
A Novel Multi-Observation System to Study the Effects of Anterior Ocular Inflammation in Zinn’s Zonule Using One Specimen

 , , , and
, , , and 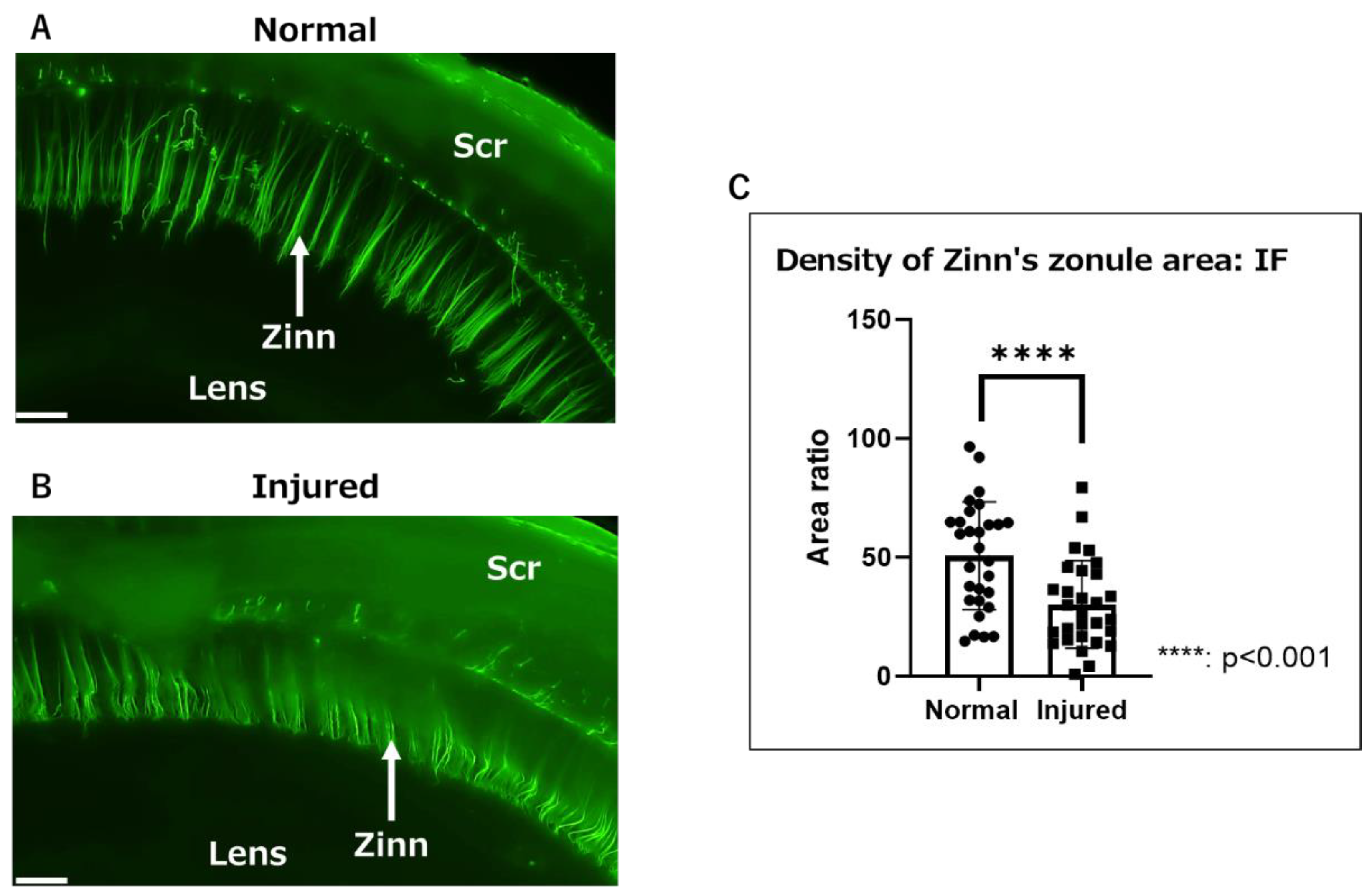{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Macroscopic Observation of Inflammatory Changes in Zinn’s Zonule Using Gel-Embedded Specimens
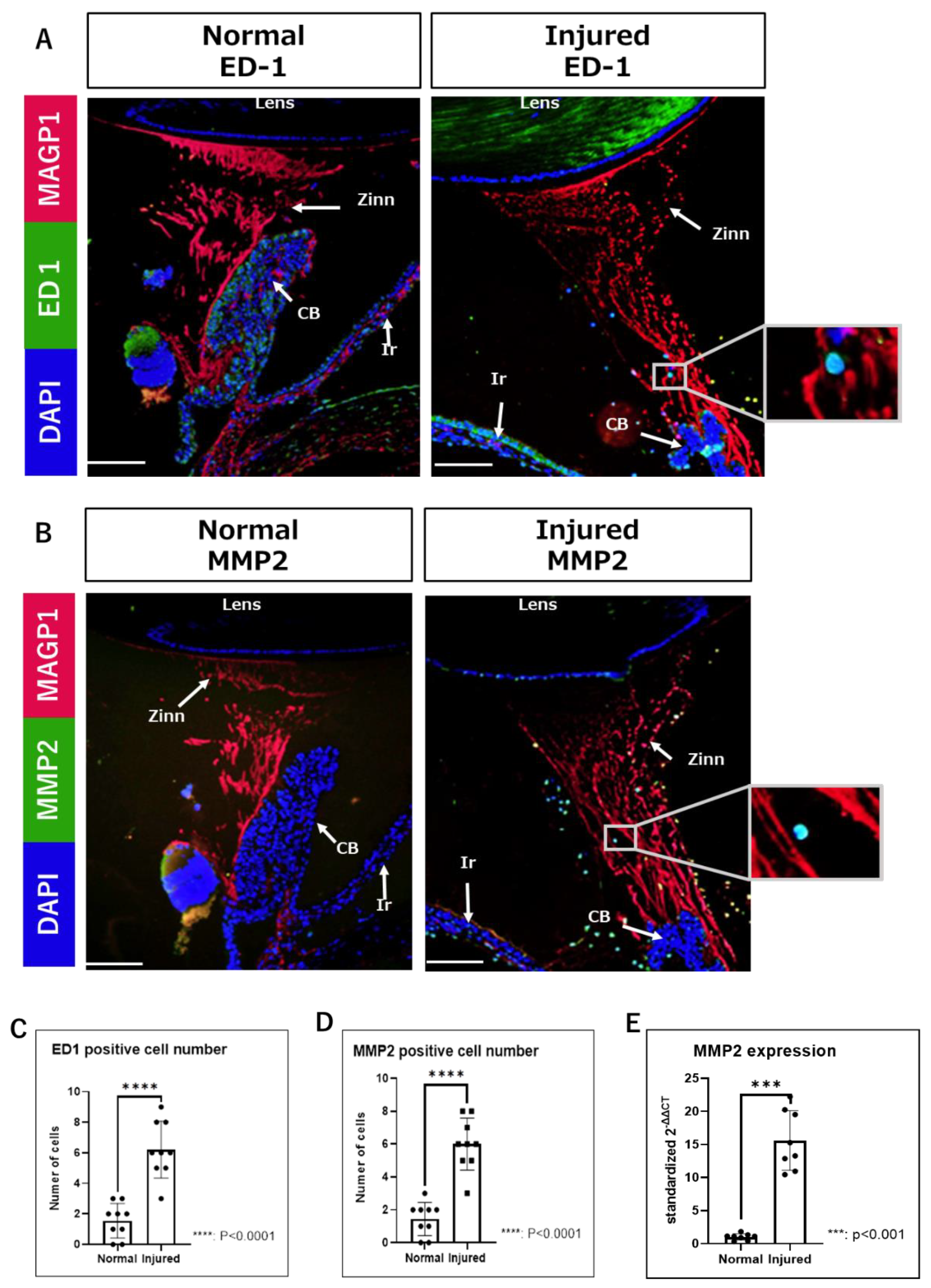
2.2. Observation of Inflammatory Changes in Zinn’s Zonule Using IF and RT-qPCR
2.3. Ultrastructural Observation of Inflammatory Changes in Zinn’s Zonule Using LV-SEM
3. Discussion
4. Materials and Methods
4.1. Animals and Alkali Burn Model
4.2. Gel- and Paraffin-Embedding of Eyes
4.3. IF Analysis
4.3.1. IF Staining of Gel-Embedded Specimens
4.3.2. Paraffin Sections
4.4. IHC with LV-SEM Analysis
4.5. RT-qPCR Analysis
4.6. Image Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ascaso, F.J.; Huerva, V.; Grzybowski, A. Epidemiology, etiology, and prevention of late IOL-capsular bag complex dislocation: Review of the literature. J. Ophthalmol. 2015, 2015, 805706. [Google Scholar] [CrossRef] [PubMed]
- Ladewig, M.S.; Robinson, P.N.; Neumann, L.M.; Holz, F.G.; Foerster, M.H. Ocular manifestations and surgical results in patients with Marfan syndrome. Ophthalmologe 2006, 103, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Canavan, Y.M.; Archer, D.B. Anterior segment consequences of blunt ocular injury. Br. J. Ophthalmol. 1982, 66, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Shingleton, B.J.; Neo, Y.N.; Cvintal, V.; Shaikh, A.M.; Liberman, P.; O’Donoghue, M.W. Outcome of phacoemulsification and intraocular lens implantation in eyes with pseudoexfoliation and weak zonules. Acta Ophthalmol. 2017, 95, 182–187. [Google Scholar] [CrossRef]
- Tao, L.W.; Hall, A. In-bag dislocation of intraocular lens in patients with uveitis: A case series. J. Ophthalmic Inflamm. Infect. 2015, 5, 10. [Google Scholar] [CrossRef]
- DeDreu, J.; Bowen, C.J.; Logan, C.M.; Pal-Ghosh, S.; Parlanti, P.; Stepp, M.A.; Menko, A.S. An immune response to the avascular lens following wounding of the cornea involves ciliary zonule fibrils. FASEB J. 2020, 34, 9316–9336. [Google Scholar] [CrossRef] [PubMed]
- Arai, Y.; Takeuchi, K.; Hatanaka, S.; Ishikawa, A.; Inoue, T.; Takakuma, S.; Kajimoto, Y.; Toda, E.; Kunugi, S.; Terasaki, M.; et al. Heavy metal enhancement technique for diaminobenzidine in immunohistochemistry enables ultrastructural observation by low-vacuum scanning electron microscopy. J. Histochem. Cytochem. 2022, 70, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Tu, Y.; De Maria, A.; Mecham, R.P.; Bassnett, S. Development, composition, and structural arrangements of the ciliary zonule of the mouse. Investig. Ophthalmol. Vis. Sci. 2013, 54, 2504–2515. [Google Scholar] [CrossRef]
- Shi, Y.; Jones, W.; Beatty, W.; Tan, Q.; Mecham, R.P.; Kumra, H.; Reinhardt, D.P.; Gibson, M.A.; Reilly, M.A.; Rodriguez, J.; et al. Latent-transforming growth factor beta-binding protein-2 (LTBP-2) is required for longevity but not for development of zonular fibers. Matrix Biol. 2021, 95, 15–31. [Google Scholar] [CrossRef]
- Bassnett, S. A method for preserving and visualizing the three-dimensional structure of the mouse zonule. Exp. Eye Res. 2019, 185, 107685. [Google Scholar] [CrossRef]
- Miyazaki, H.; Uozaki, H.; Tojo, A.; Hirashima, S.; Inaga, S.; Sakuma, K.; Morishita, Y.; Fukayama, M. Application of low-vacuum scanning electron microscopy for renal biopsy specimens. Pathol. Res. Pract. 2012, 208, 503–509. [Google Scholar] [CrossRef]
- Song, J.Y.; Saga, N.; Kawanishi, K.; Hashikami, K.; Takeyama, M.; Nagata, M. Bidirectional, non-necrotizing glomerular crescents are the critical pathology in X-linked Alport syndrome mouse model harboring nonsense mutation of human COL4A5. Sci. Rep. 2020, 10, 18891. [Google Scholar] [CrossRef]
- Fujimaru, T.; Kawanishi, K.; Mori, T.; Mishima, E.; Sekine, A.; Chiga, M.; Mizui, M.; Sato, N.; Yanagita, M.; Ooki, Y.; et al. Genetic background and clinicopathologic features of adult-onset nephronophthisis. Kidney Int. Rep. 2021, 6, 1346–1354. [Google Scholar] [CrossRef] [PubMed]
- Arima, T.; Uchiyama, M.; Shimizu, A.; Takahashi, H. Observation of corneal wound healing and angiogenesis using low-vacuum scanning electron microscopy. Transl. Vis. Sci. Technol. 2020, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Ikebukuro, T.; Arima, T.; Kasamatsu, M.; Nakano, Y.; Tobita, Y.; Uchiyama, M.; Terashima, Y.; Toda, E.; Shimizu, A.; Takahashi, H. Disulfiram ophthalmic solution inhibited macrophage infiltration by suppressing macrophage pseudopodia formation in a rat corneal alkali burn model. Int. J. Mol. Sci. 2023, 24, 735. [Google Scholar] [CrossRef]
- Sivak, J.M.; Fini, M.E. MMPs in the eye: Emerging roles for matrix metalloproteinases in ocular physiology. Prog. Retin. Eye Res. 2002, 21, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Wu, H.; Reinach, P.S.; Wu, Y.; Zhai, Y.; Lei, Y.; Ma, L.; Su, Y.; Chen, Y.; Li, F.; et al. Up-regulation of matrix metalloproteinase-2 by scleral monocyte-derived macrophages contributes to myopia development. Am. J. Pathol. 2020, 190, 1888–1908. [Google Scholar] [CrossRef]
- Shiroto, Y.; Saga, R.; Yoshino, H.; Hosokawa, Y.; Isokawa, K.; Tsuruga, E. Matrix metalloproteinase-2 activated by ultraviolet-B degrades human ciliary zonules. Acta Histochem. Cytochem. 2021, 54, 1–9. [Google Scholar] [CrossRef]
- Ashworth, J.L.; Murphy, G.; Rock, M.J.; Sherratt, M.J.; Shapiro, S.D.; Shuttleworth, C.A.; Kielty, C.M. Fibrillin degradation by matrix metalloproteinases: Implications for connective tissue remodelling. Biochem. J. 1999, 340, 171–181. [Google Scholar] [CrossRef]
- Kawagoe, M.; Tsuruga, E.; Oka, K.; Sawa, Y.; Ishikawa, H. Matrix metalloproteinase-2 degrades fibrillin-1 and fibrillin-2 of oxytalan fibers in the human eye and periodontal ligaments in vitro. Acta. Histochem. Cytochem. 2013, 46, 153–159. [Google Scholar] [CrossRef]
- Ushida, K.; Asai, N.; Uchiyama, K.; Enomoto, A.; Takahashi, M. Development of a method to preliminarily embed tissue samples using low melting temperature fish gelatin before sectioning: A technical note. Pathol. Int. 2018, 68, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Bassnett, S. Zinn’s zonule. Prog. Retin. Eye Res. 2021, 82, 100902. [Google Scholar] [CrossRef] [PubMed]
- Mecham, R.P.; Gibson, M.A. The microfibril-associated glycoproteins (MAGPs) and the microfibrillar niche. Matrix Biol. 2015, 47, 13–33. [Google Scholar] [CrossRef] [PubMed]
- LeVaillant, C.J.; Sharma, A.; Muhling, J.; Wheeler, L.P.; Cozens, G.S.; Hellström, M.; Rodger, J.; Harvey, A.R. Significant changes in endogenous retinal gene expression assessed 1 year after a single intraocular injection of AAV-CNTF or AAV-BDNF. Mol. Ther. Methods Clin. Dev. 2016, 3, 16078. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takahashi, A.; Arima, T.; Toda, E.; Kobayakawa, S.; Shimizu, A.; Takahashi, H. A Novel Multi-Observation System to Study the Effects of Anterior Ocular Inflammation in Zinn’s Zonule Using One Specimen. Int. J. Mol. Sci. 2023, 24, 6254. https://doi.org/10.3390/ijms24076254
Takahashi A, Arima T, Toda E, Kobayakawa S, Shimizu A, Takahashi H. A Novel Multi-Observation System to Study the Effects of Anterior Ocular Inflammation in Zinn’s Zonule Using One Specimen. International Journal of Molecular Sciences. 2023; 24(7):6254. https://doi.org/10.3390/ijms24076254
Chicago/Turabian StyleTakahashi, Akira, Takeshi Arima, Etsuko Toda, Shinichiro Kobayakawa, Akira Shimizu, and Hiroshi Takahashi. 2023. "A Novel Multi-Observation System to Study the Effects of Anterior Ocular Inflammation in Zinn’s Zonule Using One Specimen" International Journal of Molecular Sciences 24, no. 7: 6254. https://doi.org/10.3390/ijms24076254
APA StyleTakahashi, A., Arima, T., Toda, E., Kobayakawa, S., Shimizu, A., & Takahashi, H. (2023). A Novel Multi-Observation System to Study the Effects of Anterior Ocular Inflammation in Zinn’s Zonule Using One Specimen. International Journal of Molecular Sciences, 24(7), 6254. https://doi.org/10.3390/ijms24076254