

Microvascular Environment Influences Brain Microvascular Heterogeneity: Relative Roles of Astrocytes and Oligodendrocytes for the EPCR Expression in the Brain Endothelium


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
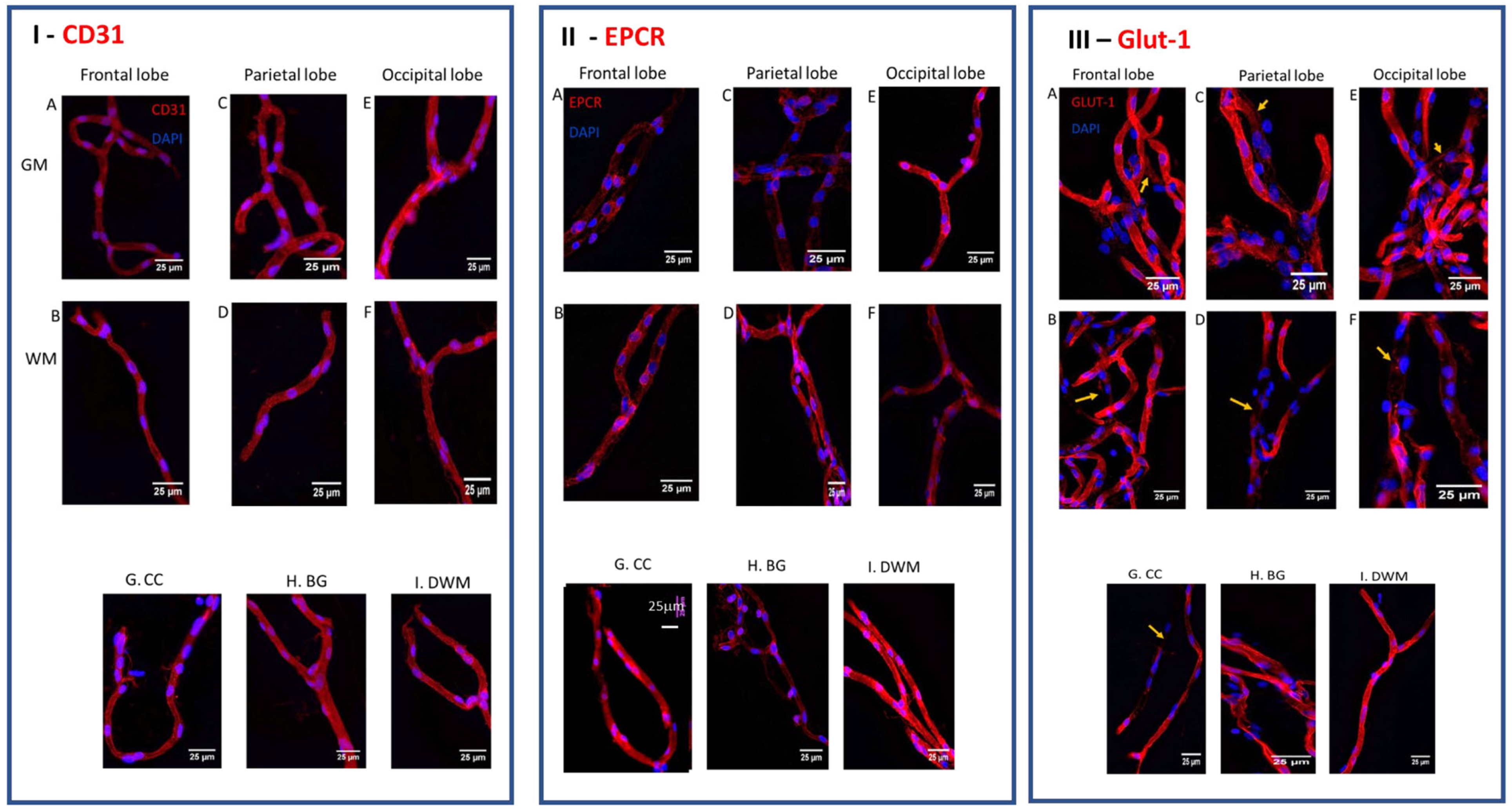
2.1. Heterogeneity in the Expression of EPCR on Cerebral Microvessels
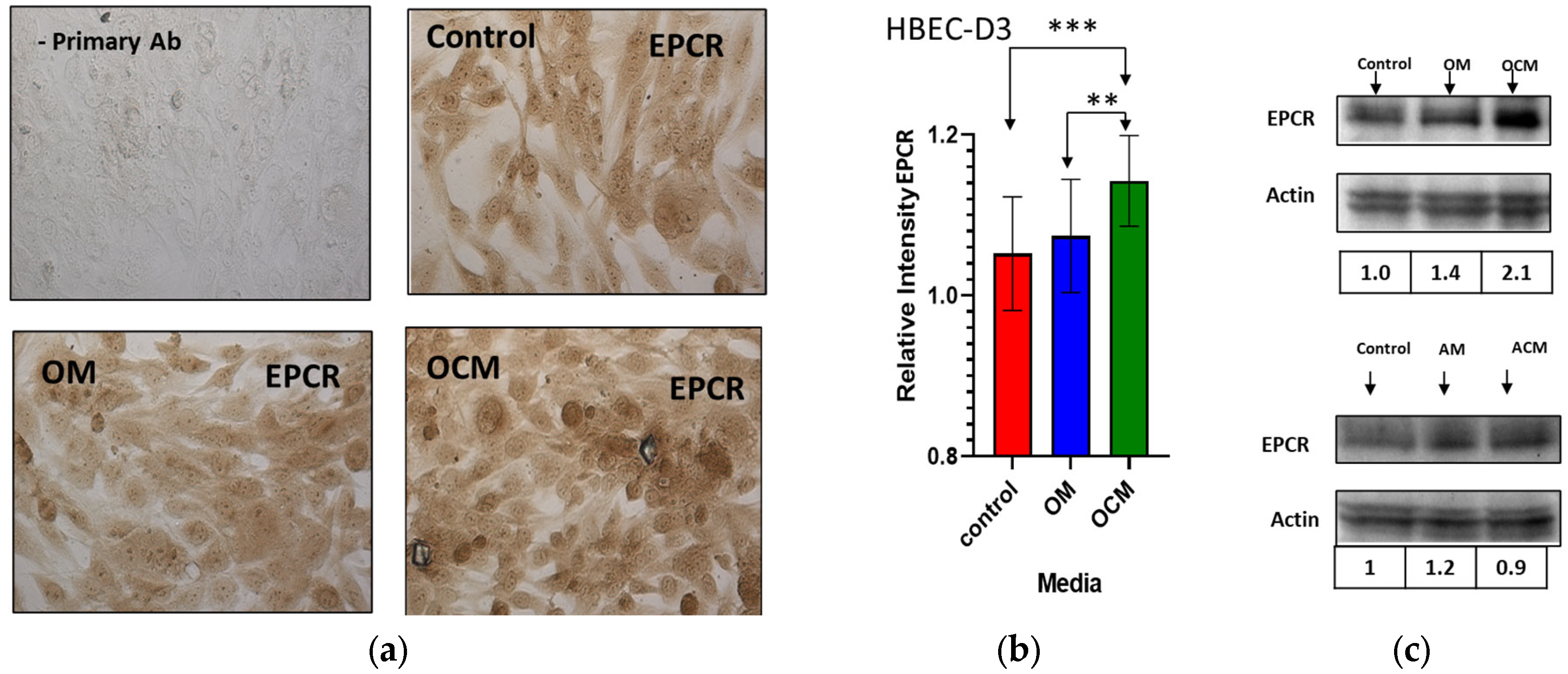
2.2. The Effect of Conditioned Media Derived from Oli-Neu and Astrocyte Cultures on the Expression of EPCR on Endothelial Cells
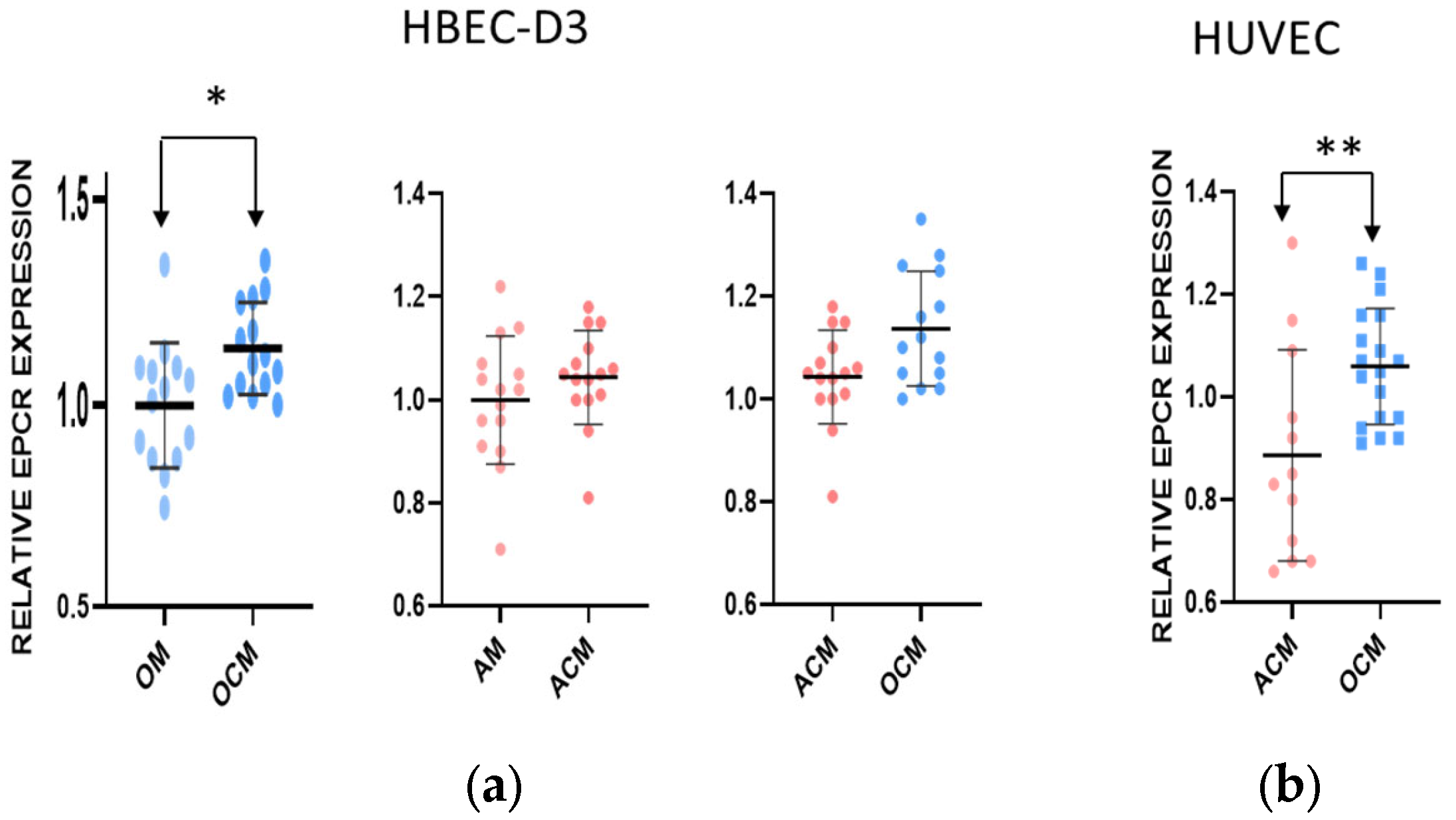
2.3. The Effect of OCM and ACM Media on the Expression of EPCR on Endothelial and Macrovascular Cells
2.4. Heat Inactivation of OCM Eliminated Its Effect on EPCR Expression
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Cultures and Conditioned Media Collection
4.3. Brain Vessel Isolations
4.4. Immunohistochemistry and Immunofluorescence
4.4.1. Microvessels
4.4.2. In Vitro Cell Cultures
4.5. Quantification by ELISA
4.6. Western Blotting
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nyul-Toth, A.; Suciu, M.; Molnar, J.; Fazakas, C.; Hasko, J.; Herman, H.; Farkas, A.E.; Kaszaki, J.; Hermenean, A.; Wilhelm, I.; et al. Differences in the molecular structure of the blood-brain barrier in the cerebral cortex and white matter: An in silico, in vitro, and ex vivo study. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H1702–H1714. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed]
- Dorovini-Zis, K.; Schmidt, K.; Huynh, H.; Fu, W.; Whitten, R.O.; Milner, D.; Kamiza, S.; Molyneux, M.; Taylor, T.E. The neuropathology of fatal cerebral malaria in malawian children. Am. J. Pathol. 2011, 178, 2146–2158. [Google Scholar] [CrossRef]
- Villabona-Rueda, A.; Erice, C.; Pardo, C.A.; Stins, M.F. The Evolving Concept of the Blood Brain Barrier (BBB): From a Single Static Barrier to a Heterogeneous and Dynamic Relay Center. Front. Cell. Neurosci. 2019, 13, 405. [Google Scholar] [CrossRef]
- MacPherson, G.G.; Warrell, M.J.; White, N.J.; Looareesuwan, S.; Warrell, D.A. Human cerebral malaria. A quantitative ultrastructural analysis of parasitized erythrocyte sequestration. Am. J. Pathol. 1985, 119, 385–401. [Google Scholar]
- Warrell, D.A. Cerebral malaria: Clinical features, pathophysiology and treatment. Ann. Trop. Med. Parasitol. 1997, 91, 875–884. [Google Scholar] [PubMed]
- Ponsford, M.J.; Medana, I.M.; Prapansilp, P.; Hien, T.T.; Lee, S.J.; Dondorp, A.M.; Esiri, M.M.; Day, N.P.; White, N.J.; Turner, G.D. Sequestration and microvascular congestion are associated with coma in human cerebral malaria. J. Infect. Dis. 2012, 205, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Moxon, C.A.; Heyderman, R.S.; Wassmer, S.C. Dysregulation of coagulation in cerebral malaria. Mol. Biochem. Parasitol. 2009, 166, 99–108. [Google Scholar] [CrossRef]
- Avril, M.; Bernabeu, M.; Benjamin, M.; Brazier, A.J.; Smith, J.D. Interaction between Endothelial Protein C Receptor and Intercellular Adhesion Molecule 1 to Mediate Binding of Plasmodium falciparum-Infected Erythrocytes to Endothelial Cells. mBio 2016, 7, e00615-16. [Google Scholar] [CrossRef]
- Brown, H.C.; Chau, T.T.; Mai, N.T.; Day, N.P.; Sinh, D.X.; White, N.J.; Hien, T.T.; Farrar, J.; Turner, G.D. Blood-brain barrier function in cerebral malaria and CNS infections in Vietnam. Neurology 2000, 55, 104–111. [Google Scholar] [CrossRef]
- Turner, L.; Lavstsen, T.; Berger, S.S.; Wang, C.W.; Petersen, J.E.; Avril, M.; Brazier, A.J.; Freeth, J.; Jespersen, J.S.; Nielsen, M.A.; et al. Severe malaria is associated with parasite binding to endothelial protein C receptor. Nature 2013, 498, 502–505. [Google Scholar] [CrossRef] [PubMed]
- Mohan Rao, L.V.; Esmon, C.T.; Pendurthi, U.R. Endothelial cell protein C receptor: A multiliganded and multifunctional receptor. Blood 2014, 124, 1553–1562. [Google Scholar] [CrossRef]
- Pendurthi, U.R.; Rao, L.V.M. Endothelial cell protein C receptor-dependent signaling. Curr. Opin. Hematol. 2018, 25, 219–226. [Google Scholar] [CrossRef]
- Thanabalasundaram, G.; Schneidewind, J.; Pieper, C.; Galla, H.J. The impact of pericytes on the blood-brain barrier integrity depends critically on the pericyte differentiation stage. Int. J. Biochem. Cell. Biol. 2011, 43, 1284–1293. [Google Scholar] [CrossRef] [PubMed]
- Cavaglia, M.; Dombrowski, S.M.; Drazba, J.; Vasanji, A.; Bokesch, P.M.; Janigro, D. Regional variation in brain capillary density and vascular response to ischemia. Brain Res. 2001, 910, 81–93. [Google Scholar] [CrossRef]
- Nonaka, H.; Akima, M.; Hatori, T.; Nagayama, T.; Zhang, Z.; Ihara, F. Microvasculature of the human cerebral white matter: Arteries of the deep white matter. Neuropathology 2003, 23, 111–118. [Google Scholar] [CrossRef]
- Chi, J.T.; Chang, H.Y.; Haraldsen, G.; Jahnsen, F.L.; Troyanskaya, O.G.; Chang, D.S.; Wang, Z.; Rockson, S.G.; van de Rijn, M.; Botstein, D.; et al. Endothelial cell diversity revealed by global expression profiling. Proc. Natl. Acad. Sci. USA 2003, 100, 10623–10628. [Google Scholar] [CrossRef]
- Ge, S.; Pachter, J.S. Isolation and culture of microvascular endothelial cells from murine spinal cord. J. Neuroimmunol. 2006, 177, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Saubamea, B.; Cochois-Guegan, V.; Cisternino, S.; Scherrmann, J.M. Heterogeneity in the rat brain vasculature revealed by quantitative confocal analysis of endothelial barrier antigen and P-glycoprotein expression. J. Cereb. Blood Flow. Metab. 2012, 32, 81–92. [Google Scholar] [CrossRef]
- Ball, H.J.; McParland, B.; Driussi, C.; Hunt, N.H. Isolating vessels from the mouse brain for gene expression analysis using laser capture microdissection. Brain Res. Brain Res. Protoc. 2002, 9, 206–213. [Google Scholar] [CrossRef]
- Fonta, C.; Barone, P.; Rodriguez Martinez, L.; Negyessy, L. Rediscovering TNAP in the Brain: A Major Role in Regulating the Function and Development of the Cerebral Cortex. Subcell. Biochem. 2015, 76, 85–106. [Google Scholar]
- Calhau, C.; Martel, F.; Pinheiro-Silva, S.; Pinheiro, H.; Soares-da-Silva, P.; Hipolito-Reis, C.; Azevedo, I. Modulation of insulin transport in rat brain microvessel endothelial cells by an ecto-phosphatase activity. J. Cell. Biochem. 2002, 84, 389–400. [Google Scholar] [CrossRef]
- Laszik, Z.; Mitro, A.; Taylor, F.B., Jr.; Ferrell, G.; Esmon, C.T. Human protein C receptor is present primarily on endothelium of large blood vessels: Implications for the control of the protein C pathway. Circulation 1997, 96, 3633–3640. [Google Scholar] [CrossRef]
- Hase, Y.; Ding, R.; Harrison, G.; Hawthorne, E.; King, A.; Gettings, S.; Platten, C.; Stevenson, W.; Craggs, L.J.L.; Kalaria, R.N. White matter capillaries in vascular and neurodegenerative dementias. Acta Neuropathol. Commun. 2019, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Moxon, C.A.; Wassmer, S.C.; Milner, D.A., Jr.; Chisala, N.V.; Taylor, T.E.; Seydel, K.B.; Molyneux, M.E.; Faragher, B.; Esmon, C.T.; Downey, C.; et al. Loss of endothelial protein C receptors links coagulation and inflammation to parasite sequestration in cerebral malaria in African children. Blood 2013, 122, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Alber, J.; Alladi, S.; Bae, H.J.; Barton, D.A.; Beckett, L.A.; Bell, J.M.; Berman, S.E.; Biessels, G.J.; Black, S.E.; Bos, I.; et al. White matter hyperintensities in vascular contributions to cognitive impairment and dementia (VCID): Knowledge gaps and opportunities. Alzheimers Dement. 2019, 5, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Wang, D.; Lan, L.; Fan, Y. Multiple Factors Involved in the Pathogenesis of White Matter Lesions. Biomed. Res. Int. 2017, 2017, 9372050. [Google Scholar] [CrossRef]
- Weksler, B.B.; Subileau, E.A.; Perriere, N.; Charneau, P.; Holloway, K.; Leveque, M.; Tricoire-Leignel, H.; Nicotra, A.; Bourdoulous, S.; Turowski, P.; et al. Blood-brain barrier-specific properties of a human adult brain endothelial cell line. FASEB J. 2005, 19, 1872–1874. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Kramer, E.; Grzenkowski, M.; Tang, K.; Blakemore, W.; Aguzzi, A.; Khazaie, K.; Chlichlia, K.; von Blankenfeld, G.; Kettenmann, H.; et al. Lines of murine oligodendroglial precursor cells immortalized by an activated neu tyrosine kinase show distinct degrees of interaction with axons in vitro and in vivo. Eur. J. Neurosci. 1995, 7, 1245–1265. [Google Scholar] [CrossRef]
- Weksler, B.; Romero, I.A.; Couraud, P.O. The hCMEC/D3 cell line as a model of the human blood brain barrier. Fluids Barriers CNS 2013, 10, 16. [Google Scholar] [CrossRef]
- Kramer-Albers, E.M.; Bretz, N.; Tenzer, S.; Winterstein, C.; Mobius, W.; Berger, H.; Nave, K.A.; Schild, H.; Trotter, J. Oligodendrocytes secrete exosomes containing major myelin and stress-protective proteins: Trophic support for axons? Proteom. Clin. Appl. 2007, 1, 1446–1461. [Google Scholar] [CrossRef]
- Stins, M.F.; Prasadarao, N.V.; Zhou, J.; Arditi, M.; Kim, K.S. Bovine brain microvascular endothelial cells transfected with SV40-large T antigen: Development of an immortalized cell line to study pathophysiology of CNS disease. In Vitro Cell. Dev. Biol. Anim. 1997, 33, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Stins, M.F.; Pearce, D.; Choi, H.; Di Cello, F.; Pardo, C.A.; Kim, K.S. CD4 and chemokine receptors on human brain microvascular endothelial cells, implications for human immunodeficiency virus type 1 pathogenesis. Endothelium 2004, 11, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.K.; Sullivan, D.J.; Stins, M.F. Plasmodium falciparum-infected erythrocytes increase intercellular adhesion molecule 1 expression on brain endothelium through NF-κB. Infect. Immun. 2006, 74, 3262–3270. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thakar, M.; Noumbissi, M.E.; Stins, M.F. Microvascular Environment Influences Brain Microvascular Heterogeneity: Relative Roles of Astrocytes and Oligodendrocytes for the EPCR Expression in the Brain Endothelium. Int. J. Mol. Sci. 2023, 24, 6908. https://doi.org/10.3390/ijms24086908
Thakar M, Noumbissi ME, Stins MF. Microvascular Environment Influences Brain Microvascular Heterogeneity: Relative Roles of Astrocytes and Oligodendrocytes for the EPCR Expression in the Brain Endothelium. International Journal of Molecular Sciences. 2023; 24(8):6908. https://doi.org/10.3390/ijms24086908
Chicago/Turabian StyleThakar, Manjusha, Midrelle E. Noumbissi, and Monique F. Stins. 2023. "Microvascular Environment Influences Brain Microvascular Heterogeneity: Relative Roles of Astrocytes and Oligodendrocytes for the EPCR Expression in the Brain Endothelium" International Journal of Molecular Sciences 24, no. 8: 6908. https://doi.org/10.3390/ijms24086908
APA StyleThakar, M., Noumbissi, M. E., & Stins, M. F. (2023). Microvascular Environment Influences Brain Microvascular Heterogeneity: Relative Roles of Astrocytes and Oligodendrocytes for the EPCR Expression in the Brain Endothelium. International Journal of Molecular Sciences, 24(8), 6908. https://doi.org/10.3390/ijms24086908