

Diacylglycerol Activates the Drosophila Light Sensitive Channel TRPL Expressed in HEK Cells
 and
and
Abstract
1. Introduction
2. Results
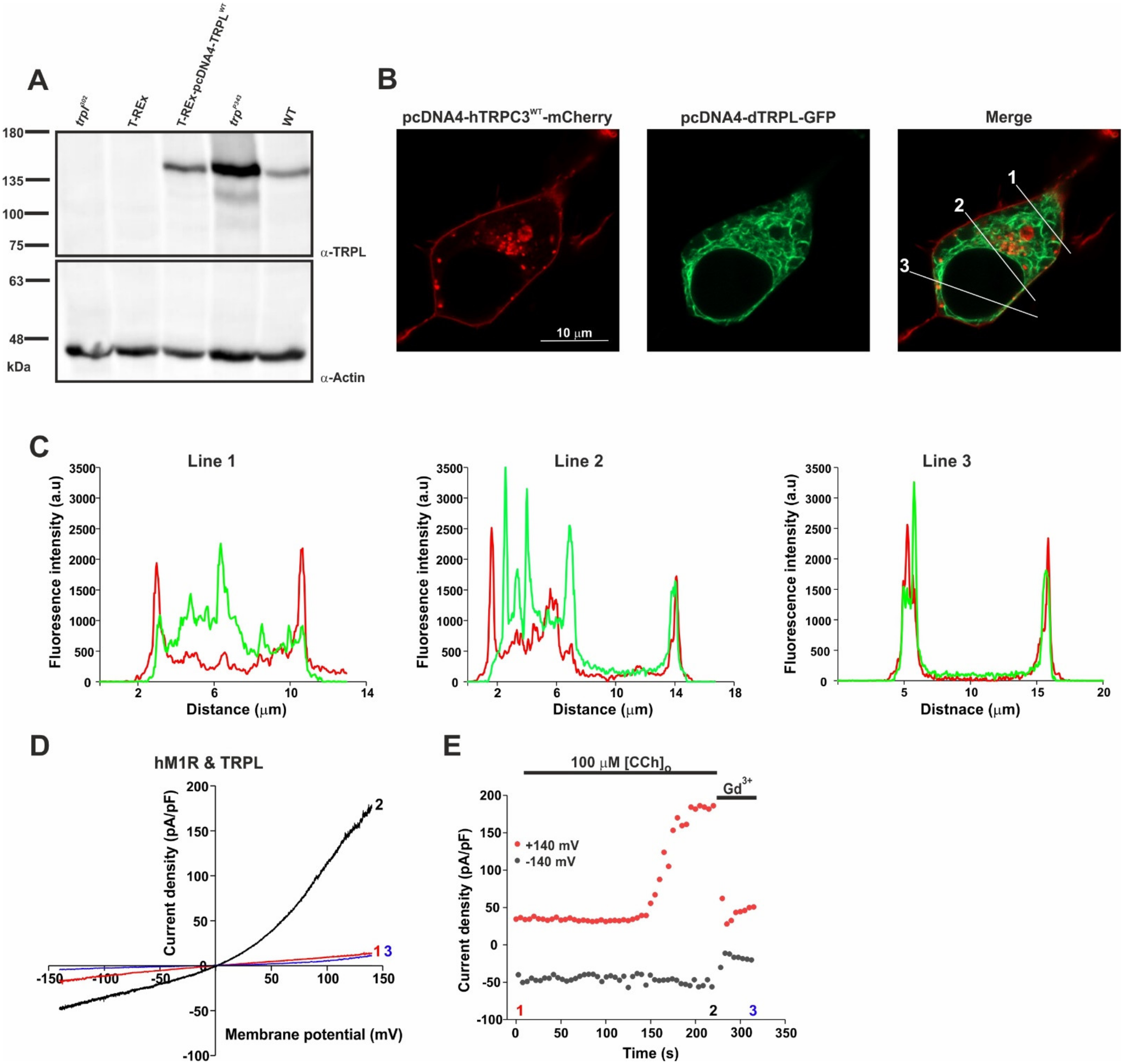
2.1. Heterologous Expression of the TRPL Channel in HEK Cells Resulted in Localization on the Surface Membrane and in Robust Functional Currents via an Endogenous Cascade
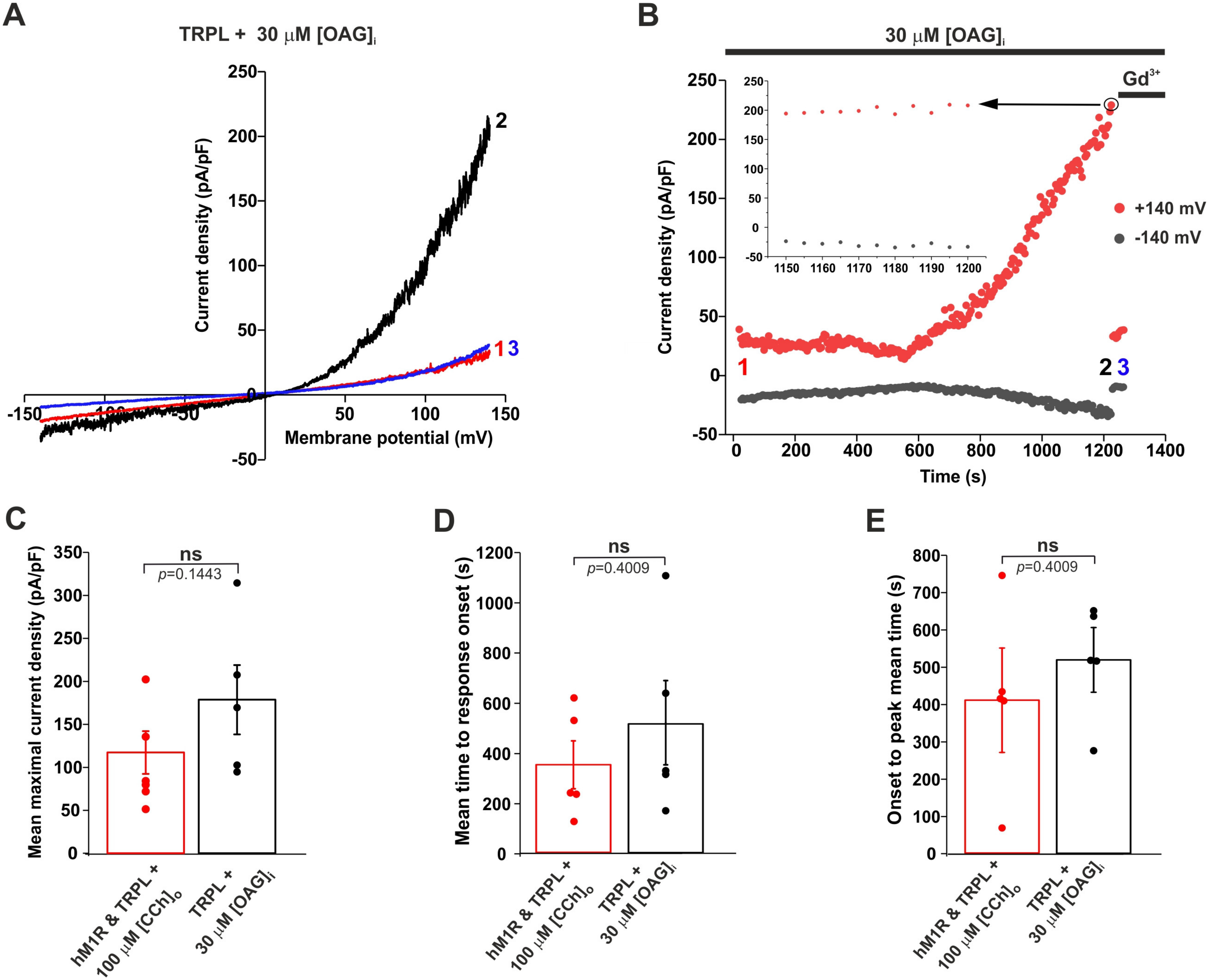
2.2. Application of a DAG Analogue to the Intracellular Solution of HEK Cells Robustly Activated the Expressed TRPL Channel
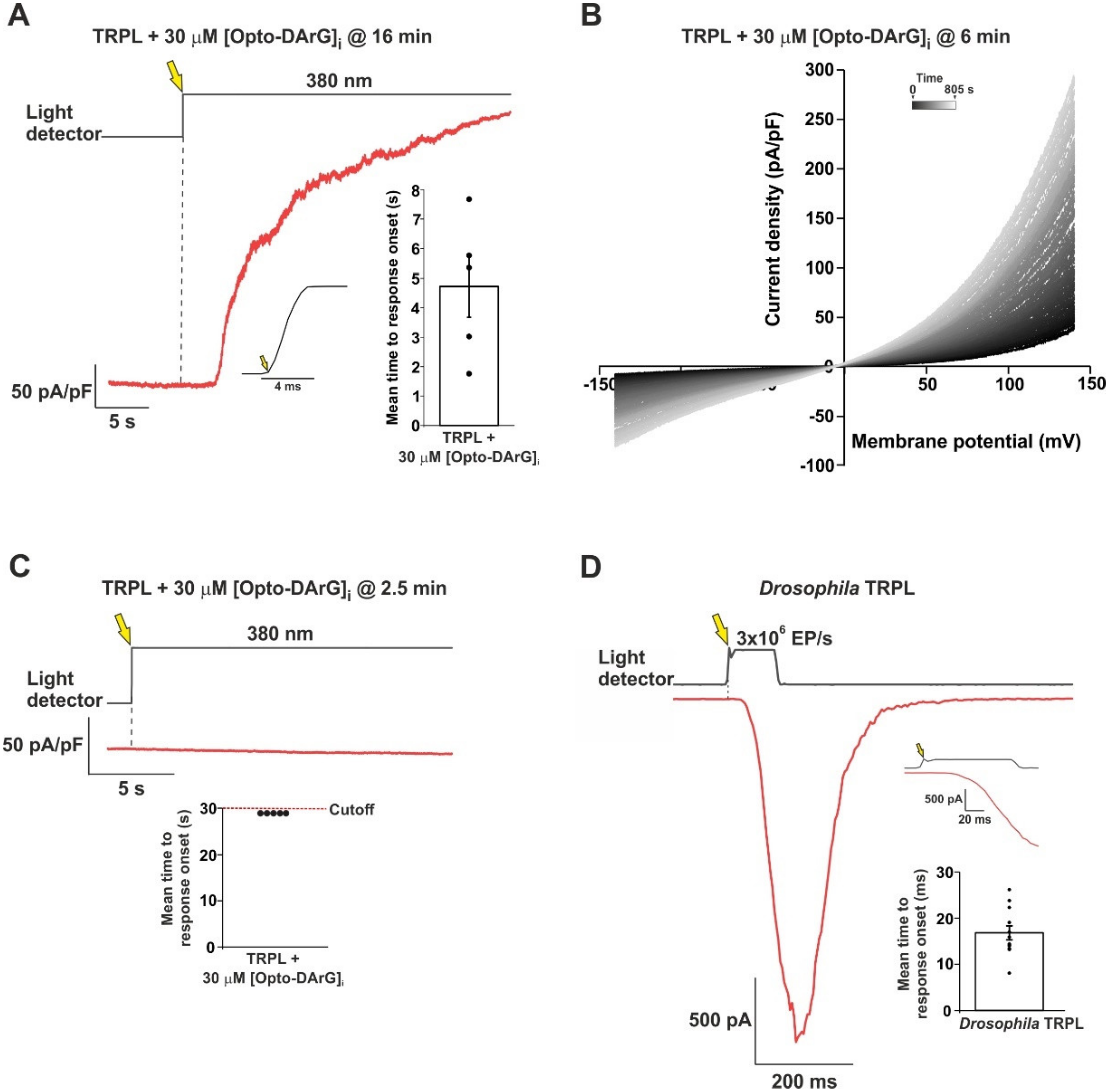
2.3. An Optically Activated DAG Analogue Accelerated the Kinetics of TRPL Channel Activation
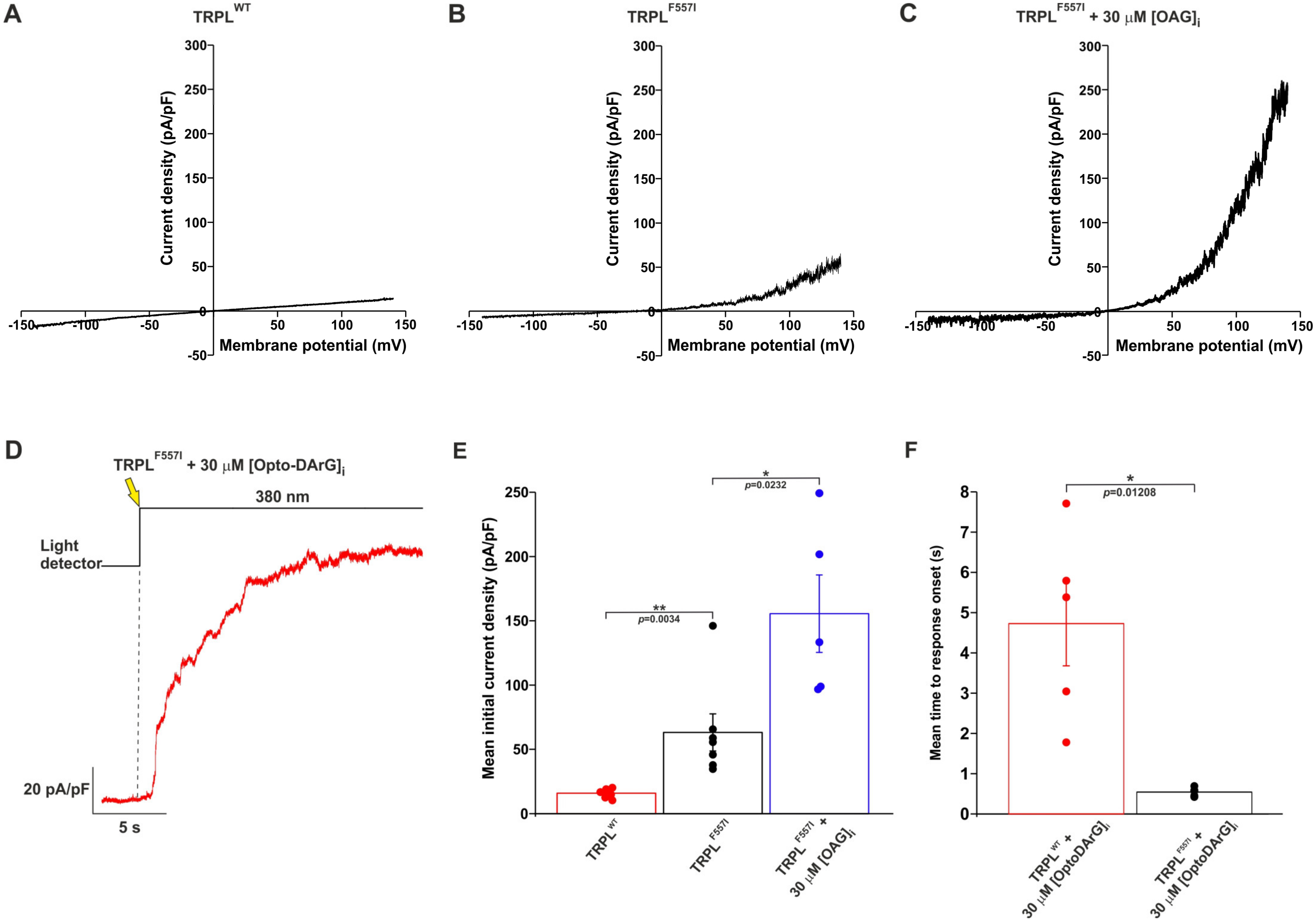
2.4. A Mutation in the TRPL Pore-Region Causing Constitutive Channel Activity Enhanced and Accelerated the OAG-Induced Activation of TRPL
3. Discussion
4. Materials and Methods
4.1. Fly Stocks
4.2. Cell Culture
4.3. Plasmids Used in This Study: See Table 1
4.4. Antibodies
4.5. Western Blot Analyses
4.6. Confocal Imaging
4.7. Electrophysiology
4.8. Solutions
4.9. Pharmacology
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Minke, B.; Pak, W.L. The light-activated TRP channel: The founding member of the TRP channel superfamily. J. Neurogenet. 2022, 36, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Cook, B.; Bar, Y.M.; Cohen-Ben, A.H.; Goldstein, R.E.; Paroush, Z.; Selinger, Z.; Minke, B. Phospholipase C and termination of G-protein-mediated signalling in vivo. Nat. Cell Biol. 2000, 2, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Hardie, R.C.; Raghu, P. Visual transduction in Drosophila. Nature 2001, 413, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Minke, B.; Cook, B. TRP channel proteins and signal transduction. Physiol. Rev. 2002, 82, 429–472. [Google Scholar] [CrossRef] [PubMed]
- Montell, C. Molecular genetics of Drosophila vision. Bioessays 1989, 11, 43–48. [Google Scholar] [CrossRef]
- Katz, B.; Minke, B. The Drosophila light-activated TRP and TRPL channels—Targets of the phosphoinositide signaling cascade. Prog. Retin. Eye Res. 2018, 66, 200–219. [Google Scholar] [CrossRef]
- Voolstra, O.; Huber, A. Ca2+ Signaling in Drosophila Photoreceptor Cells. Adv. Exp. Med. Biol. 2020, 1131, 857–879. [Google Scholar] [CrossRef]
- Masai, I.; Okazaki, A.; Hosoya, T.; Hotta, Y. Drosophila retinal degeneration A gene encodes an eye-specific diacylglycerol kinase with cysteine-rich zinc-finger motifs and ankyrin repeats. Proc. Natl. Acad. Sci. USA 1993, 90, 11157–11161. [Google Scholar] [CrossRef]
- Benzer, S. Behavioral mutants of Drosophila isolated by countercurrent distribution. Proc. Natl. Acad. Sci. USA 1967, 58, 1112–1119. [Google Scholar] [CrossRef]
- Raghu, P.; Usher, K.; Jonas, S.; Chyb, S.; Polyanovsky, A.; Hardie, R.C. Constitutive activity of the light-sensitive channels TRP and TRPL in the Drosophila diacylglycerol kinase mutant, rdgA. Neuron 2000, 26, 169–179. [Google Scholar] [CrossRef]
- Hardie, R.C. TRP channels in Drosophila photoreceptors: The lipid connection. Cell Calcium 2003, 33, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Estacion, M.; Sinkins, W.G.; Schilling, W.P. Regulation of Drosophila transient receptor potential-like (TrpL) channels by phospholipase C-dependent mechanisms. J. Physiol. 2001, 530, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Gillo, B.; Chorna, I.; Cohen, H.; Cook, B.; Manistersky, I.; Chorev, M.; Arnon, A.; Pollock, J.A.; Selinger, Z.; Minke, B. Coexpression of Drosophila TRP and TRP-like proteins in Xenopus oocytes reconstitutes capacitative Ca2+ entry. Proc. Natl. Acad. Sci. USA 1996, 93, 14146–14151. [Google Scholar] [CrossRef] [PubMed]
- Vaca, L.; Sinkins, W.G.; Hu, Y.; Kunze, D.L.; Schilling, W.P. Activation of recombinant trp by thapsigargin in Sf9 insect cells. Am. J. Physiol. 1994, 267, C1501–C1505. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Z.S.; Li, H.S.; Guggino, W.B.; Montell, C. Coassembly of TRP and TRPL produces a distinct store-operated conductance. Cell 1997, 89, 1155–1164. [Google Scholar] [CrossRef]
- Hambrecht, J.; Zimmer, S.; Flockerzi, V.; Cavalie, A. Single-channel currents through transient-receptor-potential-like (TRPL) channels. Pflug. Arch. 2000, 440, 418–426. [Google Scholar] [CrossRef]
- Hardie, R.C.; Reuss, H.; Lansdell, S.J.; Millar, N.S. Functional equivalence of native light-sensitive channels in the Drosophila trp 301 mutant and TRPL cation channels expressed in a stably transfected Drosophila cell line. Cell Calcium 1997, 21, 431–440. [Google Scholar] [CrossRef]
- Hu, Y.; Vaca, L.; Zhu, X.; Birnbaumer, L.; Kunze, D.L.; Schilling, W.P. Appearance of a novel Ca2+ influx pathway in Sf9 insect cells following expression of the transient receptor potential-like (trpl) protein of Drosophila. Biochem. Biophys. Res. Commun. 1994, 201, 1050–1056. [Google Scholar] [CrossRef]
- Zimmer, S.; Trost, C.; Cavalie, A.; Philipp, S.; Flockerzi, V. The trpl protein is a Ca2+-calmodulin activated non-selective cation (CAN) channel. Naunyn-Schmiedebergs Arch. Pharmacol. 1997, 355, 238. [Google Scholar]
- Lev, S.; Katz, B.; Tzarfaty, V.; Minke, B. Signal-dependent hydrolysis of phosphatidylinositol 4,5-bisphosphate without activation of phospholipase C: Implications on gating of Drosophila TRPL (transient receptor potential-like) channel. J. Biol. Chem. 2012, 287, 1436–1447. [Google Scholar] [CrossRef]
- Parnas, M.; Katz, B.; Minke, B. Open channel block by Ca2+ underlies the voltage dependence of Drosophila TRPL channel. J. Gen. Physiol 2007, 129, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Parnas, M.; Katz, B.; Lev, S.; Tzarfaty, V.; Dadon, D.; Gordon-Shaag, A.; Metzner, H.; Yaka, R.; Minke, B. Membrane lipid modulations remove divalent open channel block from TRP-like and NMDA channels. J. Neurosci. 2009, 29, 2371–2383. [Google Scholar] [CrossRef] [PubMed]
- Delgado, R.; Bacigalupo, J. Unitary recordings of TRP and TRPL channels from isolated Drosophila retinal photoreceptor rhabdomeres: Activation by light and lipids. J. Neurophysiol. 2009, 101, 2372–2379. [Google Scholar] [CrossRef] [PubMed]
- Delgado, R.; Muñoz, Y.; Peña-Cortés, H.; Giavalisco, P.; Bacigalupo, J. Diacylglycerol activates the light-dependent channel TRP in the photosensitive microvilli of Drosophila melanogaster photoreceptors. J. Neurosci. 2014, 34, 6679–6686. [Google Scholar] [CrossRef] [PubMed]
- Hardie, R.C. Phototransduction mechanismsin Drosophila microvillar photoreceptors. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2012, 1, 113–237. [Google Scholar] [CrossRef]
- Merino, E.; Ribagorda, M. Control over molecular motion using the cis-trans photoisomerization of the azo group. Beilstein J. Org. Chem. 2012, 8, 1071–1090. [Google Scholar] [CrossRef]
- Lichtenegger, M.; Tiapko, O.; Svobodova, B.; Stockner, T.; Glasnov, T.N.; Schreibmayer, W.; Platzer, D.; de la Cruz, G.G.; Krenn, S.; Schober, R.; et al. An optically controlled probe identifies lipid-gating fenestrations within the TRPC3 channel. Nat. Chem. Biol. 2018, 14, 396–404. [Google Scholar] [CrossRef]
- Schindl, R.; Fritsch, R.; Jardin, I.; Frischauf, I.; Kahr, H.; Muik, M.; Riedl, M.C.; Groschner, K.; Romanin, C. Canonical transient receptor potential (TRPC) 1 acts as a negative regulator for vanilloid TRPV6-mediated Ca2+ influx. J. Biol. Chem. 2012, 287, 35612–35620. [Google Scholar] [CrossRef]
- Katz, B.; Zaguri, R.; Edvardson, S.; Maayan, C.; Elpeleg, O.; Lev, S.; Davidson, E.; Peters, M.; Kfir-Erenfeld, S.; Berger, E.; et al. Nociception and pain in humans lacking functional TRPV1 channel. J. Clin. Investig. 2022, 133, e153558. [Google Scholar] [CrossRef]
- Hong, Y.S.; Park, S.; Geng, C.; Baek, K.; Bowman, J.D.; Yoon, J.; Pak, W.L. Single amino acid change in the fifth transmembrane segment of the TRP Ca2+ channel causes massive degeneration of photoreceptors. J. Biol. Chem. 2002, 277, 33884–33889. [Google Scholar] [CrossRef]
- Yoon, J.; Cohen Ben-Ami, H.; Hong, Y.S.; Park, S.; Strong, L.L.R.; Bowman, J.; Geng, C.; Baek, K.; Minke, B.; Pak, W.L. Novel mechanism of massive photoreceptor degeneration caused by mutations in the trp gene of Drosophila. J. Neurosci. 2000, 20, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Minke, B. The TRP calcium channel and retinal degeneration. Adv. Exp. Med. Biol. 2002, 514, 601–622. [Google Scholar] [PubMed]
- Wang, T.; Xu, H.; Oberwinkler, J.; Gu, Y.; Hardie, R.C.; Montell, C. Light activation, adaptation, and cell survival functions of the Na+/Ca2+ exchanger CalX. Neuron 2005, 45, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.M.; Bull, A.; Kelly, L.E. Identification of a Drosophila gene encoding a calmodulin- binding protein with homology to the trp phototransduction gene. Neuron 1992, 8, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Choi, W.; Sun, W.; Du, J.; Lü, W. Structure of the human lipid-gated cation channel TRPC3. eLife 2018, 7, e36852. [Google Scholar] [CrossRef]
- Liu, H.; Lin, W.Y.; Leibow, S.R.; Morateck, A.J.; Ahuja, M.; Muallem, S. TRPC3 channel gating by lipids requires localization at the ER/PM junctions defined by STIM1. J. Cell Biol. 2022, 221, e202107120. [Google Scholar] [CrossRef] [PubMed]
- Erkan-Candag, H.; Clarke, A.; Tiapko, O.; Gsell, M.A.; Stockner, T.; Groschner, K. Diacylglycerols interact with the L2 lipidation site in TRPC3 to induce a sensitized channel state. EMBO Rep. 2022, 23, e54276. [Google Scholar] [CrossRef]
- Huang, J.; Liu, C.H.; Hughes, S.A.; Postma, M.; Schwiening, C.J.; Hardie, R.C. Activation of TRP channels by protons and phosphoinositide depletion in Drosophila photoreceptors. Curr. Biol. 2010, 20, 189–197. [Google Scholar] [CrossRef]
- Hardie, R.C.; Franze, K. Photomechanical responses in Drosophila photoreceptors. Science 2012, 338, 260–263. [Google Scholar] [CrossRef]
- Scott, K.; Sun, Y.; Beckingham, K.; Zuker, C.S. Calmodulin regulation of Drosophila light-activated channels and receptor function mediates termination of the light response in vivo. Cell 1997, 91, 375–383. [Google Scholar] [CrossRef]
- Niemeyer, B.A.; Suzuki, E.; Scott, K.; Jalink, K.; Zuker, C.S. The Drosophila light-activated conductance is composed of the two channels TRP and TRPL. Cell 1996, 85, 651–659. [Google Scholar] [CrossRef]
- Studer, M.; McNaughton, P.A. Modulation of single-channel properties of TRPV1 by phosphorylation. J. Physiol. 2010, 588, 3743–3756. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yuan, H.; Yao, X.; Chen, S. Endogenous ion channels expressed in human embryonic kidney (HEK-293) cells. Pflug. Arch. 2022, 474, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Chyb, S.; Raghu, P.; Hardie, R.C. Polyunsaturated fatty acids activate the Drosophila light-sensitive channels TRP and TRPL. Nature 1999, 397, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, T.; Obukhov, A.G.; Schaefer, M.; Harteneck, C.; Gudermann, T.; Schultz, G. Direct activation of human TRPC6 and TRPC3 channels by diacylglycerol. Nature 1999, 397, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Acharya, J.K.; Jalink, K.; Hardy, R.W.; Hartenstein, V.; Zuker, C.S. InsP 3 receptor is essential for growth and differentiation but not for vision in Drosophila. Neuron 1997, 18, 881–887. [Google Scholar] [CrossRef]
- Raghu, P.; Colley, N.J.; Webel, R.; James, T.; Hasan, G.; Danin, M.; Selinger, Z.; Hardie, R.C. Normal phototransduction in Drosophila photoreceptors lacking an InsP 3 receptor gene. Mol. Cell Neurosci. 2000, 15, 429–445. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid Name | Used for |
|---|---|
| pcDNA4-TRPLWT-GFP | Protein expression in confocal imaging |
| pcDNA4-hTRPC3-mCherry | Protein expression in confocal imaging and electrophysiological experiments |
| pcDNA4-TRPLWT | Protein expression in electrophysiological experiments and Western blot analysis |
| pcDNA4-GFP | Protein expression in electrophysiological experiments |
| pcDNA4-hM1R | Protein expression in electrophysiological experiments |
| pcDNA4-TRPLF557I | Protein expression in electrophysiological experiments |
| Chemical Name | Concentration (mM) |
|---|---|
| CsMeSO3 | 120 |
| CsCl | 25 |
| MgCl2 | 1 |
| EGTA | 5 |
| MgATP | 3 |
| NaGTP | 0.3 |
| HEPES | 10 |
| Chemical Name | Concentration (mM) |
|---|---|
| NaCl | 147 |
| KCl | 5 |
| MgCl2 | 1 |
| HEPES | 10 |
| D-glucose | 10 |
| Chemical Name | Concentration (mM) |
|---|---|
| NaCl | 10 |
| MgCl2 | 1 |
| HEPES | 5 |
| EGTA | 1.5 |
| Na-gluconate | 140 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rhodes-Mordov, E.; Brandwine-Shemmer, T.; Zaguri, R.; Gutorov, R.; Peters, M.; Minke, B. Diacylglycerol Activates the Drosophila Light Sensitive Channel TRPL Expressed in HEK Cells. Int. J. Mol. Sci. 2023, 24, 6289. https://doi.org/10.3390/ijms24076289
Rhodes-Mordov E, Brandwine-Shemmer T, Zaguri R, Gutorov R, Peters M, Minke B. Diacylglycerol Activates the Drosophila Light Sensitive Channel TRPL Expressed in HEK Cells. International Journal of Molecular Sciences. 2023; 24(7):6289. https://doi.org/10.3390/ijms24076289
Chicago/Turabian StyleRhodes-Mordov, Elisheva, Tal Brandwine-Shemmer, Rachel Zaguri, Rita Gutorov, Maximilian Peters, and Baruch Minke. 2023. "Diacylglycerol Activates the Drosophila Light Sensitive Channel TRPL Expressed in HEK Cells" International Journal of Molecular Sciences 24, no. 7: 6289. https://doi.org/10.3390/ijms24076289
APA StyleRhodes-Mordov, E., Brandwine-Shemmer, T., Zaguri, R., Gutorov, R., Peters, M., & Minke, B. (2023). Diacylglycerol Activates the Drosophila Light Sensitive Channel TRPL Expressed in HEK Cells. International Journal of Molecular Sciences, 24(7), 6289. https://doi.org/10.3390/ijms24076289