

Mass Spectrometry-Based Untargeted Approaches to Reveal Diagnostic Signatures of Male Infertility in Seminal Plasma: A New Laboratory Perspective for the Clinical Management of Infertility?

 ,
, 
Abstract
1. Introduction
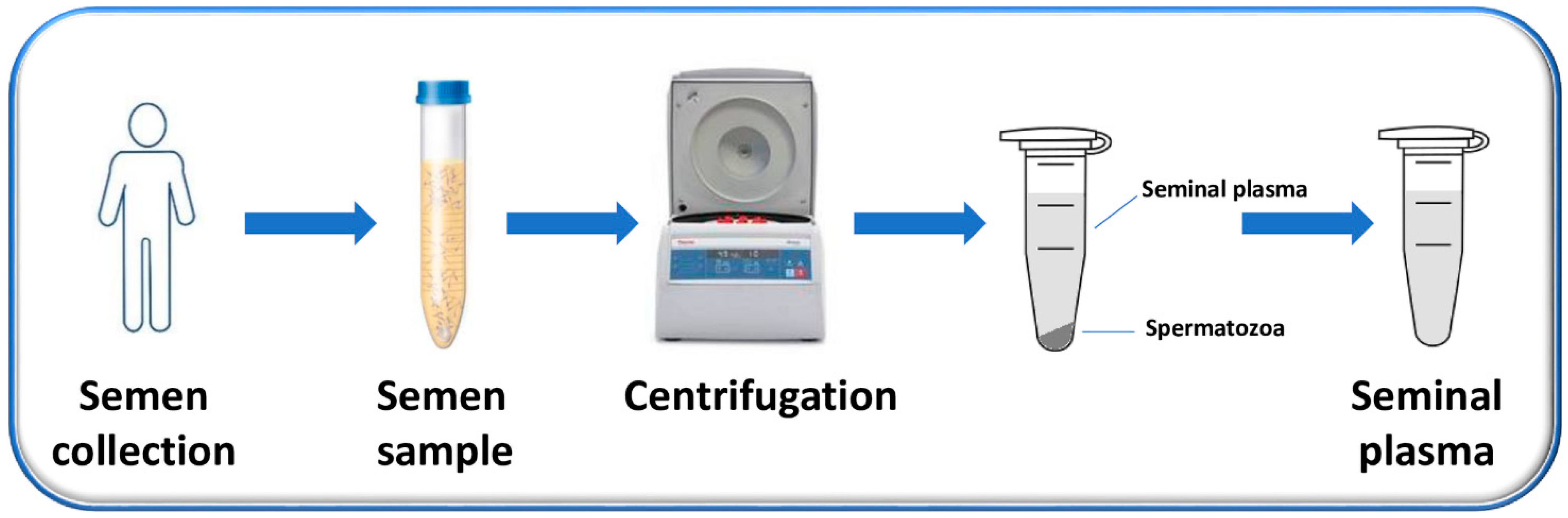
2. Semen and Seminal Plasma
2.1. Human Semen and Its Composition
2.2. Features of Human Seminal Plasma
2.3. Importance and Advantages of SP as Biomarker Source of Male Reproductive System Disorders
2.4. Assessment of Proteolytic Activity in SP by MS
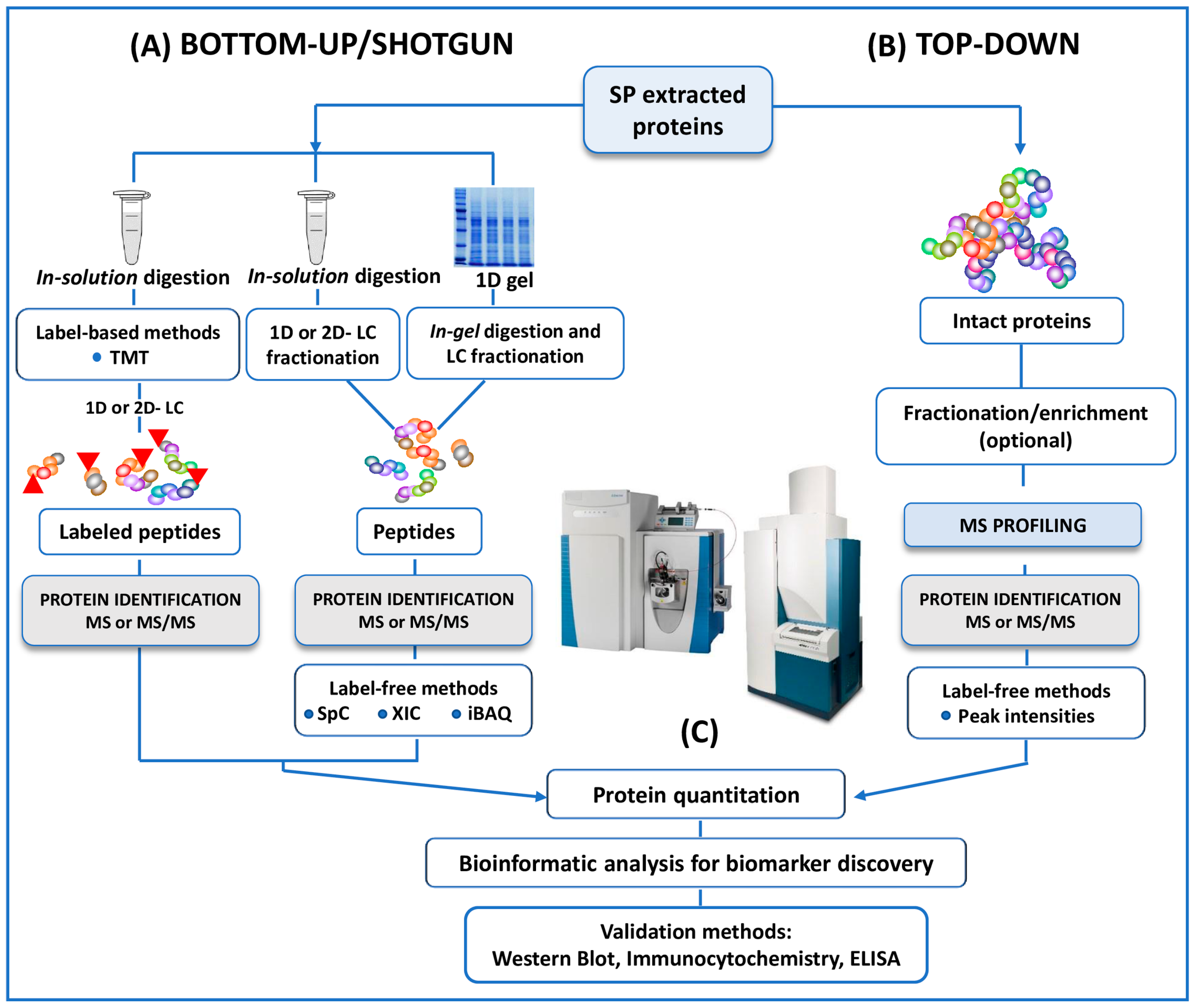
3. MS Untargeted Approaches for Differential Proteomics Analysis between Fertile and Infertile Subjects
3.1. SP Proteome Analysis by Bottom-Up Approach
3.2. SP Proteome Analysis by Top-Down Approach
4. Relevance of Pre-Analytical and Analytical Issues for SP Proteome Profiling and Identification of Male Infertility Biomarkers
4.1. Pre-Analytical Issues
4.2. Intra and Inter-Individual Variability Related Issues
4.3. Quantitation Issues
4.4. Bottom-Up vs. Top-Down Approaches
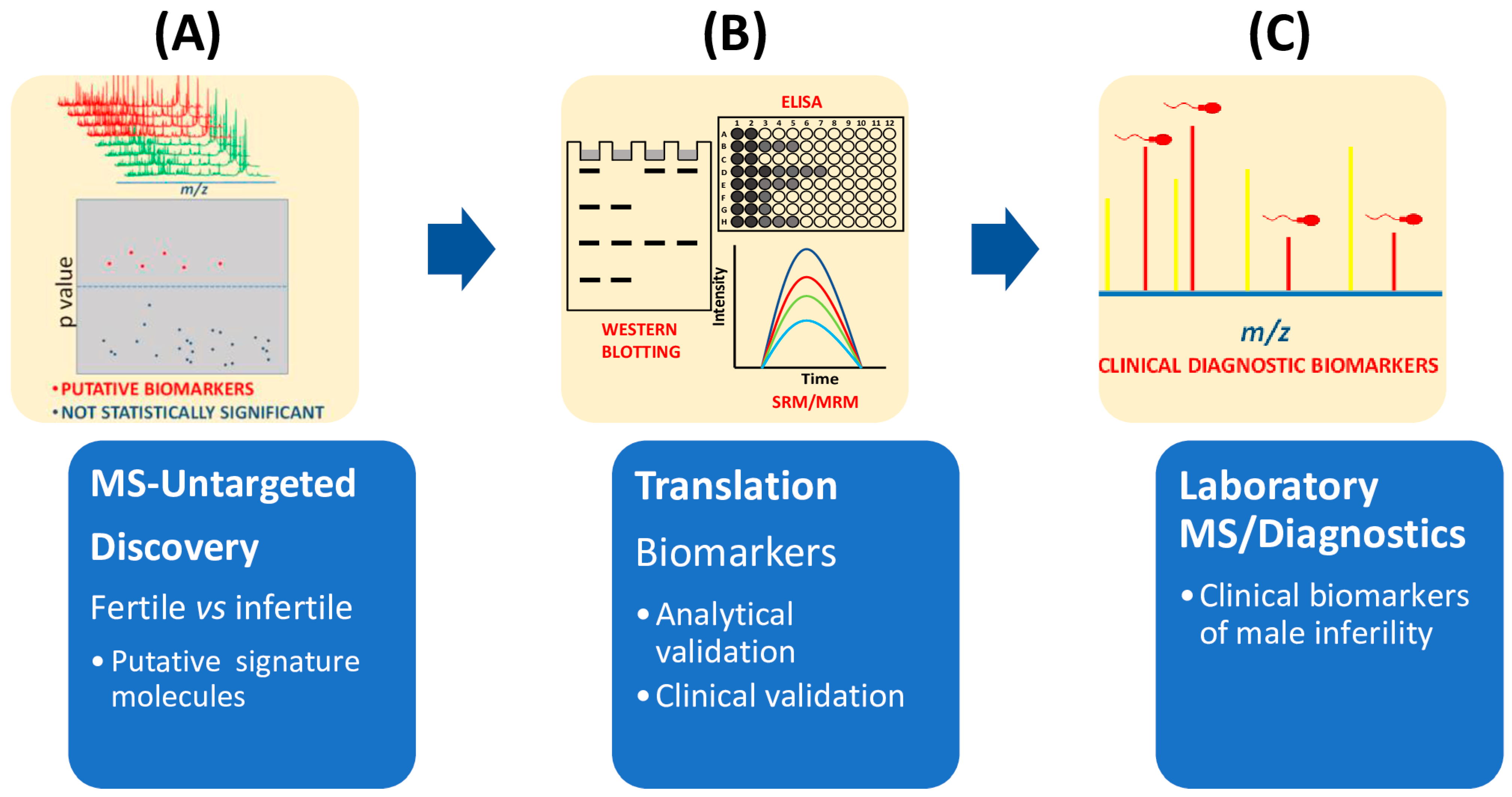
4.5. Bioinformatic Interpretation of Collected Data
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. International Classification of Diseases, 11th Revision (ICD-11). Available online: https://www.who.int/news-room/fact-sheets/detail/infertility (accessed on 2 November 2022).
- Ferlin, A.; Foresta, C. Infertility: Practical Clinical Issues for Routine Investigation of the Male Partner. J. Clin. Med. 2020, 9, 1644. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Baskaran, S.; Parekh, N.; Cho, C.L.; Henkel, R.; Vij, S.; Arafa, M.; Panner Selvam, M.K.; Shah, R. Male infertility. Lancet 2021, 397, 319–333. [Google Scholar] [CrossRef] [PubMed]
- WHO. Laboratory Manual for the Examination and Processing of Human Semen. Available online: https://www.who.int/publications/i/item/9789240030787 (accessed on 2 November 2022).
- Boitrelle, F.; Shah, R.; Saleh, R.; Henkel, R.; Kandil, H.; Chung, E.; Vogiatzi, P.; Zini, A.; Arafa, M.; Agarwal, A. The Sixth Edition of the WHO Manual for Human Semen Analysis: A Critical Review and SWOT Analysis. Life 2021, 11, 1368. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Swerdloff, R.S. Limitations of semen analysis as a test of male fertility and anticipated needs from newer tests. Fertil. Steril. 2014, 102, 1502–1507. [Google Scholar] [CrossRef] [PubMed]
- Barbăroșie, C.; Agarwal, A.; Henkel, R. Diagnostic value of advanced semen analysis in evaluation of male infertility. Andrologia 2021, 53, e13625. [Google Scholar] [CrossRef] [PubMed]
- Panner Selvam, M.K.; Agarwal, A.; Pushparaj, P.N.; Baskaran, S.; Bendou, H. Sperm Proteome Analysis and Identification of Fertility-Associated Biomarkers in Unexplained Male Infertility. Genes 2019, 10, 522. [Google Scholar] [CrossRef]
- Hamada, A.; Esteves, S.C.; Nizza, M.; Agarwal, A. Unexplained male infertility: Diagnosis and management. Int. Braz J. Urol. 2012, 38, 576–594. [Google Scholar] [CrossRef]
- Oehninger, S.; Ombelet, W. Limits of current male fertility testing. Fertil. Steril. 2019, 111, 835–841. [Google Scholar] [CrossRef]
- Bieniek, J.M.; Drabovich, A.P.; Lo, K.C. Seminal biomarkers for the evaluation of male infertility. Asian J. Androl. 2016, 18, 426–433. [Google Scholar] [CrossRef]
- Llavanera, M.; Delgado-Bermúdez, A.; Ribas-Maynou, J.; Salas-Huetos, A.; Yeste, M. A systematic review identifying fertility biomarkers in semen: A clinical approach through Omics to diagnose male infertility. Fertil. Steril. 2022, 118, 291–313. [Google Scholar] [CrossRef]
- Panner Selvam, M.K.; Agarwal, A. Update on the proteomics of male infertility: A systematic review. Arab J. Urol. 2017, 16, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Candenas, L.; Chianese, R. Exosome Composition and Seminal Plasma Proteome: A Promising Source of Biomarkers of Male Infertility. Int. J. Mol. Sci. 2020, 21, 7022. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Panner Selvam, M.K.; Baskaran, S. Proteomic Analyses of Human Sperm Cells: Understanding the Role of Proteins and Molecular Pathways Affecting Male Reproductive Health. Int. J. Mol. Sci. 2020, 21, 1621. [Google Scholar] [CrossRef] [PubMed]
- Panner Selvam, M.K.; Baskaran, S.; Agarwal, A. Proteomics of reproduction: Prospects and perspectives. Adv. Clin. Chem. 2019, 92, 217–243. [Google Scholar]
- Samanta, L.; Parida, R.; Dias, T.R.; Agarwal, A. The enigmatic seminal plasma: A proteomics insight from ejaculation to fertilization. Reprod. Biol. Endocrinol. 2018, 16, 41. [Google Scholar] [CrossRef]
- Duncan, M.W.; Thompson, H.S. Proteomics of semen and its constituents. Proteom. Clin. Appl. 2007, 1, 861–875. [Google Scholar] [CrossRef]
- Jodar, M.; Soler-Ventura, A.; Oliva, R.; Molecular Biology of Reproduction and Development Research Group. Semen proteomics and male infertility. J. Proteom. 2017, 162, 125–134. [Google Scholar] [CrossRef]
- Rusevski, A.; Plaseska-Karanfilska, D.; Davalieva, K. Proteomics of azoospermia: Towards the discovery of reliable markers for non-invasive diagnosis. Proteom. Clin. Appl. 2022, 30, e2200060. [Google Scholar] [CrossRef]
- Wang, F.; Yang, W.; Ouyang, S.; Yuan, S. The Vehicle Determines the Destination: The Significance of Seminal Plasma Factors for Male Fertility. Int. J. Mol. Sci. 2020, 21, 8499. [Google Scholar] [CrossRef]
- Bianchi, L.; Carnemolla, C.; Viviani, V.; Landi, C.; Pavone, V.; Luddi, A.; Piomboni, P.; Bini, L. Soluble protein fraction of human seminal plasma. J. Proteom. 2018, 174, 85–100. [Google Scholar] [CrossRef]
- Pilch, B.; Mann, M. Large-scale and high-confidence proteomic analysis of human seminal plasma. Genome Biol. 2006, 7, R40. [Google Scholar] [CrossRef] [PubMed]
- Fung, K.Y.; Glode, L.M.; Green, S.; Duncan, M.W. A comprehensive characterization of the peptide and protein constituents of human seminal fluid. Prostate 2004, 61, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Drabovich, A.P.; Jarvi, K.; Diamandis, E.P. Verification of male infertility biomarkers in seminal plasma by multiplex selected reaction monitoring assay. Mol. Cell Proteom. 2011, 10, M110.004127. [Google Scholar] [CrossRef] [PubMed]
- Drabovich, A.P.; Dimitromanolakis, A.; Saraon, P.; Soosaipillai, A.; Batruch, I.; Mullen, B.; Jarvi, K.; Diamandis, E.P. Differential diagnosis of azoospermia with proteomic biomarkers ECM1 and TEX101 quantified in seminal plasma. Sci. Transl. Med. 2013, 5, 212ra160. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, P.T.; Belardin, L.B.; Camargo, M.; Zylbersztejn, D.S.; Carvalho, V.M.; Cardozo, K.H.; Bertolla, R.P.; Cedenho, A.P. Determination of testicular function in adolescents with varicocoele—A proteomics approach. Andrology 2016, 4, 447–455. [Google Scholar] [CrossRef]
- Sharma, R.; Agarwal, A.; Mohanty, G.; Jesudasan, R.; Gopalan, B.; Willard, B.; Yadav, S.P.; Sabanegh, E. Functional proteomic analysis of seminal plasma proteins in men with various semen parameters. Reprod. Biol. Endocrinol. 2013, 11, 38. [Google Scholar] [CrossRef]
- Wang, J.; Wang, J.; Zhang, H.R.; Shi, H.J.; Ma, D.; Zhao, H.X.; Lin, B.; Li, R.S. Proteomic analysis of seminal plasma from asthenozoospermia patients reveals proteins that affect oxidative stress responses and semen quality. Asian J. Androl. 2009, 11, 484–491. [Google Scholar] [CrossRef]
- Batruch, I.; Smith, C.R.; Mullen, B.J.; Grober, E.; Lo, K.C.; Diamandis, E.P.; Jarvi, K.A. Analysis of seminal plasma from patients with non-obstructive azoospermia and identification of candidate biomarkers of male infertility. J. Proteome Res. 2012, 11, 1503–1511. [Google Scholar] [CrossRef]
- Herwig, R.; Knoll, C.; Planyavsky, M.; Pourbiabany, A.; Greilberger, J.; Bennett, K.L. Proteomic analysis of seminal plasma from infertile patients with oligoasthenoteratozoospermia due to oxidative stress and comparison with fertile volunteers. Fertil. Steril. 2013, 100, 355–366.e2. [Google Scholar] [CrossRef]
- Wu, Y.; Yuan, Y.; Chen, L.; Wang, M.; Yang, Y.; Wang, Y.; Quan, C.; Chen, D.; Chen, Y.; Huang, X.; et al. Quantitative Proteomic Analysis of Human Seminal Plasma from Normozoospermic and Asthenozoospermic Individuals. Biomed. Res. Int. 2019, 2019, 2735038. [Google Scholar] [CrossRef]
- Barrachina, F.; Jodar, M.; Delgado-Dueñas, D.; Soler-Ventura, A.; Estanyol, J.M.; Mallofré, C.; Ballescà, J.L.; Oliva, R. Stable-protein Pair Analysis as A Novel Strategy to Identify Proteomic Signatures: Application To Seminal Plasma From Infertile Patients. Mol. Cell Proteom. 2019, 18, S77–S90. [Google Scholar] [CrossRef] [PubMed]
- Saraswat, M.; Joenväärä, S.; Jain, T.; Tomar, A.K.; Sinha, A.; Singh, S.; Yadav, S.; Renkonen, R. Human Spermatozoa Quantitative Proteomic Signature Classifies Normo- and Asthenozoospermia. Mol. Cell Proteom. 2017, 16, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, W.; Zhu, P.; Wang, J.; Wang, Y.; Wang, X.; Liu, J.; Li, N.; Wang, X.; Lin, C.; et al. In-depth quantitative proteome analysis of seminal plasma from men with oligoasthenozoospermia and normozoospermia. Reprod. BioMedicine Online 2018, 37, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Correnti, S.; Preianò, M.; Murfone, P.; Fregola, A.; Bitonti, M.; Savino, R.; Terracciano, R. Revealing the Hidden Diagnostic Clues of Male Infertility from Human Seminal Plasma by Dispersive Solid Phase Extraction and MALDI-TOF MS. Int. J. Mol. Sci. 2022, 23, 10786. [Google Scholar] [CrossRef] [PubMed]
- Cadavid, J.A.P.; Alvarez, A.; Markert, U.R.; Cardona Maya, W. Differential protein expression in seminal plasma from fertile and infertile males. J. Hum. Reprod. Sci. 2014, 7, 206–211. [Google Scholar]
- Camargo, M.; Intasqui, P.; Bertolla, R.P. Understanding the seminal plasma proteome and its role in male fertility. Basic Clin. Androl. 2018, 28, 6. [Google Scholar] [CrossRef]
- Martins, A.D.; Panner Selvam, M.K.; Agarwal, A.; Alves, M.G.; Baskaran, S. Alterations in seminal plasma proteomic profile in men with primary and secondary infertility. Sci. Rep. 2020, 10, 7539. [Google Scholar] [CrossRef]
- Owen, D.H.; Katz, D.F. A review of the physical and chemical properties of human semen and the formulation of a semen simulant. J. Androl. 2005, 26, 459–469. [Google Scholar] [CrossRef]
- Veveris-Lowe, T.L.; Kruger, S.J.; Walsh, T.; Gardiner, R.A.; Clements, J.A. Seminal fluid characterization for male fertility and prostate cancer: Kallikrein-related serine proteases and whole proteome approaches. Semin. Thromb. Hemost. 2007, 33, 87–99. [Google Scholar] [CrossRef]
- de Lamirande, E. Semenogelin, the main protein of the human semen coagulum, regulates sperm function. Semin. Thromb. Hemost. 2007, 33, 60–68. [Google Scholar] [CrossRef]
- Anamthathmakula, P.; Winuthayanon, W. Mechanism of semen liquefaction and its potential for a novel non-hormonal contraception. Biol. Reprod. 2020, 103, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Drabovich, A.P.; Saraon, P.; Jarvi, K.; Diamandis, E.P. Seminal plasma as a diagnostic fluid for male reproductive system disorders. Nat. Rev. Urol. 2014, 11, 278–288. [Google Scholar] [CrossRef]
- Gonzales, G.F. Function of seminal vesicles and their role on male fertility. Asian J. Androl. 2001, 3, 251–258. [Google Scholar] [PubMed]
- Drake, R.R.; White, K.Y.; Fuller, T.W.; Igwe, E.; Clements, M.A.; Nyalwidhe, J.O.; Given, R.W.; Lance, R.S.; Semmes, O.J. Clinical collection and protein properties of expressed prostatic secretions as a source for biomarkers of prostatic disease. J. Proteome 2009, 72, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Mann, T. Secretory function of the prostate, seminal vesicle and other male accessory organs of reproduction. J. Reprod. Fertil. 1974, 37, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Verze, P.; Cai, T.; Lorenzetti, S. The role of the prostate in male fertility, health and disease. Nat. Rev. Urol. 2016, 13, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, R.; Saez, F. Epididymosomes, prostasomes, and liposomes: Their roles in mammalian male reproductive physiology. Reproduction 2013, 146, R21–R35. [Google Scholar] [CrossRef]
- Neyroud, A.S.; Chiechio, R.; Yefimova, M.; Lo Faro, M.J.; Dejucq-Rainsford, N.; Jaillard, S.; Even-Hernandez, P.; Marchi, V.; Ravel, C. Extra-cellular vesicles of the male genital tract: New actors in male fertility? Basic Clin. Androl. 2021, 31, 25. [Google Scholar] [CrossRef]
- Lin, Y.; Liang, A.; He, Y.; Li, Z.; Li, Z.; Wang, G.; Sun, F. Proteomic analysis of seminal extracellular vesicle proteins involved in asthenozoospermia by iTRAQ. Mol. Reprod. Dev. 2019, 86, 1094–1105. [Google Scholar] [CrossRef]
- Utleg, A.G.; Yi, E.C.; Xie, T.; Shannon, P.; White, J.T.; Goodlett, D.R.; Hood, L.; Lin, B. Proteomic analysis of human prostasomes. Prostate 2003, 56, 150–161. [Google Scholar] [CrossRef]
- Yang, C.; Guo, W.B.; Zhang, W.S.; Bian, J.; Yang, J.K.; Zhou, Q.Z.; Chen, M.K.; Peng, W.; Qi, T.; Wang, C.Y.; et al. Comprehensive proteomics analysis of exosomes derived from human seminal plasma. Andrology 2017, 5, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Murdica, V.; Cermisoni, G.C.; Zarovni, N.; Salonia, A.; Viganò, P.; Vago, R. Proteomic analysis reveals the negative modulator of sperm function glycodelin as over-represented in semen exosomes isolated from asthenozoospermic patients. Hum. Reprod. 2019, 34, 2314–2315. [Google Scholar] [CrossRef] [PubMed]
- Murdica, V.; Giacomini, E.; Alteri, A.; Bartolacci, A.; Cermisoni, G.C.; Zarovni, N.; Papaleo, E.; Montorsi, F.; Salonia, A.; Viganò, P.; et al. Seminal plasma of men with severe asthenozoospermia contain exosomes that affect spermatozoa motility and capacitation. Fertil. Steril. 2019, 111, 897–908. [Google Scholar] [CrossRef] [PubMed]
- Politch, J.A.; Tucker, L.; Bowman, F.P.; Anderson, D.J. Concentrations and significance of cytokines and other immunologic factors in semen of healthy fertile men. Hum. Reprod. 2007, 22, 2928–2935. [Google Scholar] [CrossRef]
- Robert, M.; Gibbs, B.F.; Jacobson, E.; Gagnon, C. Characterization of prostate-specific antigen proteolytic activity on its ma-jor physiological substrate, the sperm motility inhibitor precursor/semenogelin I. Biochemistry 1997, 36, 3811–3819. [Google Scholar] [CrossRef]
- Cupp-Sutton, K.A.; Wu, S. High-throughput quantitative top-down proteomics. Mol. Omics. 2020, 16, 91–99. [Google Scholar] [CrossRef]
- Milardi, D.; Grande, G.; Vincenzoni, F.; Messana, I.; Pontecorvi, A.; De Marinis, L.; Castagnola, M.; Marana, R. Proteomic approach in the identification of fertility pattern in seminal plasma of fertile men. Fertil. Steril. 2012, 97, 67–73.e1. [Google Scholar] [CrossRef]
- Shi, T.; Su, D.; Liu, T.; Tang, K.; Camp, D.G., 2nd; Qian, W.J.; Smith, R.D. Advancing the sensitivity of selected reaction monitoring-based targeted quantitative proteomics. Proteomics 2012, 12, 1074–1092. [Google Scholar] [CrossRef]
- Schwanhäusser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global quantification of mammalian gene expression control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef]
- Arike, L.; Peil, L. Spectral counting label-free proteomics. Methods Mol. Biol. 2014, 1156, 213–222. [Google Scholar]
- Di Palma, S.; Hennrich, M.L.; Heck, A.J.; Mohammed, S. Recent advances in peptide separation by multidimensional liquid chromatography for proteome analysis. J. Proteom. 2012, 75, 3791–3813. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, S.; Heck, A., Jr. Strong cation exchange (SCX) based analytical methods for the targeted analysis of protein post-translational modifications. Curr. Opin. Biotechnol. 2011, 22, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Rozanova, S.; Barkovits, K.; Nikolov, M.; Schmidt, C.; Urlaub, H.; Marcus, K. Quantitative Mass Spectrometry-Based Proteomics: An Overview. In Quantitative Methods in Proteomics, 2nd ed.; Marcus, K., Eisenacher, M., Sitek, B., Eds.; Humana: New York, NY, USA, 2021; Volume 2228, pp. 85–116. [Google Scholar]
- Chen, X.; Sun, Y.; Zhang, T.; Shu, L.; Roepstorff, P.; Yang, F. Quantitative Proteomics Using Isobaric Labeling: A Practical Guide. Genom. Proteom. Bioinform. 2021, 19, 689–706. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Samuel, M.; Ang, C.S.; Keerthikumar, S.; Mathivanan, S. Label-Based and Label-Free Strategies for Protein Quantitation. Methods Mol. Biol. 2017, 1549, 31–43. [Google Scholar] [PubMed]
- Gilar, M.; Olivova, P.; Daly, A.E.; Gebler, J.C. Two-dimensional separation of peptides using RP-RP-HPLC system with different pH in first and second separation dimensions. J. Sep. Sci. 2005, 28, 1694–1703. [Google Scholar] [CrossRef]
- Savino, R.; Terracciano, R. Mesopore-assisted profiling strategies in clinical proteomics for drug/target discovery. Drug Discov. Today 2012, 17, 143–152. [Google Scholar] [CrossRef]
- Diamandis, E.P. Mass spectrometry as a diagnostic and a cancer biomarker discovery tool: Opportunities and potential limitations. Mol. Cell Proteom. 2004, 3, 367–378. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, J.; Zhang, W.; Long, W.; Zhang, X.B. Proteomics technology for screening biomarkers of oligospermic candidates. Natl. J. Androl. 2007, 12, 39–42. [Google Scholar]
- Bai, J.; Sun, L.; Zhu, H.; Zhang, L.W.; Ma, J.L.; Cong, Y.L. Protein compositions in the seminal plasma are similar between severe oligospermic and healthy fertile men. Natl. J. Androl. 2008, 14, 298–302. [Google Scholar]
- Codina, M.; Estanyol, J.M.; Fidalgo, M.J.; Ballescà, J.L.; Oliva, R. Advances in sperm proteomics: Best-practise methodology and clinical potential. Expert. Rev. Proteom. 2015, 12, 255–277. [Google Scholar] [CrossRef]
- Preianò, M.; Maggisano, G.; Lombardo, N.; Montalcini, T.; Paduano, S.; Pelaia, G.; Savino, R.; Terracciano, R. Influence of storage conditions on MALDI-TOF MS profiling of gingival crevicular fluid: Implications on the role of S100A8 and S100A9 for clinical and proteomic based diagnostic investigations. Proteomics 2016, 16, 1033–1045. [Google Scholar] [CrossRef] [PubMed]
- Preianò, M.; Maggisano, G.; Murfuni, M.S.; Villella, C.; Pelaia, C.; Montalcini, T.; Lombardo, N.; Pelaia, G.; Savino, R.; Terracciano, R. An Analytical Method for Assessing Optimal Storage Conditions of Gingival Crevicular Fluid and Disclosing a Peptide Biomarker Signature of Gingivitis by MALDI-TOF MS. Proteom. Clin. Appl. 2018, 12, e1800005. [Google Scholar] [CrossRef] [PubMed]
- Preianò, M.; Savino, R.; Villella, C.; Pelaia, C.; Terracciano, R. Gingival Crevicular Fluid Peptidome Profiling in Healthy and in Periodontal Diseases. Int. J. Mol. Sci. 2020, 21, 5270. [Google Scholar] [CrossRef]
- Panner Selvam, M.K.; Agarwal, A. Sperm and Seminal Plasma Proteomics: Molecular Changes Associated with Varicocele-Mediated Male Infertility. World J. Mens Health 2020, 38, 472–483. [Google Scholar] [CrossRef] [PubMed]
- Diz, A.P.; Truebano, M.; Skibinski, D.O.F. The consequences of sample pooling in proteomics: An empirical study. Electrophoresis 2009, 30, 2967–2975. [Google Scholar] [CrossRef]
- Molinari, N.; Roche, S.; Peoc’h, K.; Tiers, L.; Séveno, M.; Hirtz, C.; Lehmann, S. Sample Pooling and Inflammation Linked to the False Selection of Biomarkers for Neurodegenerative Diseases in Top-Down Proteomics: A Pilot Study. Front. Mol. Neurosci. 2018, 11, 477. [Google Scholar] [CrossRef] [PubMed]
- Savino, R.; Paduano, S.; Preianò, M.; Terracciano, R. The proteomics big challenge for biomarkers and new drug-targets discovery. Int. J. Mol. Sci. 2012, 13, 13926–13948. [Google Scholar] [CrossRef]
- Drabovich, A.P.; Pavlou, M.P.; Batruch, I.; Diamandis, E.P. Proteomic and Mass Spectrometry Technologies for Biomarker Discovery. In Proteomic and Metabolomic Approaches to Biomarker Discovery; Issaq, H.J., Veenstra, T.D., Eds.; Academic Press (Elsevier): Waltham, MA, USA, 2013; pp. 17–37. [Google Scholar]
- Zhang, J.; Kanoatov, M.; Jarvi, K.; Gauthier-Fisher, A.; Moskovtsev, S.I.; Librach, C.; Drabovich, A.P. Germ cell-specific proteins ACRV1 and AKAP4 facilitate identification of rare spermatozoa in semen of non-obstructive azoospermia patients. MedRxiv 2022. [Google Scholar]
- Jarvi, K.A.; Schlegel, P.; Schiza, C.; Drabovich, A.; Lau, S.; Soosaipillai, A.; Korbakis, D.; Brinc, D.; Mullen, B.; Diamandis, E. Semen biomarker TEX101 predicts sperm retrieval success for men with testicular failure. F1000Research 2021, 10, 569. [Google Scholar] [CrossRef]
- Korbakis, D.; Schiza, C.; Brinc, D.; Soosaipillai, A.; Karakosta, T.D.; Légaré, C.; Sullivan, R.; Mullen, B.; Jarvi, K.; Diamandis, E.P.; et al. Preclinical evaluation of a TEX101 protein ELISA test for the differential diagnosis of male infertility. BMC Med. 2017, 15, 60. [Google Scholar] [CrossRef]
- Iles, R.; Mahmoud, N. Future Laboratory Medicine: Rapid, Efficient and Affordable Screening for Haemoglobinopathies by MALDI-ToF Mass Spectrometry. Adv. Biochem. Biotehcnol. 2018, 10, 2574–7258. [Google Scholar]
- Biroccio, A.; Urbani, A.; Massoud, R.; Di Ilio, C.; Sacchetta, P.; Bernardini, S.; Cortese, C.; Federici, G. A quantitative method for the analysis of glycated and glutathionylated hemoglobin by matrix-assisted laser desorption ionization-time of flight mass spectrometry. Anal. Biochem. 2005, 336, 279–288. [Google Scholar] [CrossRef]
- Haslam, C.; Hellicar, J.; Dunn, A.; Fuetterer, A.; Hardy, N.; Marshall, P.; Paape, R.; Pemberton, M.; Resemannand, A.; Leveridge, M. The Evolution of MALDI-TOF Mass Spectrometry toward Ultra-High-Throughput Screening: 1536-Well Format and Beyond. J. Biomol. Screen. 2016, 21, 176–186. [Google Scholar] [CrossRef]
- Vitorino, R. Digging Deep into Peptidomics Applied to Body Fluids. Proteomics 2018, 18, 1700401. [Google Scholar] [CrossRef] [PubMed]
- Maes, E.; Dyer, J.M.; McKerchar, H.J.; Deb-Choudhury, S.; Clerens, S. Protein-protein cross-linking and human health: The challenge of elucidating with mass spectrometry. Expert. Rev. Proteom. 2017, 14, 917–929. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xin, L.; Shan, B.; Chen, W.; Xie, M.; Yuen, D.; Zhang, W.; Zhang, Z.; Lajoie, G.A.; Ma, B. PEAKS DB: De novo sequencing assisted database search for sensitive and accurate peptide identification. Mol. Cell. Proteom. 2012, 11, M111.010587. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Zhang, K.; Hendrie, C.; Liang, C.; Li, M.; Doherty-Kirby, A.; Lajoie, G. PEAKS: Powerful software for peptide de novo sequencing by tandem mass spectrometry. Rapid. Commun. Mass Spectrom. 2003, 17, 2337–2342. [Google Scholar] [CrossRef] [PubMed]
- Budamgunta, H.; Olexiouk, V.; Luyten, W.; Schildermans, K.; Maes, E.; Boonen, K.; Menschaert, G.; Baggerman, G. Comprehensive Peptide Analysis of Mouse Brain Striatum Identifies Novel sORF-Encoded Polypeptides. Proteomics 2018, 18, e1700218. [Google Scholar] [CrossRef]
- Aiello, D.; Casadonte, F.; Terracciano, R.; Damiano, R.; Savino, R.; Sindona, G.; Napoli, A. Targeted proteomic approach in prostatic tissue: A panel of potential biomarkers for cancer detection. Oncoscience 2016, 3, 220–241. [Google Scholar] [CrossRef]
- Sinha, A.; Singh, V.; Yadav, S. Multi-omics and male infertility: Status, integration and future prospects. Front. Biosci. 2017, 9, 375–394. [Google Scholar]
- Xu, W.; Hu, H.; Wang, Z.; Chen, X.; Yang, F.; Zhu, Z.; Fang, P.; Dai, J.; Wang, L.; Shi, H.; et al. Proteomic characteristics of spermatozoa in normozoospermic patients with infertility. J. Proteom. 2012, 75, 5426–5436. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wu, Y.; Jin, K.; Lu, H.; Liu, F.; Guo, Y.; Yan, F.; Shi, W.; Liu, Y.; Cao, X.; et al. Differential proteomic profiling in human spermatozoa that did or did not result in pregnancy via IVF and AID. Proteom. Clin. Appl. 2013, 7, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Azpiazu, R.; Amaral, A.; Castillo, J.; Estanyol, J.M.; Guimerà, M.; Ballescà, J.L.; Balasch, J.; Oliva, R. High-throughput sperm differential proteomics suggests that epigenetic alterations contribute to failed assisted reproduction. Hum. Reprod. 2014, 29, 1225–1237. [Google Scholar] [CrossRef] [PubMed]
- McReynolds, S.; Dzieciatkowska, M.; Stevens, J.; Hansen, K.C.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Toward the identification of a subset of unexplained infertility: A sperm proteomic approach. Fertil. Steril. 2014, 102, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, G.; Liu, J.; Zhu, P.; Wang, J.; Wang, Y.; Wang, W.; Li, N.; Wang, X.; Zhang, C.; et al. iTRAQ-based analysis of sperm proteome from normozoospermic men achieving the rescue-ICSI pregnancy after the IVF failure. Clin. Proteom. 2018, 15, 27. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Proteomic Approach | SP Processing and Fractionation | Samples Used for Proteomics | Normalization of Protein Quantitation | Quantification Strategies | Instrumentation/Techniques | Study Groups | References |
|---|---|---|---|---|---|---|---|
| Bottom-up |
| 58 individual samples | Yes | Label-free method (SpC 1) | 1D-PAGE 2 ESI 3—Ion Trap/Orbitrap |
| Wang et al., 2009 [29] |
| Bottom-up |
|
| Yes | Label-free method (SpC) | ESI-Triple-Quadrupole and Ion Trap/Orbitrap |
| Drabovich et al., 2011 [25] |
| Bottom-up |
| Pooled samples for each group | Yes | Label-free methods (SpC and XIC 10) | ESI-Ion Trap/Orbitrap |
| Batruch et al., 2012 [30] |
| Bottom-up |
| 119 individual samples | Yes | Stable isotope dilution SRM 11 | ESI-Triple-Quadrupole |
| Drabovich et al., 2013 [26] |
| Bottom-up |
| Pooled samples for each group | Yes | Label-free method (SpC) | ESI-Ion Trap/Quadrupole/Orbitrap |
| Herwig et al., 2013 [31] |
| Bottom-up |
| Pooled samples for each group | Not mentioned | Label-free method (SpC) | ESI-Ion Trap |
| Sharma et al., 2013 [28] |
| Top-down |
| 16 individual samples | Yes | Label-free method (peak intensities) | SELDI-TOF 18 MS |
| Cadavid et al., 2014 [37] |
| Bottom-up |
| Pooled samples for each group | Yes | Label-free method (iBAQ 21) | ESI- Quadrupole/Orbitrap |
| Del Giudice et al., 2016 [27] |
| Bottom-up |
| 17 individual samples | Yes | Label-free method | UPLC-MS |
| Saraswat et al., 2017 [34] |
| Bottom-up |
| Pooled samples for each group | Yes | Label-based method (iTRAQ 22) | MALDI-TOF/TOF 23 MS |
| Liu et al., 2018 [35] |
| Bottom-up |
| 6 individual samples | Yes | Label-based (TMT 24) and label-free methods (iBAQ) | ESI-Ion Trap/Orbitrap |
| Wu et al., 2019 [32] |
| Bottom-up |
| Pooled samples for each group | Yes | Label-based method (TMT) | ESI-Ion Trap/Orbitrap |
| Barrachina et al., 2019 [33] |
| Top-down |
| 30 individual samples | Yes | Label-free method (peak intensities) | MALDI-TOF/TOF MS |
| Correnti et al., 2022 [36] |
| References | Study Groups | Main Findings | Candidate Biomarkers Expression Levels | Validation Assay |
|---|---|---|---|---|
| Wang et al., 2009 [29] |
| 45 proteins were ↑ 3 and 56 proteins were ↓ 4 in the AS group. Most of these proteins originated from the epididymis and prostate. ITLN1, ADH, ALAD, DJ-1 were suggested as biomarkers for oxidative stress in AS patients |
| Western Blot (DJ-1) |
| Drabovich et al., 2011 [25] |
| 18 biomarkers were identified at differential abundance between NZ, Non-obstructive AZ, and PV. Some testis-specific proteins (LDHC, TEX101, and SPAG11B) performed with absolute or nearly absolute specificities and sensitivities. |
| Stable-isotope dilution SRM 7 |
| Batruch et al., 2012 [30] |
|
|
| None |
| Drabovich et al., 2013 [26] |
| Epididymis-expressed ECM1 and testis-expressed TEX101 were differentially expressed between Obstructive AZ and Non-obstructive AZ patients and may serve as biomarkers of azoospermia |
| ELISA (ECM1) Immunohistochemistry analysis (TEX101) Immuno-SRM assay (TEX101) |
| Herwig et al., 2013 [31] |
| 24 proteins were ↑ in the OAT compared to NZ men. These proteins are mainly involved in metabolism and inflammation, defense, and stress responses | AACT and ALDR were ↑ in OAT. | None |
| Sharma et al., 2013 [28] |
| 20 proteins were differentially expressed among the 4 groups. Biological regulation is the main process affected. |
| None |
| Cadavid et al., 2014 [37] |
| Ten proteins were identified with a statistical difference between the NZ and OA patients | UBE2C, CSTA, DCD, GNAO, IDE, CERU, SLIT2 Isoform 1, UGT 1 precursor, IQGAP1 were ↑ in OA | None |
| Del Giudice et al., 2016 [27] |
|
|
| Western Blot (Cab45 and CRISP-3) |
| Saraswat et al., 2017 [34] |
| 429 proteins were differentially expressed between NZ and AS |
| None |
| Liu et al., 2018 [35] |
| 22 proteins were ↑ and 20 proteins were ↓ in the OA patients. These proteins were involved in physiological processes, including metabolism, transport, antioxidation and immune response. |
| Western Blot (PSA and LTF) |
| Wu et al., 2019 [32] |
| 29 proteins were differentially expressed between the two groups:
|
| Western Blot (KLK2, HSPA2, SORD, ANAX2) |
| Barrachina et al., 2019 [33] |
| 6 proteins were differentially expressed among the groups. These proteins are involved in sperm-oocyte binding, fertilization and immune response. | CRISP1, NPC2, SPINT3, and ECM1 were ↓ in OZ and AZ IGHG2 was ↑ in NZ ANPEP was ↓ in AS | Western Blot (ECM1) |
| Correnti et al., 2022 [36] |
| 7 peaks were statistically different between NZ and infertile men. | All the peaks were ↓ in infertile men compared to NZ:
| None |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Preianò, M.; Correnti, S.; Butt, T.A.; Viglietto, G.; Savino, R.; Terracciano, R. Mass Spectrometry-Based Untargeted Approaches to Reveal Diagnostic Signatures of Male Infertility in Seminal Plasma: A New Laboratory Perspective for the Clinical Management of Infertility? Int. J. Mol. Sci. 2023, 24, 4429. https://doi.org/10.3390/ijms24054429
Preianò M, Correnti S, Butt TA, Viglietto G, Savino R, Terracciano R. Mass Spectrometry-Based Untargeted Approaches to Reveal Diagnostic Signatures of Male Infertility in Seminal Plasma: A New Laboratory Perspective for the Clinical Management of Infertility? International Journal of Molecular Sciences. 2023; 24(5):4429. https://doi.org/10.3390/ijms24054429
Chicago/Turabian StylePreianò, Mariaimmacolata, Serena Correnti, Tahreem Arshad Butt, Giuseppe Viglietto, Rocco Savino, and Rosa Terracciano. 2023. "Mass Spectrometry-Based Untargeted Approaches to Reveal Diagnostic Signatures of Male Infertility in Seminal Plasma: A New Laboratory Perspective for the Clinical Management of Infertility?" International Journal of Molecular Sciences 24, no. 5: 4429. https://doi.org/10.3390/ijms24054429
APA StylePreianò, M., Correnti, S., Butt, T. A., Viglietto, G., Savino, R., & Terracciano, R. (2023). Mass Spectrometry-Based Untargeted Approaches to Reveal Diagnostic Signatures of Male Infertility in Seminal Plasma: A New Laboratory Perspective for the Clinical Management of Infertility? International Journal of Molecular Sciences, 24(5), 4429. https://doi.org/10.3390/ijms24054429