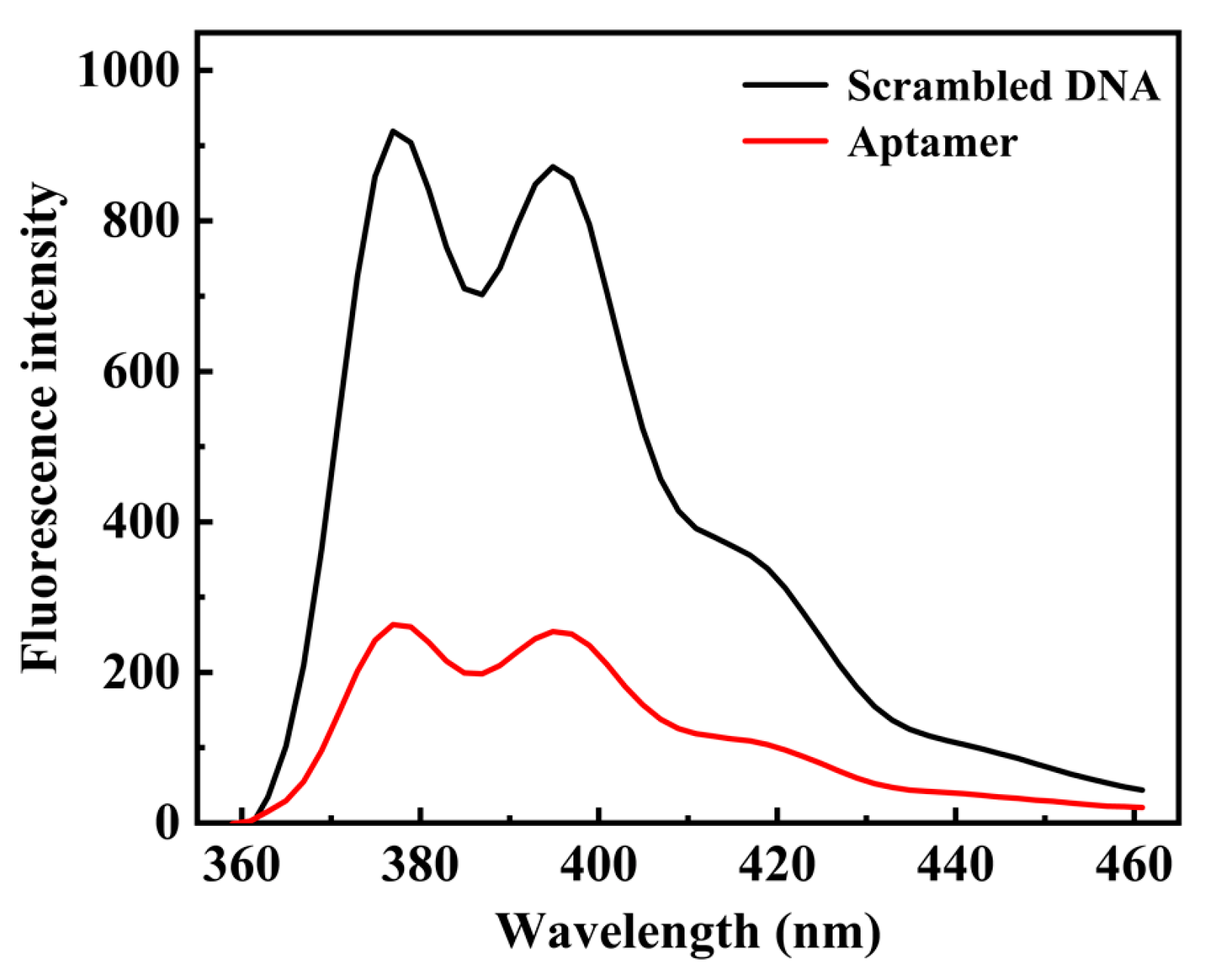
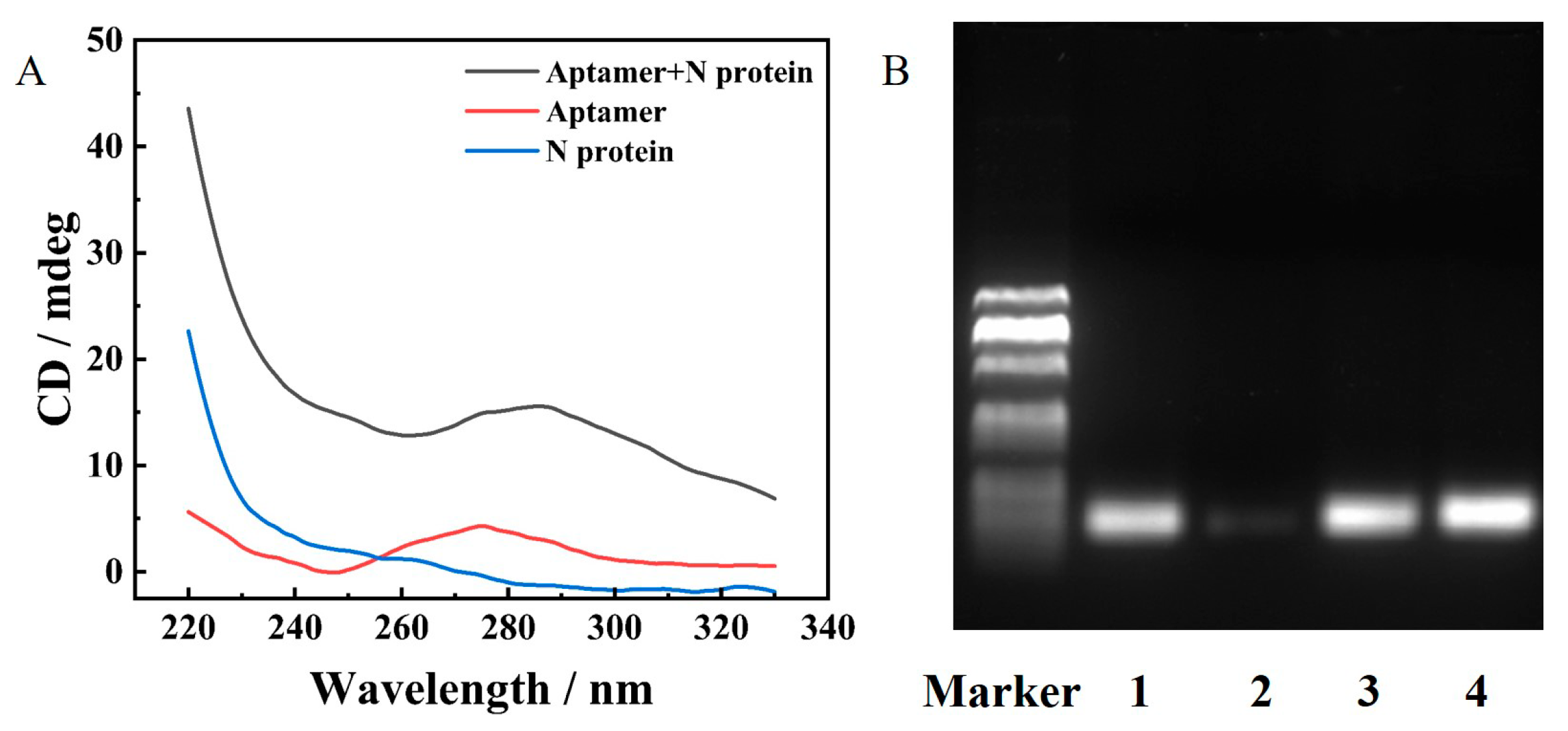
2.2. The Performance of N Protein Aptamer
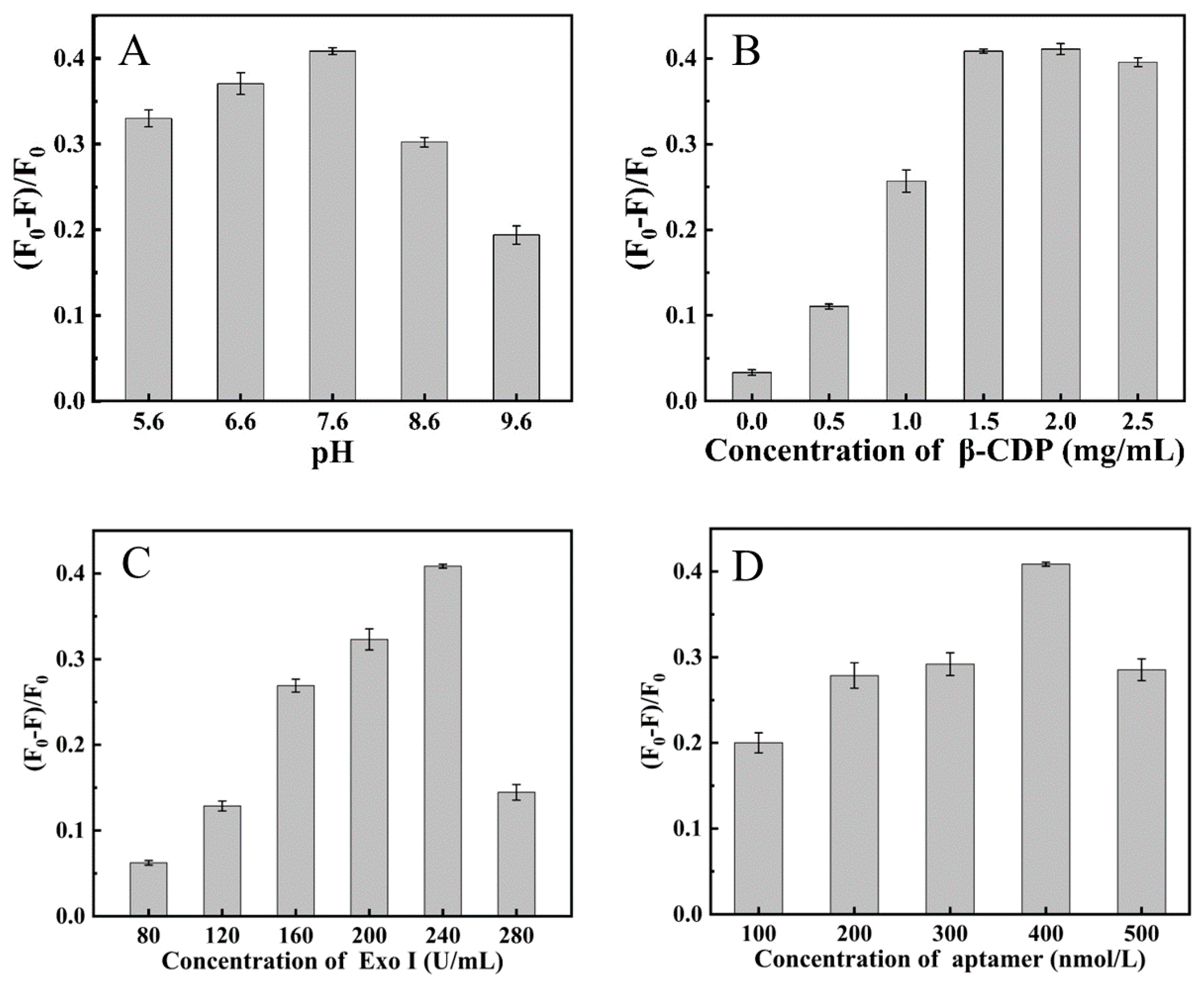
2.3. Optimization Experiment
2.4. Analytical Performance
2.5. Selectivity
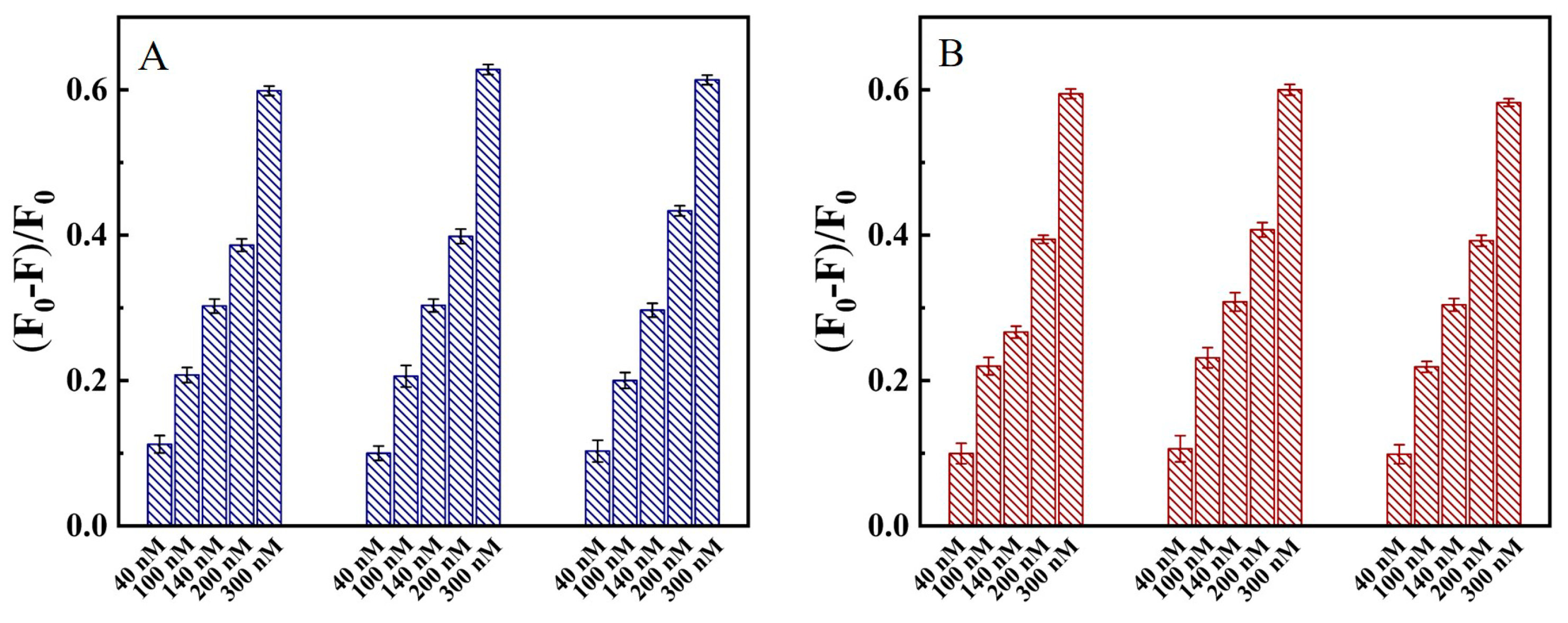
2.6. N Protein Analysis in Clinical Blood and Throat Swabs Samples
2.7. The Recovery of N Protein in Clinical Blood and Swabs Sample
3. Materials and Methods
3.1. Materials
3.2. DNA Sequences Used in this Work
3.3. Preparation of β-Cyclodextrin Polymer (β-CDP)
3.4. N Protein Detection
3.5. Investigation of Assay Selectivity
3.6. Detection of N Protein in Clinical Blood and Swab Samples
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Z.; Morrissey, J.J. Plasmonically enhanced ultrasensitive epitope-specific serologic assay for COVID-19. Anal. Chem. 2022, 94, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Rösch, E.L.; Viereck, T.; Schilling, M.; Ludwig, F. Toward rapid and sensitive detection of sars-cov-2 with functionalized magnetic nanoparticles. ACS Sens. 2021, 6, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in china. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A novel coronavirus from patients with pneumonia in china 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Hellewell, J.; Abbott, S.; Gimma, A.; Bosse, N.I.; Jarvis, C.I.; Russell, T.W.; Munday, J.D.; Kucharski, A.J.; Edmunds, W.J.; Funk, S.; et al. Feasibility of controlling COVID-19 outbreaks by isolation of cases and contacts. Lancet Glob. Health 2020, 8, e488–e496. [Google Scholar] [CrossRef]
- Lukose, J.; Chidangil, S.; George, S.D. Optical technologies for the detection of viruses like COVID-19: Progress and prospects. Biosens. Bioelectron. 2021, 178, 113004. [Google Scholar] [CrossRef]
- Cho, S.Y.; Jin, X.; Gong, X. Antibody-free rapid detection of SARS-CoV-2 proteins using corona phase molecular recognition to accelerate development time. Anal. Chem. 2021, 93, 14685–14693. [Google Scholar] [CrossRef]
- Udugama, B.; Kadhiresan, P.; Kozlowski, H.N.; Malekjahani, A.; Li, M.; Osborne, V.Y.C.; Chen, H.; Mubareka, S.; Gubbay, J.B.; Chan, W.C.W. Diagnosing COVID-19: The disease and tools for detection. ACS Nano 2020, 14, 3822–3835. [Google Scholar] [CrossRef]
- Shi, L.; Wang, L.; Ma, X.; Fang, X.; Xiang, L.; Yi, Y.; Li, J.; Luo, Z.; Li, G. Aptamer-functionalized nanochannels for one-step detection of SARS-CoV-2 in samples from COVID-19 patients. Anal. Chem. 2021, 93, 16646–16654. [Google Scholar] [CrossRef]
- Eissa, S.; Zourob, M. Development of a low-cost cotton-tipped electrochemical immunosensor for the detection of SARS-CoV-2. Anal. Chem. 2021, 93, 1826–1833. [Google Scholar] [CrossRef]
- Wei, C.; Yang, H.; Wang, S.; Zhao, J.; Liu, C.; Gao, L.; Xia, E.; Lu, Y.; Tai, Y.; She, G.; et al. Draft genome sequence of camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA 2018, 115, E4151–E4158. [Google Scholar] [CrossRef]
- Peng, Y.; Chen, L.; Li, S.; Zhang, Y.; Xu, R.; Liu, Z.; Liu, W.; Kong, J.; Huang, X.; Wang, Y.; et al. BRI1 and BAK1 interact with G proteins and regulate sugar-responsive growth and development in Arabidopsis. Nat. Commun. 2018, 9, 1522. [Google Scholar] [CrossRef]
- Kumar, S.; Nyodu, R.; Maurya, V.K.; Saxena, S.K. Morphology, genome organization, replication, and pathogenesis of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). In Coronavirus Disease 2019 (COVID-19): Epidemiology, Pathogenesis, Diagnosis, and Therapeutics; Springer: Singapore, 2020; Volume 3, pp. 23–31. [Google Scholar]
- Le Bert, N.; Tan, A.; Kunasegaran, K.; Tham, C.; Hafezi, M.; Chia, A.; Chng, M.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-Specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef]
- Li, T.; Wang, L.; Wang, H.; Li, X.; Wei, W. Serum SARS-COV-2 nucleocapsid protein: A sensitivity and specificity early diagnostic marker for SARS-COV-2 infection. Front. Cell Infect. Microbiol. 2020, 10, 470. [Google Scholar] [CrossRef]
- Li, J.; Lillehoj, P.B. Microfluidic Magneto Immunosensor for Rapid, High sensitivity measurements of SARS-CoV-2 nucleocapsid protein in serum. ACS Sens. 2021, 6, 1270–1278. [Google Scholar] [CrossRef]
- Yu, M.; Zhang, X.; Zhang, X.; Zahra, Q.U.A.; Huang, Z.; Chen, Y.; Song, C.; Song, M.; Jiang, H.; Luo, Z.; et al. An electrochemical aptasensor with N protein binding aptamer-complementary oligonucleotide as probe for ultra-sensitive detection of COVID-19. Biosens. Bioelectron. 2022, 213, 114436. [Google Scholar] [CrossRef]
- Chen, J.; Tang, J.; Meng, H.; Liu, Z.; Wang, L.; Geng, X.; Wu, Y.; Qu, L.; Li, Z. Recognition triggered assembly of split aptamers to initiate a hybridization chain reaction for wash-free and amplified detection of exosomes. Chem. Commun. 2020, 56, 9024–9027. [Google Scholar] [CrossRef]
- Ahn, D.G.; Jeon, I.J.; Kim, J.; Song, M.S.; Han, S.R. RNA aptamer-based sensitive detection of SARS coronavirus nucleocapsid protein. Analyst 2009, 134, 1896–1901. [Google Scholar] [CrossRef]
- Hong, S.L.; Xiang, M.Q.; Tang, M.; Pang, D.W. Ebola virus aptamers: From highly efficient selection to application on magnetism-controlled chips. Anal. Chem. 2019, 91, 3367–3373. [Google Scholar] [CrossRef]
- Bhardwaj, J.; Chaudhary, N.; Kim, H.; Jang, J. Subtyping of influenza a H1N1 virus using a label-free electrochemical biosensor based on the DNA aptamer targeting the stem region of HA protein. Anal. Chim. Acta 2019, 1064, 94–103. [Google Scholar] [CrossRef]
- Zhang, L.; Fang, X.; Liu, X.; Ou, H.; Luo, Z. Discovery of the sandwich type COVID-19 nucleocapsid protein DNA aptamers. Chem. Commun. 2020, 56, 10235–10238. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Sun, S.; Guo, X.; Yang, X.; Huang, J.; Wang, K.; Wang, Q.; Liu, J.; He, L. Competitive host-guest interaction between beta_cyclodextrin polymer and pyrene-labeled probes for fluorescence analyses. Anal. Chem. 2015, 87, 2665–2671. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Yang, G.; Zhang, X.; Lu, Y.; Chen, Y.; Wu, X.; Song, C. β-Cyclodextrin polymer-based host–guest interaction and fluorescence enhancement of pyrene for sensitive isocarbophos detection. ACS Omega 2022, 7, 12747–12752. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Yang, X.; Wang, K.; Wang, Q.; Liu, J.; Huang, J.; He, L.; Liu, P.; Qing, Z.; Liu, W. A Sensitive detection of T4 polynucleotide kinase activity based on beta-cyclodextrin polymer enhanced fluorescence combined with an exonuclease reaction. Chem. Commun. 2015, 51, 1815–1818. [Google Scholar] [CrossRef]
- Song, C.; Xiao, Y.; Li, K.; Zhang, X.; Lu, Y. Beta-cyclodextrin polymer based fluorescence enhancement method for sensitive adenosine triphosphate detection. Chin. Chem. Lett. 2019, 30, 1249–1252. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volunteer | Sex | Age | Height (cm) | Weight (kg) | Healthy | Nucleic Acid Test Results Within 48 h |
|---|---|---|---|---|---|---|
| 1 | Female | 22 | 158 | 45 | Yes | negative |
| 2 | Male | 25 | 174 | 63 | Yes | negative |
| 3 | Female | 42 | 167 | 53 | Yes | negative |
| Spiked (nM) | Blood Samples | Throat Swabs | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Recovery (%) | RSD (%) | Recovery (%) | RSD (%) | |||||||
| 1 | 2 | 3 | Average (%) | 1 | 2 | 3 | Average (%) | |||
| 40 | 108.1 | 101.8 | 108.6 | 106.2 | 3.8 | 99.3 | 107.4 | 98.0 | 101.6 | 5.1 |
| 100 | 96.3 | 103.6 | 92.3 | 97.4 | 5.7 | 102.7 | 108.7 | 103.4 | 104.9 | 3.3 |
| 140 | 103.6 | 105.5 | 102.3 | 103.8 | 1.6 | 106.4 | 90.83 | 104.8 | 100.7 | 8.6 |
| 200 | 94.9 | 104.4 | 107.3 | 102.2 | 6.5 | 97.02 | 100.4 | 96.5 | 98.0 | 2.1 |
| 300 | 100.3 | 105.9 | 103.0 | 103.1 | 2.8 | 100.5 | 99.607 | 97.5 | 99.2 | 1.6 |
| Name | Sequence (5′ to 3′) |
|---|---|
| N protein-binding aptamer | 5′-GCTGGATGTCGCTTACGACAATATTCCTTAGGGGCACCGCTACATTGACACATCCAGC-3′ |
| Pyrene-labeled N protein binding aptamer | 5′-SH-GCTGGATGTCGCTTACGACAATATTCCTTAGGGGCACCGCTACATTGACACATCCAGC-pyrene-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, S.; Yang, G.; Zhang, X.; Shi, R.; Chen, R.; Zhang, X.; Peng, Y.; Yang, H.; Lu, Y.; Song, C. β-Cyclodextrin Polymer-Based Fluorescence Enhancement Strategy via Host–Guest Interaction for Sensitive Assay of SARS-CoV-2. Int. J. Mol. Sci. 2023, 24, 7174. https://doi.org/10.3390/ijms24087174
Gao S, Yang G, Zhang X, Shi R, Chen R, Zhang X, Peng Y, Yang H, Lu Y, Song C. β-Cyclodextrin Polymer-Based Fluorescence Enhancement Strategy via Host–Guest Interaction for Sensitive Assay of SARS-CoV-2. International Journal of Molecular Sciences. 2023; 24(8):7174. https://doi.org/10.3390/ijms24087174
Chicago/Turabian StyleGao, Shanshan, Gege Yang, Xiaohui Zhang, Rui Shi, Rongrong Chen, Xin Zhang, Yuancheng Peng, Hua Yang, Ying Lu, and Chunxia Song. 2023. "β-Cyclodextrin Polymer-Based Fluorescence Enhancement Strategy via Host–Guest Interaction for Sensitive Assay of SARS-CoV-2" International Journal of Molecular Sciences 24, no. 8: 7174. https://doi.org/10.3390/ijms24087174
APA StyleGao, S., Yang, G., Zhang, X., Shi, R., Chen, R., Zhang, X., Peng, Y., Yang, H., Lu, Y., & Song, C. (2023). β-Cyclodextrin Polymer-Based Fluorescence Enhancement Strategy via Host–Guest Interaction for Sensitive Assay of SARS-CoV-2. International Journal of Molecular Sciences, 24(8), 7174. https://doi.org/10.3390/ijms24087174