
Interplay of Vitamin D and SIRT1 in Tissue-Specific Metabolism—Potential Roles in Prevention and Treatment of Non-Communicable Diseases Including Cancer

Abstract
1. Introduction
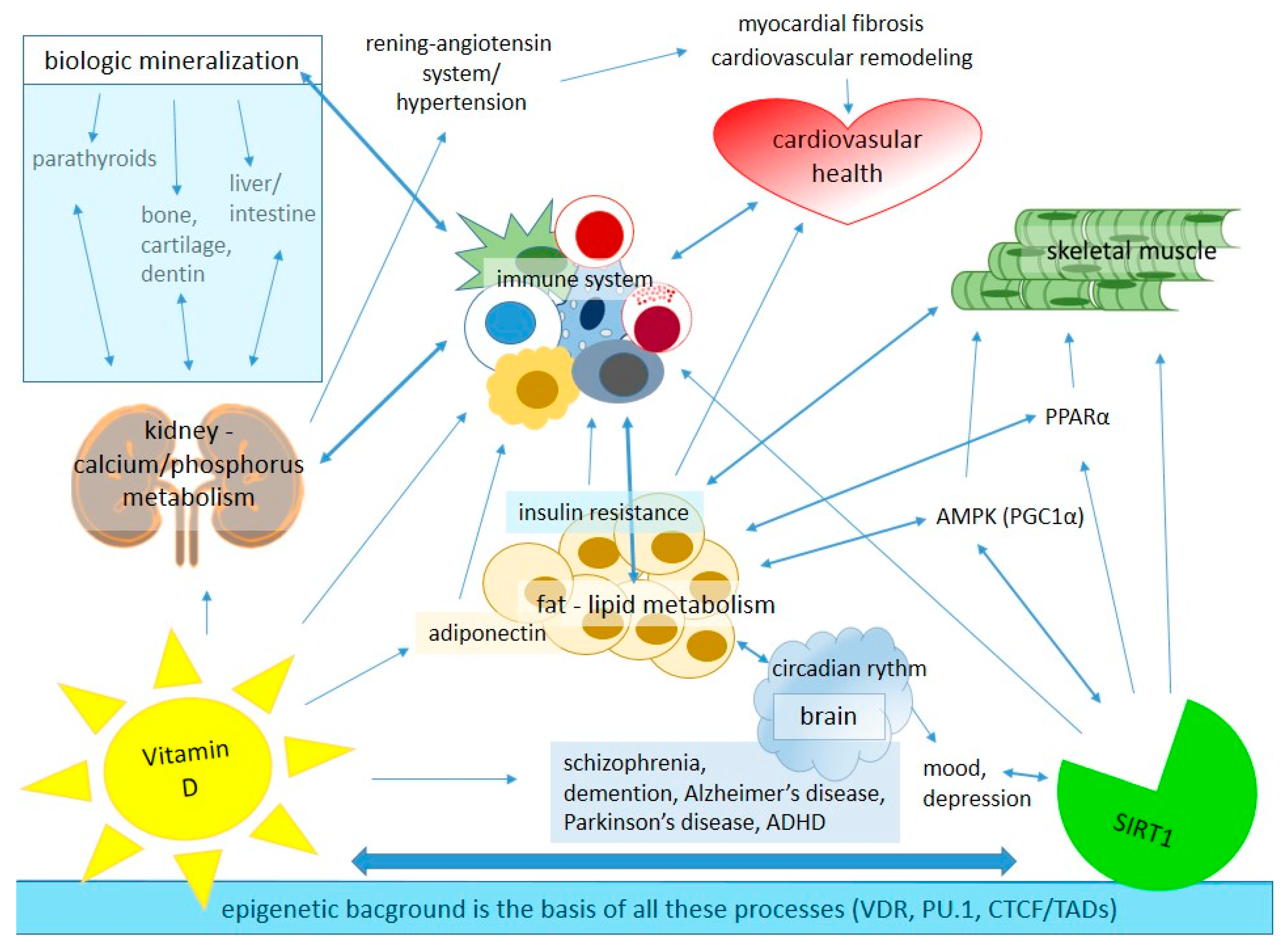
2. The Role of Vitamin D and SIRT1 in Healthy Metabolic Regulation of Different Tissues
2.1. Modes of Action of Vitamin D
2.2. Role of Vitamin D in Lipid Metabolism
2.3. Role of Vitamin D in Cardiovascular System
2.4. Role of Vitamin D in the Immune System
2.5. Role of Vitamin D in Redox Homeostasis
2.6. Role of SIRT1 in Healthy Metabolic Regulation
2.7. Connection and Importance of Circadian Rhythm in SIRT1-Regulated Metabolism
3. Vitamin D and SIRT1 in Relation to Non-Communicable Diseases Including Cancer
3.1. Effect of Vitamin D and SIRT1 on Metabolic Syndrome
3.2. Effect of Vitamin D and SIRT1 on CVD
3.3. Effect of Vitamin D and SIRT1 on CKD
3.4. Effect of Vitamin D and SIRT1 on Immune System-Related Diseases
3.5. Effect of Vitamin D and SIRT1 on Cancer Cells
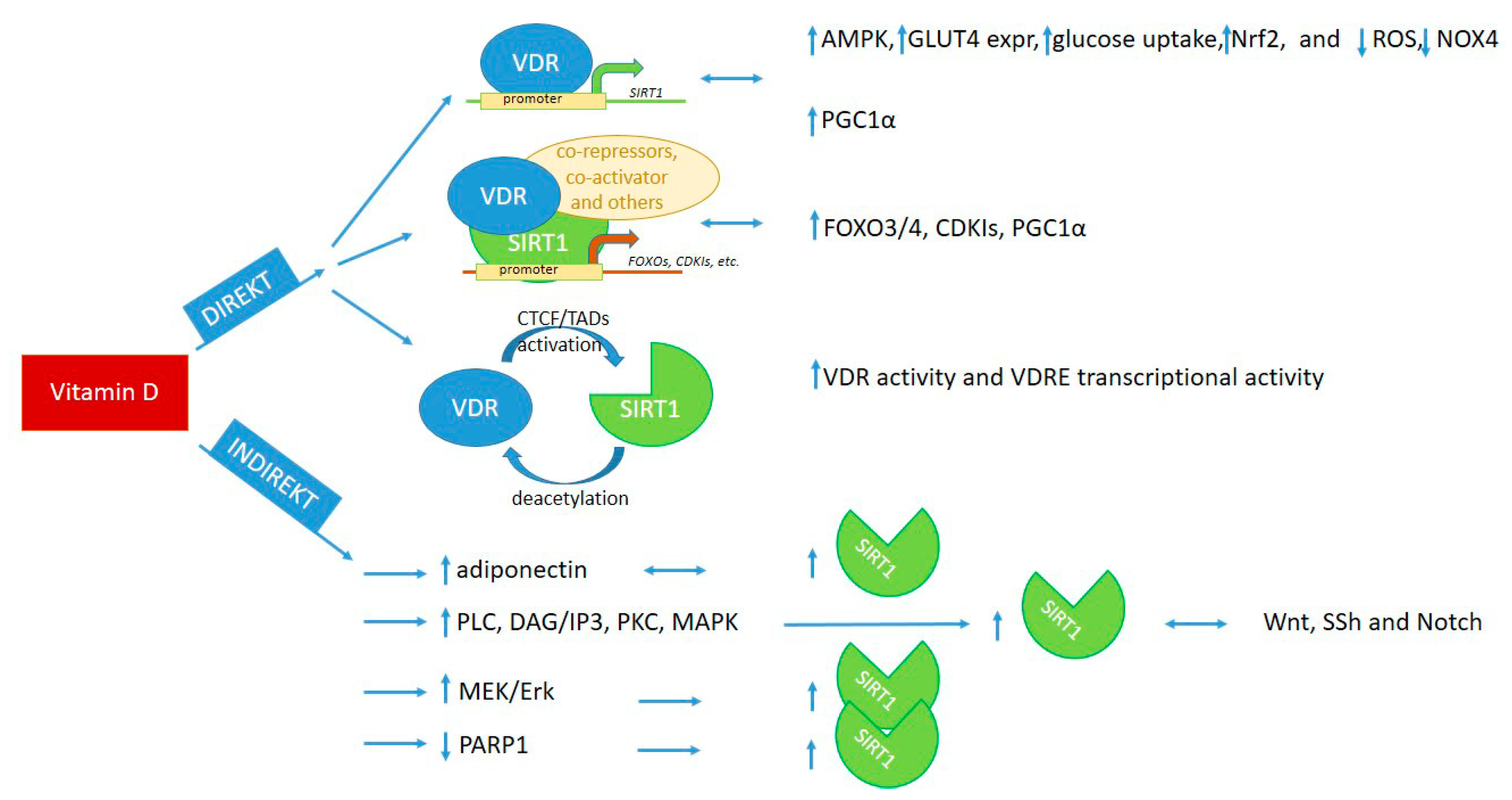
4. Cooperation of Vitamin D and SIRT1 Pathways
4.1. Direct Interaction of Vitamin D and SIRT1
4.1.1. Interaction through Binding of VDR to SIRT1 Promoter Region
4.1.2. Interaction through Binding of VDR to SIRT1 Protein or via Epigenetic Modifications of Each Other
4.2. Indirect Interaction of Vitamin D and SIRT1
5. Potential Applications of Vitamin D and Induction of SIRT1 in the Prevention, Treatment, and Reduction of Mortality Risk of Non-Communicable Diseases
5.1. Guidelines for Vitamin D Status and Its Daily Intake
5.2. Recommendation for Healthy Individuals to Achieve the 30–50 ng/mL of Serum 25(OH)D3 Level
5.3. Recommendation for Patients with Diseases to Achieve the 30–50 ng/mL of Serum 25(OH)D3 Level
5.4. Recommendation to Maintain an Optimal SIRT1 Level in the Body
5.5. Recommendation to Preventive and Complementary Treatments
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martins, I.J. COVID-19 Infection and Anti-Aging Gene Inactivation. Acta Sci. Nutr. Health 2020, 4, 29554–29573. [Google Scholar] [CrossRef]
- Nadalin, S.; Jakovac, H.; Peitl, V.; Karlović, D.; Buretić-Tomljanović, A. Dysregulated inflammation may predispose patients with serious mental illnesses to severe COVID-19 (Review). Mol. Med. Rep. 2021, 24, 611. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.; Kim, Y. Vitamin D Ameliorates Fat Accumulation with AMPK/SIRT1 Activity in C2C12 Skeletal Muscle Cells. Nutrients 2019, 11, 2806. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, E.A.T.; Peñafiel, A.M.; Pedraza, A.G.; Leo, E.E.M. Molecular mechanisms from insulin-mimetic effect of vitamin D: Treatment alternative in Type 2 diabetes mellitus. Food Funct. 2021, 12, 6682–6690. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Shen, P.; Liu, S.; Wang, J.; Zeng, J.; Du, C. Vitamin D alleviates skeletal muscle loss and insulin resistance by inducing vitamin D receptor expression and regulating the AMPK/SIRT1 signaling pathway in mice. Food Sci. Technol. 2022, 42, e47921. [Google Scholar] [CrossRef]
- Yuan, Q.; Yang, J.; Sun, M.; Tang, S.; Dong, M.; Mao, L. Vitamin D deficiency impairs mTorc2/Akt signalling through down regulating Sirt1 and results in increased hepatic gluconeogenesis. Diabetologia 2020, 63, S222. [Google Scholar]
- Manna, P.; Achari, A.E.; Jain, S.K. Vitamin D supplementation inhibits oxidative stress and upregulate SIRT1/AMPK/GLUT4 cascade in high glucose-treated 3T3L1 adipocytes and in adipose tissue of high fat diet-fed diabetic mice. Arch. Biochem. Biophys. 2017, 615, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Polidoro, L.; Properzi, G.; Marampon, F.; Gravina, G.L.; Festuccia, C.; Di Cesare, E.; Scarsella, L.; Ciccarelli, C.; Zani, B.M.; Ferri, C. Vitamin D Protects Human Endothelial Cells from H2O2 Oxidant Injury Through the Mek/Erk-Sirt1 Axis Activation. J. Cardiovasc. Transl. Res. 2013, 6, 221–231. [Google Scholar] [CrossRef]
- Lin, Z.; Teng, C.; Ni, L.; Zhang, Z.; Lu, X.; Lou, J.; Wang, L.; Wang, Y.; Chen, W.; Zhang, X.; et al. Echinacoside Upregulates Sirt1 to Suppress Endoplasmic Reticulum Stress and Inhibit Extracellular Matrix Degradation In Vitro and Ameliorates Osteoarthritis In Vivo. Oxidative Med. Cell. Longev. 2021, 2021, 3137066. [Google Scholar] [CrossRef]
- Jia, R.J.; Yang, F.; Yan, P.F.; Ma, L.M.; Yang, L.F.; Li, L.H. Paricalcitol inhibits oxidative stress-induced cell senescence of the bile duct epithelium dependent on modulating Sirt1 pathway in cholestatic mice. Free. Radic. Biol. Med. 2021, 169, 158–168. [Google Scholar] [CrossRef]
- Charoenngam, N.; Holick, M.F. Immunologic Effects of Vitamin D on Human Health and Disease. Nutrients 2020, 12, 2097. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.; Kim, Y. Vitamin D Insufficiency Exacerbates Adipose Tissue Macrophage Infiltration and Decreases AMPK/SIRT1 Activity in Obese Rats. Nutrients 2017, 9, 338. [Google Scholar] [CrossRef] [PubMed]
- Karlic, H.; Varga, F. Impact of vitamin D metabolism on clinical epigenetics. Clin. Epigenetics 2011, 2, 55–61. [Google Scholar] [CrossRef]
- Yuan, Q.; Zhang, R.D.; Sun, M.Y.; Guo, X.; Yang, J.L.; Bian, W.; Xie, C.; Miao, D.; Mao, L. Sirt1 Mediates Vitamin D Deficiency-Driven Gluconeogenesis in the Liver via mTorc2/Akt Signaling. J. Diabetes Res. 2022, 2022, 1755563. [Google Scholar] [CrossRef] [PubMed]
- Abiri, B.; Vafa, M. Effects of vitamin D and/or magnesium supplementation on mood, serum levels of BDNF, inflammatory biomarkers, and SIRT1 in obese women: A study protocol for a double-blind, randomized, placebo-controlled trial. Trials 2020, 21, 225. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.-L.; Liao, M.-T.; Hou, Y.-C.; Fang, Y.-W.; Zheng, C.-M.; Liu, W.-C.; Chao, C.-T.; Lu, K.-C.; Ng, Y.-Y. Sirtuin-1 and Its Relevance in Vascular Calcification. Int. J. Mol. Sci. 2020, 21, 1593. [Google Scholar] [CrossRef]
- Liu, Y.; Peng, W.; Li, Y.; Wang, B.; Yu, J.; Xu, Z. Vitamin D Deficiency Harms Patients with Coronary Heart Disease by Enhancing Inflammation. Experiment 2018, 24, 9376–9384. [Google Scholar] [CrossRef]
- Al-Khalidi, B.; Kimball, S.M.; Kuk, J.L.; Ardern, C.I. Metabolically healthy obesity, vitamin D, and all-cause and cardiometabolic mortality risk in NHANES III. Clin. Nutr. 2019, 38, 820–828. [Google Scholar] [CrossRef]
- Degerud, E.; Nygård, O.; De Vogel, S.; Hoff, R.; Svingen, G.F.T.; Pedersen, E.R.; Nilsen, D.W.T.; Nordrehaug, J.E.; Midttun, M.; Ueland, P.M.; et al. Plasma 25-Hydroxyvitamin D and Mortality in Patients with Suspected Stable Angina Pectoris. J. Clin. Endocrinol. Metab. 2018, 103, 1161–1170. [Google Scholar] [CrossRef]
- Gaksch, M.; Jorde, R.; Grimnes, G.; Joakimsen, R.; Schirmer, H.; Wilsgaard, T.; Mathiesen, E.B.; Njølstad, I.; Løchen, M.-L.; März, W.; et al. Vitamin D and mortality: Individual participant data meta-analysis of standardized 25-hydroxyvitamin D in 26916 individuals from a European consortium. PLoS ONE 2017, 12, e0170791. [Google Scholar] [CrossRef]
- Hu, K.; Callen, D.F.; Li, J.; Zheng, H. Circulating Vitamin D and Overall Survival in Breast Cancer Patients: A Dose-Response Meta-Analysis of Cohort Studies. Integr. Cancer Ther. 2018, 17, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-Q.; Langhammer, A.; Skorpen, F.; Chen, Y.; Mai, X.-M. Serum 25-hydroxyvitamin D level, chronic diseases and all-cause mortality in a population-based prospective cohort: The HUNT Study, Norway. BMJ Open 2017, 7, e017256. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Knuiman, M.; Divitini, M.; Hung, J.; Lim, E.M.; Cooke, B.R.; Walsh, J.P. Serum 25-hydroxyvitamin D as a predictor of mortality and cardiovascular events: A 20-year study of a community-based cohort. Clin. Endocrinol. 2018, 88, 154–163. [Google Scholar] [CrossRef]
- Pereira, M.; Costa, P.R.F.; Pereira, E.M.; Lago, I.R.L.; Oliveira, A.M. Does vitamin D deficiency increase the risk of obesity in adults and the elderly? A systematic review of prospective cohort studies. Public Health 2021, 190, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Melguizo-Rodríguez, L.; Costela-Ruiz, V.; García-Recio, E.; De Luna-Bertos, E.; Ruiz, C.; Illescas-Montes, R. Role of Vitamin D in the Metabolic Syndrome. Nutrients 2021, 13, 830. [Google Scholar] [CrossRef]
- DiNicolantonio, J.J.; O’Keefe, J.H. Magnesium and Vitamin D Deficiency as a Potential Cause of Immune Dysfunction, Cytokine Storm and Disseminated Intravascular Coagulation in COVID-19 patients. Mo. Med. 2021, 118, 68–73. [Google Scholar]
- Rafiq, S.; Jeppesen, P.B. Vitamin D Deficiency Is Inversely Associated with Homeostatic Model Assessment of Insulin Resistance. Nutrients 2021, 13, 4358. [Google Scholar] [CrossRef]
- Al-Ishaq, R.K.; Kubatka, P.; Brozmanova, M.; Gazdikova, K.; Caprnda, M.; Büsselberg, D. Health implication of vitamin D on the cardiovascular and the renal system. Arch. Physiol. Biochem. 2021, 127, 195–209. [Google Scholar] [CrossRef]
- Ham, N.S.; Hwang, S.W.; Oh, E.H.; Kim, J.; Lee, H.-S.; Park, S.H.; Yang, D.-H.; Ye, B.D.; Byeon, J.-S.; Myung, S.-J.; et al. Influence of Severe Vitamin D Deficiency on the Clinical Course of Inflammatory Bowel Disease. Dig. Dis. Sci. 2021, 66, 587–596. [Google Scholar] [CrossRef]
- Diaconu, A.-D.; Ostafie, I.; Ceasovschih, A.; Șorodoc, V.; Lionte, C.; Ancuța, C.; Șorodoc, L. Role of Vitamin D in Systemic Sclerosis: A Systematic Literature Review. J. Immunol. Res. 2021, 2021, 9782994. [Google Scholar] [CrossRef]
- Voutsadakis, I.A. Vitamin D baseline levels at diagnosis of breast cancer: A systematic review and meta-analysis. Hematol. Stem Cell Ther. 2020, 14, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Carlberg, C.; Muñoz, A. An update on vitamin D signaling and cancer. Semin. Cancer Biol. 2022, 79, 217–230. [Google Scholar] [CrossRef]
- Nithila, M.R.; Roy, N.M.; Al-Harthi, L.; Sampat, N.; Al-Mujaini, R.; Mahadevan, S.; Al Adawi, S.; Essa, M.M.; Al Subhi, L.; Al-Balushi, B.; et al. Impact of vitamin D on neurocognitive function in dementia, depression, schizophrenia and ADHD. Front. Biosci. 2021, 26, 566–611. [Google Scholar] [CrossRef]
- Nemeth, Z.; Kiss, E.; Takacs, I. The Role of Epigenetic Regulator SIRT1 in Balancing the Homeostasis and Preventing the Formation of Specific “Soil” of Metabolic Disorders and Related Cancers. Front. Biosci. 2022, 27, 253. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, Z.; Kiss, E.; Takacs, I. A Healthy Balance of Homeostasis by Epigenetic Regulator SIRT1 May Prevent the Development of a Specific “Soil” that Supports Metabolic Disorders and Related Cancers. In Current Overview on Disease and Health Research; Karaman, D.R., Ed.; B P International: London, UK, 2022; Volume 4, pp. 71–111. [Google Scholar] [CrossRef]
- Fernández-Ruiz, M.; Corbella, L.; Morales-Cartagena, A.; González, E.; Polanco, N.; Ruiz-Merlo, T.; Parra, P.; Silva, J.T.; López-Medrano, F.; Juan, R.S.; et al. Vitamin D deficiency and infection risk in kidney transplant recipients: A single-center cohort study. Transpl. Infect. Dis. 2018, 20, e12988. [Google Scholar] [CrossRef]
- Schreiber, P.W.; Bischoff-Ferrari, H.A.; Boggian, K.; van Delden, C.; Enriquez, N.; Fehr, T.; Garzoni, C.; Hirsch, H.H.; Hirzel, C.; Manuel, O.; et al. Vitamin D status and risk of infections after liver transplantation in the Swiss Transplant Cohort Study. Transpl. Int. 2019, 32, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Mirzakhani, M.; Mohammadkhani, S.; Hekmatirad, S.; Aghapour, S.; Gorjizadeh, N.; Shahbazi, M.; Mohammadnia-Afrouzi, M. The association between vitamin D and acute rejection in human kidney transplantation: A systematic review and meta-analysis study. Transpl. Immunol. 2021, 67, 101410. [Google Scholar] [CrossRef]
- Zhou, Q.; Li, L.; Chen, Y.; Zhang, J.; Zhong, L.; Peng, Z.; Xing, T. Vitamin D supplementation could reduce the risk of acute cellular rejection and infection in vitamin D deficient liver allograft recipients. Int. Immunopharmacol. 2019, 75, 105811. [Google Scholar] [CrossRef]
- Ponticelli, C.; Favi, E.; Ferraresso, M. New-Onset Diabetes after Kidney Transplantation. Medicina 2021, 57, 250. [Google Scholar] [CrossRef]
- Bouillon, R.; Marcocci, C.; Carmeliet, G.; Bikle, D.; White, J.H.; Dawson-Hughes, B.; Lips, P.; Munns, C.F.; Lazaretti-Castro, M.; Giustina, A.; et al. Skeletal and Extraskeletal Actions of Vitamin D: Current Evidence and Outstanding Questions. Endocr. Rev. 2019, 40, 1109–1151. [Google Scholar] [CrossRef]
- Dimitrov, V.; Barbier, C.; Ismailova, A.; Wang, Y.; Dmowski, K.; Salehi-Tabar, R.; Memari, B.; Groulx-Boivin, E.; White, J.H. Vitamin D-regulated Gene Expression Profiles: Species-specificity and Cell-specific Effects on Metabolism and Immunity. Endocrinology 2021, 162, bqaa218. [Google Scholar] [CrossRef] [PubMed]
- Carlberg, C. Vitamin D and Its Target Genes. Nutrients 2022, 14, 1354. [Google Scholar] [CrossRef]
- Zmijewski, M.A.; Carlberg, C. Vitamin D receptor(s): In the nucleus but also at membranes? Exp. Dermatol. 2020, 29, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Haberle, V.; Stark, A. Eukaryotic core promoters and the functional basis of transcription initiation. Nat. Rev. Mol. Cell Biol. 2018, 19, 621–637. [Google Scholar] [CrossRef]
- Tuoresmäki, P.; Väisänen, S.; Neme, A.; Heikkinen, S.; Carlberg, C. Patterns of Genome-Wide VDR Locations. PLoS ONE 2014, 9, e96105. [Google Scholar] [CrossRef]
- Neme, A.; Seuter, S.; Carlberg, C. Vitamin D-dependent chromatin association of CTCF in human monocytes. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2016, 1859, 1380–1388. [Google Scholar] [CrossRef] [PubMed]
- Seuter, S.; Neme, A.; Carlberg, C. Epigenome-wide effects of vitamin D and their impact on the transcriptome of human monocytes involve CTCF. Nucleic Acids Res. 2016, 44, 4090–4104. [Google Scholar] [CrossRef]
- Nikooyeh, B.; Hollis, B.W.; Neyestani, T.R. The effect of daily intake of vitamin D-fortified yogurt drink, with and without added calcium, on serum adiponectin and sirtuins 1 and 6 in adult subjects with type 2 diabetes. Nutr. Diabetes 2021, 11, 26. [Google Scholar] [CrossRef]
- Lee, Y.; Ka, S.-O.; Cha, H.-N.; Chae, Y.-N.; Kim, M.-K.; Park, S.-Y.; Bae, E.J.; Park, B.-H. Myeloid Sirtuin 6 Deficiency Causes Insulin Resistance in High-Fat Diet–Fed Mice by Eliciting Macrophage Polarization Toward an M1 Phenotype. Diabetes 2017, 66, 2659–2668. [Google Scholar] [CrossRef]
- Bae, E.J. Sirtuin 6, a possible therapeutic target for type 2 diabetes. Arch. Pharmacal Res. 2017, 40, 1380–1389. [Google Scholar] [CrossRef]
- Martinez-Huenchullan, S.F.; Tam, C.S.; Ban, L.A.; Ehrenfeld-Slater, P.; Mclennan, S.V.; Twigg, S.M. Skeletal muscle adiponectin induction in obesity and exercise. Metabolism 2020, 102, 154008. [Google Scholar] [CrossRef]
- Yanai, H.; Yoshida, H. Beneficial Effects of Adiponectin on Glucose and Lipid Metabolism and Atherosclerotic Progression: Mechanisms and Perspectives. Int. J. Mol. Sci. 2019, 20, 1190. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Shimomura, I. Roles of adiponectin and oxidative stress in obesity-associated metabolic and cardiovascular diseases. Rev. Endocr. Metab. Disord. 2014, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Walsh, K. Adiponectin as an anti-inflammatory factor. Clin. Chim. Acta 2007, 380, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.M.; Wolf, D.; Rumpold, H.; Enrich, B.; Tilg, H. Adiponectin induces the anti-inflammatory cytokines IL-10 and IL-1RA in human leukocytes. Biochem. Biophys. Res. Commun. 2004, 323, 630–635. [Google Scholar] [CrossRef]
- Gardner, D.G.; Chen, S.; Glenn, D.J.; Ni, W. Chapter 31—Vitamin D and the Cardiovascular System. In Vitamin D, 3rd ed.; Feldman, D., Pike, J.W., Adams, J.S., Eds.; Academic Press: San Diego, CA, USA, 2011; pp. 541–563. [Google Scholar]
- Merke, J.; Milde, P.; Lewicka, S.; Hügel, U.; Klaus, G.; Mangelsdorf, D.; Haussler, M.R.; Rauterberg, E.W.; Ritz, E. Identification and regulation of 1,25-dihydroxyvitamin D3 receptor activity and biosynthesis of 1,25-dihydroxyvitamin D3. Studies in cultured bovine aortic endothelial cells and human dermal capillaries. J. Clin. Investig. 1989, 83, 1903–1915. [Google Scholar] [CrossRef]
- Merke, J.; Hofmann, W.; Goldschmidt, D.; Ritz, E. Demonstration of 1,25(OH)2 vitamin D3 receptors and actions in vascular smooth muscle cells in vitro. Calcif. Tissue Int. 1987, 41, 112–114. [Google Scholar] [CrossRef]
- O’Connell, T.D.; Simpson, R.U. Immunochemical identification of the 1,25-dihydroxyvitamin D3 receptor protein in human heart. Cell Biol. Int. 1996, 20, 621–624. [Google Scholar] [CrossRef]
- Wong, M.S.K.; Delansorne, R.; Man, R.Y.K.; Vanhoutte, P.M. Vitamin D derivatives acutely reduce endothelium-dependent contractions in the aorta of the spontaneously hypertensive rat. Am. J. Physiol. Circ. Physiol. 2008, 295, H289–H296. [Google Scholar] [CrossRef]
- Wu-Wong, J.R.; Noonan, W.; Nakane, M.; Brooks, K.A.; Segreti, J.A.; Polakowski, J.S.; Cox, P. Vitamin d receptor activation mitigates the impact of uremia on endothelial function in the 5/6 nephrectomized rats. Int. J. Endocrinol. 2010, 2010, 625852. [Google Scholar] [CrossRef]
- Towler, D.A. Chapter 73—Vitamin D: Cardiovascular Effects and Vascular Calcification. In Vitamin D, 3rd ed.; Feldman, D., Pike, J.W., Adams, J.S., Eds.; Academic Press: San Diego, CA, USA, 2011; pp. 1403–1426. [Google Scholar]
- Badi, I.; Mancinelli, L.; Polizzotto, A.; Ferri, D.; Zeni, F.; Burba, I.; Milano, G.; Brambilla, F.; Saccu, C.; Bianchi, M.E.; et al. miR-34a Promotes Vascular Smooth Muscle Cell Calcification by Downregulating SIRT1 (Sirtuin 1) and Axl (AXL Receptor Tyrosine Kinase). Arter. Thromb. Vasc. Biol. 2018, 38, 2079–2090. [Google Scholar] [CrossRef]
- O’Neill, W. The fallacy of the calcium-phosphorus product. Kidney Int. 2007, 72, 792–796. [Google Scholar] [CrossRef]
- Demer, L.L.; Tintut, Y. Vascular calcification: Pathobiology of a multifaceted disease. Circulation 2008, 117, 2938–2948. [Google Scholar] [CrossRef] [PubMed]
- Tintut, Y.; Patel, J.; Territo, M.; Saini, T.; Parhami, F.; Demer, L.L. Monocyte/Macrophage Regulation of Vascular Calcification In Vitro. Circulation 2002, 105, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Nadra, I.; Mason, J.C.; Philippidis, P.; Florey, O.; Smythe, C.D.; McCarthy, G.M.; Landis, R.; Haskard, O. Proinflammatory activation of macrophages by basic calcium phosphate crystals via protein kinase C and MAP kinase pathways: A vicious cycle of inflammation and arterial calcification? Circ. Res. 2005, 96, 1248–1256. [Google Scholar] [CrossRef] [PubMed]
- Husain, K.; Ferder, L.; Mizobuchi, M.; Finch, J.; Slatopolsky, E. Combination Therapy with Paricalcitol and Enalapril Ameliorates Cardiac Oxidative Injury in Uremic Rats. Am. J. Nephrol. 2009, 29, 465–472. [Google Scholar] [CrossRef]
- Riek, A.E.; Oh, J.; Bernal-Mizrachi, C. Vitamin D regulates macrophage cholesterol metabolism in diabetes. J. Steroid Biochem. Mol. Biol. 2010, 121, 430–433. [Google Scholar] [CrossRef] [PubMed]
- Mantell, D.J.; Owens, P.E.; Bundred, N.J.; Mawer, E.B.; Canfield, A.E. 1 alpha,25-dihydroxyvitamin D(3) inhibits angiogenesis in vitro and in vivo. Circ Res. 2000, 87, 214–220. [Google Scholar] [CrossRef]
- Koh, E.; Morimoto, S.; Fukuo, K.; Itoh, K.; Hironaka, T.; Shiraishi, T.; Onishi, T.; Kumahara, Y. 1,25-dihydroxyvitamin D3 binds specifically to rat vascular smooth muscle cells and stimulates their proliferation in vitro. Life Sci. 1988, 42, 215–223. [Google Scholar] [CrossRef]
- Cardús, A.; Parisi, E.; Gallego, C.; Aldea, M.; Fernández, E.; Valdivielso, J. 1,25-Dihydroxyvitamin D3 stimulates vascular smooth muscle cell proliferation through a VEGF-mediated pathway. Kidney Int. 2006, 69, 1377–1384. [Google Scholar] [CrossRef]
- Hanel, A.; Carlberg, C. Vitamin D and evolution: Pharmacologic implications. Biochem. Pharmacol. 2020, 173, 113595. [Google Scholar] [CrossRef]
- Chun, R.; Liu, P.T.; Modlin, R.L.; Adams, J.S.; Hewison, M. Impact of vitamin D on immune function: Lessons learned from genome-wide analysis. Front. Physiol. 2014, 5, 151. [Google Scholar] [CrossRef]
- Vintilescu, B.; Niculescu, C.E.; Stepan, M.D.; Ioniță, E. Involvement of Vitamin D in Chronic Infections of the Waldeyer’s Ring in the School Aged Child. Curr. Health Sci. J. 2019, 45, 291–295. [Google Scholar] [CrossRef]
- Koivisto, O.; Hanel, A.; Carlberg, C. Key Vitamin D Target Genes with Functions in the Immune System. Nutrients 2020, 12, 1140. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, J.; DeLuca, H.F. Where is the vitamin D receptor? Arch. Biochem. Biophys. 2012, 523, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Bouillon, R.; Carmeliet, G.; Verlinden, L.; van Etten, E.; Verstuyf, A.; Luderer, H.F.; Lieben, L.; Mathieu, C.; Demay, M. Vitamin D and Human Health: Lessons from Vitamin D Receptor Null Mice. Endocr. Rev. 2008, 29, 726–776. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; McComish, B.J.; Burdon, K.P.; Taylor, B.V.; Körner, H. The Association Between Vitamin D and Multiple Sclerosis Risk: 1,25(OH)2D3 Induces Super-Enhancers Bound by VDR. Front. Immunol. 2019, 10, 488. [Google Scholar] [CrossRef]
- Caraba, A.; Crişan, V.; Romoşan, I.; Mozos, I.; Murariu, M.-S. Vitamin D Status, Disease Activity, and Endothelial Dysfunction in Early Rheumatoid Arthritis Patients. Dis. Markers 2017, 2017, 5241012. [Google Scholar] [CrossRef]
- Harrison, S.R.; Li, D.; Jeffery, L.E.; Raza, K.; Hewison, M. Vitamin D, Autoimmune Disease and Rheumatoid Arthritis. Calcif. Tissue Int. 2020, 106, 58–75. [Google Scholar] [CrossRef]
- Hanel, A.; Neme, A.; Malinen, M.; Hämäläinen, E.; Malmberg, H.-R.; Etheve, S.; Tuomainen, T.-P.; Virtanen, J.K.; Bendik, I.; Carlberg, C. Common and personal target genes of the micronutrient vitamin D in primary immune cells from human peripheral blood. Sci. Rep. 2020, 10, 21051. [Google Scholar] [CrossRef]
- Nurminen, V.; Seuter, S.; Carlberg, C. Primary Vitamin D Target Genes of Human Monocytes. Front. Physiol. 2019, 10, 194. [Google Scholar] [CrossRef]
- Wang, H.; Chen, W.; Li, D.; Yin, X.; Zhang, X.; Olsen, N.; Zheng, S.G. Vitamin D and Chronic Diseases. Aging Dis. 2017, 8, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; He, Y.; Shen, Y.; Zhang, Q.; Chen, D.; Zuo, C.; Qin, J.; Wang, H.; Wang, J.; Yu, Y. Vitamin D Inhibits COX-2 Expression and Inflammatory Response by Targeting Thioesterase Superfamily Member 4. J. Biol. Chem. 2014, 289, 11681–11694. [Google Scholar] [CrossRef] [PubMed]
- Scuto, M.; Salinaro, A.T.; Caligiuri, I.; Ontario, M.L.; Greco, V.; Sciuto, N.; Crea, R.; Calabrese, E.J.; Rizzolio, F.; Canzonieri, V.; et al. Redox modulation of vitagenes via plant polyphenols and vitamin D: Novel insights for chemoprevention and therapeutic interventions based on organoid technology. Mech. Ageing Dev. 2021, 199, 111551. [Google Scholar] [CrossRef]
- Yoon, B.R.; Oh, Y.-J.; Kang, S.W.; Lee, E.B.; Lee, W.-W. Role of SLC7A5 in Metabolic Reprogramming of Human Monocyte/Macrophage Immune Responses. Front. Immunol. 2018, 9, 53. [Google Scholar] [CrossRef]
- Crowe, W.; Elliott, C.T.; Green, B.D. A Review of the In Vivo Evidence Investigating the Role of Nitrite Exposure from Processed Meat Consumption in the Development of Colorectal Cancer. Nutrients 2019, 11, 2673. [Google Scholar] [CrossRef]
- Crowe, W.; Pan, X.; Mackle, J.; Harris, A.; Hardiman, G.; Elliott, C.T.; Green, B. Dietary inclusion of nitrite-containing frankfurter exacerbates colorectal cancer pathology and alters metabolism in APC (min) mice. NPJ Sci. Food 2022, 6, 60. [Google Scholar] [CrossRef] [PubMed]
- Laveti, D.; Kumar, M.; Hemalatha, R.; Sistla, R.; Naidu, V.; Talla, V.; Verma, V.; Kaur, N.; Nagpal, R. Anti-Inflammatory Treatments for Chronic Diseases: A Review. Inflamm. Allergy Drug Targets 2013, 12, 349–361. [Google Scholar] [CrossRef]
- Han, Y.; Jo, H.; Cho, J.H.; Dhanasekaran, D.N.; Song, Y.S. Resveratrol as a Tumor-Suppressive Nutraceutical Modulating Tumor Microenvironment and Malignant Behaviors of Cancer. Int. J. Mol. Sci. 2019, 20, 925. [Google Scholar] [CrossRef]
- Sajadimajd, S.; Khazaei, M. Oxidative Stress and Cancer: The Role of Nrf2. Curr. Cancer Drug Targets 2018, 18, 538–557. [Google Scholar] [CrossRef]
- Enkhbat, T.; Nishi, M.; Yoshikawa, K.; Jun, H.; Tokunaga, T.; Takasu, C.; Kashihara, H.; Ishikawa, D.; Tominaga, M.; Shimada, M. Epigallocatechin-3-gallate Enhances Radiation Sensitivity in Colorectal Cancer Cells Through Nrf2 Activation and Autophagy. Anticancer. Res. 2018, 38, 6247–6252. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.X.; Li, Y.Y.; Wang, N.; Luo, G.; Wang, J.; Luo, C.; Yu, W.; Hao, L. 1 alpha,25-Dihydroxyvitamin D-3 prevents renal oxidative damage via the PARP1/SIRT1/NOX4 pathway in Zucker diabetic fatty rats. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E343–E356. [Google Scholar] [CrossRef]
- Canto, C.; Auwerx, J. PGC-1alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr. Opin. Lipidol. 2009, 20, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Gerhart-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Puigserver, P.; Auwerx, J. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature 2009, 458, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-Y.; Kao, C.-L.; Ma, H.-I.; Hung, C.-H.; Wang, C.-T.; Liu, D.-H.; Chen, P.-Y.; Tsai, K.-L. SIRT1-related inhibition of pro-inflammatory responses and oxidative stress are involved in the mechanism of nonspecific low back pain relief after exercise through modulation of Toll-like receptor 4. J. Biochem. 2015, 158, 299–308. [Google Scholar] [CrossRef]
- Maya-Monteiro, C.M.; Bozza, P.T. Leptin and mTOR: Partners in metabolism and inflammation. Cell Cycle 2008, 7, 1713–1717. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Shaw, R.J. AMPK: Mechanisms of Cellular Energy Sensing and Restoration of Metabolic Balance. Mol. Cell 2017, 66, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-J.; Liaw, S.-F.; Lin, C.-C.; Chiu, C.-H.; Lin, M.-W.; Chang, F.-T. Effect of Nasal CPAP on SIRT1 and Endothelial Function in Obstructive Sleep Apnea Syndrome. Lung 2015, 193, 1037–1045. [Google Scholar] [CrossRef]
- Nakamaru, Y.; Vuppusetty, C.; Wada, H.; Milne, J.C.; Ito, M.; Rossios, C.; Elliot, M.; Hogg, J.; Kharitonov, S.; Goto, H.; et al. A protein deacetylase SIRT1 is a negative regulator of metalloproteinase-9. FASEB J. 2009, 23, 2810–2819. [Google Scholar] [CrossRef]
- Guo, C.; Sinnott, B.; Niu, B.; Lowry, M.B.; Fantacone, M.L.; Gombart, A.F. Synergistic induction of human cathelicidin antimicrobial peptide gene expression by vitamin D and stilbenoids. Mol. Nutr. Food Res. 2014, 58, 528–536. [Google Scholar] [CrossRef]
- Švajger, U.; Jeras, M. Anti-inflammatory Effects of Resveratrol and Its Potential Use in Therapy of Immune-mediated Diseases. Int. Rev. Immunol. 2012, 31, 202–222. [Google Scholar] [CrossRef] [PubMed]
- Guha, P.; Dey, A.; Sen, R.; Chatterjee, M.; Chattopadhyay, S.; Bandyopadhyay, S.K. Intracellular GSH Depletion Triggered Mitochondrial Bax Translocation to Accomplish Resveratrol-Induced Apoptosis in the U937 Cell Line. J. Pharmacol. Exp. Ther. 2011, 336, 206–214. [Google Scholar] [CrossRef]
- Ávila-Gálvez, M.Á.; García-Villalba, R.; Martínez-Díaz, F.; Ocaña-Castillo, B.; Monedero-Saiz, T.; Torrecillas-Sánchez, A.; Abellán, B.; González-Sarrías, A.; Espín, J.C. Metabolic Profiling of Dietary Polyphenols and Methylxanthines in Normal and Malignant Mammary Tissues from Breast Cancer Patients. Mol. Nutr. Food Res. 2019, 63, 1801239. [Google Scholar] [CrossRef]
- Kueck, A.; Opipari, A.W.; Griffith, K.A.; Tan, L.; Choi, M.; Huang, J.; Wahl, H.; Liu, J.R. Resveratrol inhibits glucose metabolism in human ovarian cancer cells. Gynecol. Oncol. 2007, 107, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Gwak, H.; Haegeman, G.; Tsang, B.K.; Song, Y.S. Cancer-specific interruption of glucose metabolism by resveratrol is mediated through inhibition of Akt/GLUT1 axis in ovarian cancer cells. Mol. Carcinog. 2015, 54, 1529–1540. [Google Scholar] [CrossRef] [PubMed]
- Turan, B.; Tuncay, E.; Vassort, G. Resveratrol and diabetic cardiac function: Focus on recent in vitro and in vivo studies. J. Bioenerg. Biomembr. 2012, 44, 281–296. [Google Scholar] [CrossRef]
- García-Zepeda, S.P.; García-Villa, E.; Díaz-Chávez, J.; Hernández-Pando, R.; Gariglio, P. Resveratrol induces cell death in cervical cancer cells through apoptosis and autophagy. Eur. J. Cancer Prev. 2013, 22, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Enríquez, S.; Pacheco-Velázquez, S.C.; Marín-Hernández, M.; Gallardo-Pérez, J.C.; Robledo-Cadena, D.X.; Hernández-Reséndiz, I.; García-García, J.D.; Belmont-Díaz, J.; López-Marure, R.; Hernández-Esquivel, L.; et al. Resveratrol inhibits cancer cell proliferation by impairing oxidative phosphorylation and inducing oxidative stress. Toxicol. Appl. Pharmacol. 2019, 370, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Richard, T.; Pawlus, A.D.; Iglésias, M.-L.; Pedrot, E.; Waffo-Teguo, P.; Mérillon, J.-M.; Monti, J.-P. Neuroprotective properties of resveratrol and derivatives. Ann. N. Y. Acad. Sci. 2011, 1215, 103–108. [Google Scholar] [CrossRef]
- Duez, H.; Staels, B. Rev-erb-α: An integrator of circadian rhythms and metabolism. J. Appl. Physiol. 2009, 107, 1972–1980. [Google Scholar] [CrossRef]
- Ch, R.; Chevallier, O.; Elliott, C.T. Metabolomics reveal circadian control of cellular metabolism. TrAC Trends Anal. Chem. 2020, 130, 115986. [Google Scholar] [CrossRef]
- Takahashi, J.S.; Hong, H.-K.; Ko, C.H.; McDearmon, E.L. The genetics of mammalian circadian order and disorder: Implications for physiology and disease. Nat. Rev. Genet. 2008, 9, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Eckel-Mahan, K.; Sassone-Corsi, P. Metabolism and the Circadian Clock Converge. Physiol. Rev. 2013, 93, 107–135. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Sundar, I.K.; Yao, H.; Sellix, M.T.; Rahman, I. Circadian clock function is disrupted by environmental tobacco/cigarette smoke, leading to lung inflammation and injury via a SIRT1-BMAL1 pathway. FASEB J. 2014, 28, 176–194. [Google Scholar] [CrossRef]
- Chang, H.-C.; Guarente, L. SIRT1 and other sirtuins in metabolism. Trends Endocrinol. Metab. 2014, 25, 138–145. [Google Scholar] [CrossRef]
- Hewison, M.; Adams, J.S. Chapter 45—Extrarenal 1α-Hydroxylase. In Vitamin D, 3rd ed.; Feldman, D., Pike, J.W., Adams, J.S., Eds.; Academic Press: San Diego, CA, USA, 2011; pp. 777–804. [Google Scholar]
- Gray, R.W.; Caldas, A.E.; Wilz, D.R.; Lemann, J.; Smith, G.A.; DeLuca, H.F. Metabolism and excretion of 3H-1,25-(OH)2-vitamin D3 in healthy adults. J. Clin. Endocrinol. Metab. 1978, 46, 756–765. [Google Scholar] [CrossRef] [PubMed]
- Seeman, E.; Kumar, R.; Hunder, G.G.; Scott, M.; Heath, H.; Riggs, B.L. Production, degradation, and circulating levels of 1,25-dihydroxyvitamin D in health and in chronic glucocorticoid excess. J. Clin. Investig. 1980, 66, 664–669. [Google Scholar] [CrossRef]
- Wiesner, R.H.; Kumar, R.; Seeman, E.; Go, V.L. Enterohepatic physiology of 1,25-dihydroxyvitamin D3 metabolites in normal man. J. Lab. Clin. Med. 1980, 96, 1094–1100. [Google Scholar]
- Harnden, D.; Kumar, R.; Holick, M.F.; DeLuca, H.F. Side Chain Metabolism of 25-Hydroxy-[26,27−14C] Vitamin D3 and 1,25-Dihydroxy-[26,27−14C] Vitamin D3 in Vivo. Science 1976, 193, 493–494. [Google Scholar] [CrossRef]
- Demers, C.; Lemay, J.; Hendy, G.N.; Gascon-Barré, M. Comparative in vivo expression of the calcitriol-24-hydroxylase gene in kidney and intestine. J. Mol. Endocrinol. 1997, 18, 37–48. [Google Scholar] [CrossRef]
- Furuichi, T.; Kawata, S.; Asoh, Y.; Kumaki, K.; Ohyama, Y. Differential time course of induction of 1alpha,25-dihydroxyvitamin D3-24-hydroxylase mRNA expression in rats by 1alpha,25-dihydroxyvitamin D3 and its analogs. Life Sci. 1998, 62, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Knutson, J.C.; DeLuca, H.F. 25-Hydroxyvitamin D3-24-hydroxylase. Subcellular location and properties. Biochemistry 1974, 13, 1543–1548. [Google Scholar] [CrossRef] [PubMed]
- Omdahl, J.L.; Morris, H.A.; May, B.K. Hydroxylase enzymes of the vitamin d pathway: Expression, function, and regulation. Annu. Rev. Nutr. 2002, 22, 139–166. [Google Scholar] [CrossRef] [PubMed]
- Petkovich, M.; Helvig, C.; Epps, T. Chapter 80—CYP24A1 Regulation in Health and Disease. In Vitamin D, 3rd ed.; Feldman, D., Pike, J.W., Adams, J.S., Eds.; Academic Press: San Diego, CA, USA, 2011; pp. 1525–1554. [Google Scholar]
- King, A.N.; Beer, D.G.; Christensen, P.J.; Simpson, R.U.; Ramnath, N. The Vitamin D/CYP24A1 Story in Cancer. Anti Cancer Agents Med. Chem. 2010, 10, 213–224. [Google Scholar] [CrossRef]
- Horváth, H.C.; Lakatos, P.; Kósa, J.P.; Bácsi, K.; Borka, K.; Bises, G.; Nittke, T.; Hershberger, P.A.; Speer, G.; Kállay, E. The Candidate Oncogene CYP24A1: A Potential Biomarker for Colorectal Tumorigenesis. J. Histochem. Cytochem. 2010, 58, 277–285. [Google Scholar] [CrossRef]
- Tenenhouse, H.S.; Scriver, C.R. X-linked hypophosphatemia. A phenotype in search of a cause. Int. J. Biochem. 1992, 24, 685–691. [Google Scholar] [CrossRef]
- Roy, S.; Martel, J.; Ma, S.; Tenenhouse, H.S. Increased renal 25-hydroxyvitamin D3-24-hydroxylase messenger ribonucleic acid and immunoreactive protein in phosphate-deprived Hyp mice: A mechanism for accelerated 1,25-dihydroxyvitamin D3 catabolism in X-linked hypophosphatemic rickets. Endocrinology 1994, 134, 1761–1767. [Google Scholar] [CrossRef]
- Hough, S.; Fausto, A.; Sonn, Y.; Jo, O.K.D.; Birge, S.J.; Avioli, L.V. Vitamin D Metabolism in the Chronic Streptozotocin-Induced Diabetic Rat*. Endocrinology 1983, 113, 790–796. [Google Scholar] [CrossRef]
- Rao, L.G.; Kung, M.S. Effect of long-term and short-term diabetes on the parathyroid hormone sensitive rat renal adenylate cyclase: Correlation with vitamin D metabolism. Can. J. Physiol. Pharmacol. 1988, 66, 1313–1318. [Google Scholar] [CrossRef]
- Zehnder, D.; Quinkler, M.; Eardley, K.S.; Bland, R.; Lepenies, J.; Hughes, S.V.; Raymond, N.T.; Howie, A.J.; Cockwell, P.; Stewart, P.M.; et al. Reduction of the vitamin D hormonal system in kidney disease is associated with increased renal inflammation. Kidney Int. 2008, 74, 1343–1353. [Google Scholar] [CrossRef]
- Helvig, C.F.; Cuerrier, D.; Hosfield, C.M.; Ireland, B.; Kharebov, A.Z.; Kim, J.W.; Ramjit, N.J.; Ryder, K.; Tabash, S.P.; Herzenberg, A.M.; et al. Dysregulation of renal vitamin D metabolism in the uremic rat. Kidney Int. 2010, 78, 463–472. [Google Scholar] [CrossRef]
- Holick, M.F. Vitamin D deficiency. N. Engl. J. Med. 2007, 357, 266–281. [Google Scholar] [CrossRef]
- Bovolini, A.; Garcia, J.; Andrade, M.A.; Duarte, J.A. Metabolic Syndrome Pathophysiology and Predisposing Factors. Int. J. Sports Med. 2021, 42, 199–214. [Google Scholar] [CrossRef]
- Litwin, M.; Kulaga, Z. Obesity, metabolic syndrome, and primary hypertension. Pediatr. Nephrol. 2021, 36, 825–837. [Google Scholar] [CrossRef]
- Karra, P.; Winn, M.; Pauleck, S.; Bulsiewicz-Jacobsen, A.; Peterson, L.; Coletta, A.; Doherty, J.; Ulrich, C.M.; Summers, S.A.; Gunter, M.; et al. Metabolic dysfunction and obesity-related cancer: Beyond obesity and metabolic syndrome. Obesity 2022, 30, 1323–1334. [Google Scholar] [CrossRef]
- Chasens, E.R.; Imes, C.C.; Kariuki, J.K.; Luyster, F.S.; Morris, J.L.; DiNardo, M.M.; Godzik, M.; Jeon, B.; Yang, K. Sleep and Metabolic Syndrome. Nurs. Clin. N. Am. 2021, 56, 203–217. [Google Scholar] [CrossRef]
- Więckowska-Gacek, A.; Mietelska-Porowska, A.; Wydrych, M.; Wojda, U. Western diet as a trigger of Alzheimer’s disease: From metabolic syndrome and systemic inflammation to neuroinflammation and neurodegeneration. Ageing Res. Rev. 2021, 70, 101397. [Google Scholar] [CrossRef]
- Kouvari, M.; D’Cunha, N.M.; Travica, N.; Sergi, D.; Zec, M.; Marx, W.; Naumovski, N. Metabolic Syndrome, Cognitive Impairment and the Role of Diet: A Narrative Review. Nutrients 2022, 14, 333. [Google Scholar] [CrossRef]
- Harinarayan, C.V. Vitamin D and diabetes mellitus. Hormones 2014, 13, 163–181. [Google Scholar] [CrossRef]
- Lim, S.; Kim, M.J.; Choi, S.H.; Shin, C.S.; Park, K.S.; Jang, H.C.; Billings, L.K.; Meigs, J.B. Association of vitamin D deficiency with incidence of type 2 diabetes in high-risk Asian subjects. Am. J. Clin. Nutr. 2013, 97, 524–530. [Google Scholar] [CrossRef]
- Al-Timimi, D.J.; Ali, A.F. Serum 25(OH) D in Diabetes Mellitus Type 2: Relation to Glycaemic Control. J. Clin. Diagn. Res. 2013, 7, 2686–2688. [Google Scholar] [CrossRef]
- Theik, N.W.Y.; Raji, O.E.; Shenwai, P.; Shah, R.; Kalluri, S.R.; Bhutta, T.H.; Hannoodee, H.; Al Khalili, M.; Khan, S. Relationship and Effects of Vitamin D on Metabolic Syndrome: A Systematic Review. Cureus 2021, 13, e17419. [Google Scholar] [CrossRef]
- Latic, N.; Erben, R.G. Vitamin D and Cardiovascular Disease, with Emphasis on Hypertension, Atherosclerosis, and Heart Failure. Int. J. Mol. Sci. 2020, 21, 6483. [Google Scholar] [CrossRef]
- Cosentino, N.; Campodonico, J.; Milazzo, V.; De Metrio, M.; Brambilla, M.; Camera, M.; Marenzi, G. Vitamin D and Cardiovascular Disease: Current Evidence and Future Perspectives. Nutrients 2021, 13, 3603. [Google Scholar] [CrossRef]
- Lavie, C.J.; DiNicolantonio, J.J.; Milani, R.V.; O’Keefe, J.H. Vitamin D and Cardiovascular Health. Circulation 2013, 128, 2404–2406. [Google Scholar] [CrossRef]
- Zittermann, A.; Trummer, C.; Theiler-Schwetz, V.; Lerchbaum, E.; März, W.; Pilz, S. Vitamin D and Cardiovascular Disease: An Updated Narrative Review. Int. J. Mol. Sci. 2021, 22, 2896. [Google Scholar] [CrossRef]
- Pilz, S.; Trummer, C.; Theiler-Schwetz, V.; Grübler, M.R.; Verheyen, N.D.; Odler, B.; Karras, S.N.; Zittermann, A.; März, W. Critical Appraisal of Large Vitamin D Randomized Controlled Trials. Nutrients 2022, 14, 303. [Google Scholar] [CrossRef]
- Zhou, A.; Selvanayagam, J.B.; Hyppönen, E. Non-linear Mendelian randomization analyses support a role for vitamin D deficiency in cardiovascular disease risk. Eur. Hearth J. 2022, 43, 1731–1739. [Google Scholar] [CrossRef]
- Timmers, S.; Konings, E.; Bilet, L.; Houtkooper, R.H.; van de Weijer, T.; Goossens, G.H.; Hoeks, J.; van der Krieken, S.; Ryu, D.; Kersten, S.; et al. Calorie Restriction-like Effects of 30 Days of Resveratrol Supplementation on Energy Metabolism and Metabolic Profile in Obese Humans. Cell Metab. 2011, 14, 612–622. [Google Scholar] [CrossRef]
- Parsamanesh, N.; Asghari, A.; Sardari, S.; Tasbandi, A.; Jamialahmadi, T.; Xu, S.; Sahebkar, A. Resveratrol and endothelial function: A literature review. Pharmacol. Res. 2021, 170, 105725. [Google Scholar] [CrossRef]
- Dusso, A.S.; Slatopolsky, E. Chapter 70—Vitamin D and Renal Disease. In Vitamin D, 3rd ed.; Feldman, D., Pike, J.W., Adams, J.S., Eds.; Academic Press: San Diego, CA, USA, 2011; pp. 1325–1357. [Google Scholar]
- Dusso, A.S.; Brown, A.J.; Slatopolsky, E. Vitamin D. Am. J. Physiol. Renal. Physiol. 2005, 289, F8–F28. [Google Scholar] [CrossRef] [PubMed]
- Nykjaer, A.; Dragun, D.; Walther, D.; Vorum, H.; Jacobsen, C.; Herz, J.; Melsen, F.; Christensen, E.I.; E Willnow, T. An Endocytic Pathway Essential for Renal Uptake and Activation of the Steroid 25-(OH) Vitamin D3. Cell 1999, 96, 507–515. [Google Scholar] [CrossRef]
- Usatii, M.; Rousseau, L.; Demers, C.; Petit, J.-L.; Brossard, J.-H.; Gascon-Barré, M.; Lavigne, J.; Zahradnik, R.; Nemeth, E.; D’Amour, P. Parathyroid hormone fragments inhibit active hormone and hypocalcemia-induced 1,25(OH)2D synthesis. Kidney Int. 2007, 72, 1330–1335. [Google Scholar] [CrossRef] [PubMed]
- Al-Badr, W.; Martin, K.J. Vitamin D and Kidney Disease. Clin. J. Am. Soc. Nephrol. 2008, 3, 1555–1560. [Google Scholar] [CrossRef]
- Al-Aly, Z.; Qazi, R.A.; González, E.A.; Zeringue, A.; Martin, K.J. Changes in Serum 25-Hydroxyvitamin D and Plasma Intact PTH Levels Following Treatment with Ergocalciferol in Patients With CKD. Am. J. Kidney Dis. 2007, 50, 59–68. [Google Scholar] [CrossRef]
- Liu, W.; Yu, W.R.; Carling, T.; Juhlin, C.; Rastad, J.; Ridefelt, P.; Akerström, G.; Hellman, P. Regulation of gp330/megalin expression by vitamins A and D. Eur. J. Clin. Investig. 1998, 28, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Wang, X.; Li, L.; Huang, Z.; Fan, Y.; Song, T.; Lin, T. Prevalence of vitamin D deficiency and impact on clinical outcomes after kidney transplantation: A systematic review and meta-analysis. Nutr. Rev. 2022, 80, 950–961. [Google Scholar] [CrossRef] [PubMed]
- Stein, E.M.; Shane, E. Chapter 68—Vitamin D and Organ Transplantation. In Vitamin D, 3rd ed.; Feldman, D., Pike, J.W., Adams, J.S., Eds.; Academic Press: San Diego, CA, USA, 2011; pp. 1291–1298. [Google Scholar]
- Guichelaar, M.M.J.; Schmoll, J.; Malinchoc, M.; Hay, J.E. Fractures and avascular necrosis before and after orthotopic liver transplantation: Long-term follow-up and predictive factors. Hepatology 2007, 46, 1198–1207. [Google Scholar] [CrossRef]
- Oufroukhi, L.; Kamar, N.; Muscari, F.; Lavayssière, L.; Guitard, J.; Ribes, D.; Esposito, L.; Alric, L.; Hanaire, H.; Rostaing, L. Predictive Factors for Posttransplant Diabetes Mellitus Within One-Year of Liver Transplantation. Transplantation 2008, 85, 1436–1442. [Google Scholar] [CrossRef]
- Fernández-Ruiz, M.; Rodríguez-Goncer, I.; Ruiz-Merlo, T.; Parra, P.; López-Medrano, F.; Andrés, A.; Aguado, J.M. Low 25-hydroxyvitamin D Levels and the Risk of Late CMV Infection After Kidney Transplantation: Role for CMV-specific Mediated Immunity. Transplantation 2019, 103, e216–e217. [Google Scholar] [CrossRef]
- Kalluri, H.V.; Sacha, L.M.; Ingemi, A.; Shullo, M.A.; Johnson, H.J.; Sood, P.; Tevar, A.D.; Humar, A.; Venkataramanan, R. Low vitamin D exposure is associated with higher risk of infection in renal transplant recipients. Clin. Transplant. 2017, 31, e12955. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.E.; Kim, H.; Oh, H.J.; Park, J.T.; Han, S.H.; Ryu, D.-R.; Yoo, T.-H.; Kang, S.-W. Vitamin D Deficiency Is an Independent Risk Factor for Urinary Tract Infections After Renal Transplants. Medicine 2015, 94, e594. [Google Scholar] [CrossRef]
- Lowery, E.M.; Bemiss, B.; Cascino, T.; Durazo-Arvizu, R.A.; Forsythe, S.M.; Alex, C.; Laghi, F.; Love, R.B.; Camacho, P. Low vitamin D levels are associated with increased rejection and infections after lung transplantation. J. Heart Lung Transplant. 2012, 31, 700–707. [Google Scholar] [CrossRef] [PubMed]
- van Etten, E.; Mathieu, C. Immunoregulation by 1,25-dihydroxyvitamin D3: Basic concepts. J. Steroid Biochem. Mol. Biol. 2005, 97, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Zhang, Z.; Musch, M.W.; Ning, G.; Sun, J.; Hart, J.; Bissonnette, M.; Li, Y.C. Novel role of the vitamin D receptor in maintaining the integrity of the intestinal mucosal barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G208–G216. [Google Scholar] [CrossRef]
- Lagishetty, V.; Misharin, A.V.; Liu, N.Q.; Lisse, T.S.; Chun, R.F.; Ouyang, Y.; McLachlan, M.; Adams, S.; Hewison, M. Vitamin D deficiency in mice impairs colonic antibacterial activity and predisposes to colitis. Endocrinology 2010, 151, 2423–2432. [Google Scholar] [CrossRef]
- Meeker, S.; Seamons, A.; Paik, J.; Treuting, P.M.; Brabb, T.; Grady, W.M.; Maggio-Prince, L. Increased Dietary Vitamin D Suppresses MAPK Signaling, Colitis, and Colon Cancer. Cancer Res. 2014, 74, 4398–4408. [Google Scholar] [CrossRef]
- Speer, G. Impact of vitamin D in neurological diseases and neurorehabilitation: From dementia to multiple sclerosis. Part I: The role of vitamin D in the prevention and treatment of multiple sclerosis. Ideggyogy Sz. 2013, 66, 293–303. [Google Scholar]
- de Carvalho, J.F.; Pereira, R.M.; Shoenfeld, Y. Pearls in autoimmunity. Auto Immun. Highlights 2011, 2, 1–4. [Google Scholar] [CrossRef]
- van Etten, E.; Gysemans, C.; Branisteanu, D.D.; Verstuyf, A.; Bouillon, R.; Overbergh, L.; Mathieu, C. Novel insights in the immune function of the vitamin D system: Synergism with interferon-beta. J. Steroid Biochem. Mol. Biol. 2007, 103, 546–551. [Google Scholar] [CrossRef]
- Gregori, S.; Giarratana, N.; Smiroldo, S.; Uskokovic, M.; Adorini, L. A 1α,25-Dihydroxyvitamin D3 Analog Enhances Regulatory T-Cells and Arrests Autoimmune Diabetes in NOD Mice. Diabetes 2002, 51, 1367–1374. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, C.; Waer, M.; Laureys, J.; Rutgeerts, O.; Bouillon, R. Prevention of autoimmune diabetes in NOD mice by 1,25 dihydroxyvitamin D3. Diabetologia 1994, 37, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Takiishi, T.; Ding, L.; Baeke, F.; Spagnuolo, I.; Sebastiani, G.; Laureys, J.; Verstuyf, A.; Carmeliet, G.; Dotta, F.; Van Belle, T.L.; et al. Dietary Supplementation with High Doses of Regular Vitamin D3 Safely Reduces Diabetes Incidence in NOD Mice When Given Early and Long Term. Diabetes 2014, 63, 2026–2036. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.; Khandker, S.S.; Alam, S.S.; Kotyla, P.; Hassan, R. Vitamin D status in patients with systemic lupus erythematosus (SLE): A systematic review and meta-analysis. Autoimmun. Rev. 2019, 18, 102392. [Google Scholar] [CrossRef]
- Wang, M.; Liu, M.; Wang, C.; Xiao, Y.; An, T.; Zou, M.; Cheng, G. Association between vitamin D status and asthma control: A meta-analysis of randomized trials. Respir. Med. 2019, 150, 85–94. [Google Scholar] [CrossRef]
- Mitri, J.; Pittas, A.G. Vitamin D and diabetes. Endocrinol. Metab. Clin. N. Am. 2014, 43, 205–232. [Google Scholar] [CrossRef]
- Grammatiki, M.; Rapti, E.; Karras, S.; Ajjan, R.A.; Kotsa, K. Vitamin D and diabetes mellitus: Causal or casual association? Rev. Endocr. Metab. Disord. 2017, 18, 227–241. [Google Scholar] [CrossRef]
- Maestro, B.; Campion, J.; Dávila, N.; Calle, C. Stimulation by 1,25-Dihydroxyvitamin D3 of Insulin Receptor Expression and Insulin Responsiveness for Glucose Transport in U-937 Human Promonocytic Cells. Endocr. J. 2000, 47, 383–391. [Google Scholar] [CrossRef]
- Pittas, A.G.; Dawson-Hughes, B.; Sheehan, P.; Ware, J.H.; Knowler, W.C.; Aroda, V.R.; Brodsky, I.; Ceglia, L.; Chadha, C.; Chatterjee, R.; et al. Vitamin D Supplementation and Prevention of Type 2 Diabetes. N. Engl. J. Med. 2019, 381, 520–530. [Google Scholar] [CrossRef]
- Sacerdote, A.; Dave, P.; Lokshin, V.; Bahtiyar, G. Type 2 Diabetes Mellitus, Insulin Resistance, and Vitamin D. Curr. Diab. Rep. 2019, 19, 101. [Google Scholar] [CrossRef]
- Jeffery, L.E.; Henley, P.; Marium, N.; Filer, A.; Sansom, D.M.; Hewison, M.; Raza, K. Decreased sensitivity to 1,25-dihydroxyvitamin D3 in T cells from the rheumatoid joint. J. Autoimmun. 2018, 88, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.H.; Jeffery, L.E.; Soskic, B.; Briggs, Z.; Hou, T.Z.; Raza, K.; Sansom, D.M. 1,25(OH)2D3 Promotes the Efficacy of CD28 Costimulation Blockade by Abatacept. J. Immunol. 2015, 195, 2657–2665. [Google Scholar] [CrossRef] [PubMed]
- Charoenngam, N. Vitamin D and Rheumatic Diseases: A Review of Clinical Evidence. Int. J. Mol. Sci. 2021, 22, 10659. [Google Scholar] [CrossRef] [PubMed]
- Na, S.-Y.; Kim, K.B.; Lim, Y.J.; Song, H.J. Vitamin D and Colorectal Cancer: Current Perspectives and Future Directions. J. Cancer Prev. 2022, 27, 147–156. [Google Scholar] [CrossRef]
- Trump, D.L. Calcitriol and cancer therapy: A missed opportunity. Bone Rep. 2018, 9, 110–119. [Google Scholar] [CrossRef]
- Watanabe, R.; Inoue, D. Current Topics on Vitamin D. Anti-cancer effects of vitamin D. Clin. Calcium. 2015, 25, 373–380. [Google Scholar]
- Giammanco, M.; Di Majo, D.; La Guardia, M.; Aiello, S.; Crescimannno, M.; Flandina, C.; Tumminello, F.M.; Leto, G. Vitamin D in cancer chemoprevention. Pharm. Biol. 2015, 53, 1399–1434. [Google Scholar] [CrossRef]
- Ramnath, N.; Nadal, E.; Jeon, C.K.; Sandoval, J.; Colacino, J.; Rozek, L.S.; Christensen, P.J.; Esteller, M.; Beer, D.G.; Kim, S.H. Epigenetic Regulation of Vitamin D Metabolism in Human Lung Adenocarcinoma. J. Thorac. Oncol. 2014, 9, 473–482. [Google Scholar] [CrossRef]
- Scaglione-Sewell, B.A.; Bissonnette, M.; Skarosi, S.; Abraham, C.; Brasitus, T.A. A vitamin D3 analog induces a G1-phase arrest in CaCo-2 cells by inhibiting cdk2 and cdk6: Roles of cyclin E, p21Waf1, and p27Kip1. Endocrinology 2000, 141, 3931–3939. [Google Scholar] [CrossRef]
- Razak, S.; Afsar, T.; Almajwal, A.; Alam, I.; Jahan, S. Growth inhibition and apoptosis in colorectal cancer cells induced by Vitamin D-Nanoemulsion (NVD): Involvement of Wnt/beta-catenin and other signal transduction pathways. Cell Biosci. 2019, 9, 15. [Google Scholar] [CrossRef]
- Beildeck, M.E.; Islam, M.; Shah, S.; Welsh, J.; Byers, S.W. Control of TCF-4 Expression by VDR and Vitamin D in the Mouse Mammary Gland and Colorectal Cancer Cell Lines. PLoS ONE 2009, 4, e7872. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, A.; Grant, W.B. Vitamin D and Cancer: An Historical Overview of the Epidemiology and Mechanisms. Nutrients 2022, 14, 1448. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Jiang, C.; Cong, L.; Wu, N.; Wang, X.; Hao, M.; Liu, T.; Wang, L.; Liu, Y.; Cong, X. CYP24A1 Inhibition Facilitates the Antiproliferative Effect of 1,25(OH)(2)D(3) Through Downregulation of the WNT/beta-Catenin Pathway and Methylation-Mediated Regulation of CYP24A1 in Colorectal Cancer Cells. DNA Cell Biol. 2018, 37, 742–749. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, A.; Malki, A. Vitamin D Signaling in Inflammation and Cancer: Molecular Mechanisms and Therapeutic Implications. Molecules 2020, 25, 3219. [Google Scholar] [CrossRef]
- The Human Protein Atlas: VDR. Available online: http://www.proteinatlas.org/ENSG00000111424-VDR/tissue (accessed on 19 August 2022).
- Alkhalaf, M. Resveratrol-Induced Apoptosis Is Associated with Activation of p53 and Inhibition of Protein Translation in T47D Human Breast Cancer Cells. Pharmacology 2007, 80, 134–143. [Google Scholar] [CrossRef]
- Singh, B.; Shoulson, R.; Chatterjee, A.; Ronghe, A.; Bhat, N.K.; Dim, D.C.; Bhat, H.K. Resveratrol inhibits estrogen-induced breast carcinogenesis through induction of NRF2-mediated protective pathways. Carcinogenesis 2014, 35, 1872–1880. [Google Scholar] [CrossRef]
- Zhou, X.; Zhao, Y.; Wang, J.; Wang, X.; Chen, C.; Yin, D.; Zhao, F.; Yin, J.; Guo, M.; Zhang, L.; et al. Resveratrol represses estrogen-induced mammary carcinogenesis through NRF2-UGT1A8-estrogen metabolic axis activation. Biochem. Pharmacol. 2018, 155, 252–263. [Google Scholar] [CrossRef]
- Jin, X.; Wei, Y.; Liu, Y.; Lu, X.; Ding, F.; Wang, J.; Yang, S. Resveratrol promotes sensitization to Doxorubicin by inhibiting epithelial-mesenchymal transition and modulating SIRT1/beta-catenin signaling pathway in breast cancer. Cancer Med. 2019, 8, 1246–1257. [Google Scholar] [CrossRef]
- An, B.-S.; Tavera-Mendoza, L.E.; Dimitrov, V.; Wang, X.; Calderon, M.R.; Wang, H.-J.; White, J.H. Stimulation of Sirt1-Regulated FoxO Protein Function by the Ligand-Bound Vitamin D Receptor. Mol. Cell. Biol. 2010, 30, 4890–4900. [Google Scholar] [CrossRef]
- An, B.; Tavera-Mendoza, L.; Dimitrov, V.; White, J. Stimulation of Sirt1-Regulated FoxO Protein Function by the Hormone-Bound Vitamin D Receptor. In Endocrine Reviews; Endocrine Society: Chevy Chase, MD, USA, 2010; Volume 31. [Google Scholar]
- Chen, H.Y.; Hu, X.Q.; Yang, R.L.; Wu, G.P.; Tan, Q.; Goltzman, D.; Miao, D. SIRT1/FOXO3a axis plays an important role in the prevention of mandibular bone loss induced by 1,25(OH)(2)D deficiency. Int. J. Biol. Sci. 2020, 16, 2712–2726. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tong, L.; Luo, Y.; Li, X.; Chen, G.; Wang, Y. Resveratrol inhibits the proliferation and induces the apoptosis in ovarian cancer cells via inhibiting glycolysis and targeting AMPK/mTOR signaling pathway. J. Cell. Biochem. 2018, 119, 6162–6172. [Google Scholar] [CrossRef] [PubMed]
- Gweon, E.J.; Kim, S.-J. Resveratrol attenuates matrix metalloproteinase-9 and -2-regulated differentiation of HTB94 chondrosarcoma cells through the p38 kinase and JNK pathways. Oncol. Rep. 2014, 32, 71–78. [Google Scholar] [CrossRef]
- Chen, L.; Liu, T.; Wang, Q.; Liu, J. Anti-inflammatory effect of combined tetramethylpyrazine, resveratrol and curcumin in vivo. BMC Complement. Altern. Med. 2017, 17, 233. [Google Scholar] [CrossRef]
- Qi, B.; Shi, C.; Meng, J.; Xu, S.; Liu, J. Resveratrol alleviates ethanol-induced neuroinflammation in vivo and in vitro: Involvement of TLR2-MyD88-NF-κB pathway. Int. J. Biochem. Cell Biol. 2018, 103, 56–64. [Google Scholar]
- Patel, K.R.; Brown, V.A.; Jones, D.J.; Britton, R.G.; Hemingway, D.; Miller, A.S.; West, K.P.; Booth, T.D.; Perloff, M.; Crowell, J.A.; et al. Clinical Pharmacology of Resveratrol and Its Metabolites in Colorectal Cancer Patients. Cancer Res. 2010, 70, 7392–7399. [Google Scholar] [CrossRef]
- Chung, S.S.; Dutta, P.; Austin, D.; Wang, P.; Awad, A.; Vadgama, J.V. Combination of resveratrol and 5-flurouracil enhanced anti-telomerase activity and apoptosis by inhibiting STAT3 and Akt signaling pathways in human colorectal cancer cells. Oncotarget 2018, 9, 32943–32957. [Google Scholar] [CrossRef]
- Manna, P.; Achari, A.E.; Jain, S.K. 1,25(OH)(2)-vitamin D-3 upregulates glucose uptake mediated by SIRT1/IRS1/GLUT4 signaling cascade in C2C12 myotubes. Mol. Cell. Biochem. 2018, 444, 103–108. [Google Scholar] [CrossRef]
- Qu, H.; Lin, K.; Wang, H.; Wei, H.L.; Ji, B.L.; Yang, Z.S.; Peng, C.; Xiao, X.; Deng, H. 1,25(OH)(2)D-3 improves cardiac dysfunction, hypertrophy, and fibrosis through PARP1/SIRT1/mTOR-related mechanisms in type 1 diabetes. Mol. Nutr. Food Res. 2017, 61, 1600338. [Google Scholar] [CrossRef]
- Safarpour, P.; Daneshi-Maskooni, M.; Vafa, M.; Nourbakhsh, M.; Janani, L.; Maddah, M.; Amiri, F.-S.; Mohammadi, F.; Sadeghi, H. Vitamin D supplementation improves SIRT1, Irisin, and glucose indices in overweight or obese type 2 diabetic patients: A double-blind randomized placebo-controlled clinical trial. BMC Fam. Pract. 2020, 21, 26. [Google Scholar] [CrossRef]
- Sabir, M.S.; Khan, Z.; Hu, C.C.; Galligan, M.A.; Dussik, C.M.; Mallick, S.; Stone, A.D.; Batie, S.F.; Jacobs, E.T.; Whitfield, G.K.; et al. SIRT1 enzymatically potentiates 1,25-dihydroxyvitamin D-3 signaling via vitamin D receptor deacetylation. J. Steroid Biochem. Mol. Biol. 2017, 172, 117–129. [Google Scholar] [PubMed]
- Glosse, P.; Feger, M.; Mutig, K.; Chen, H.; Hirche, F.; Hasan, A.A.; Gaballa, M.; Hocher, B.; Lang, F.; Föller, M. AMP-activated kinase is a regulator of fibroblast growth factor 23 production. Kidney Int. 2018, 94, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Stone, A.; Batie, S.F.; Sabir, M.S.; Jacobs, E.T.; Lee, J.H.; Whitfield, G.K.; Haussler, M.R.; Jurutka, P.W. Resveratrol potentiates vitamin D and nuclear receptor signaling. J. Cell Biochem. 2015, 116, 1130–1143. [Google Scholar]
- Feigerlova, E.; Demarquet, L.; Melhem, H.; Ghemrawi, R.; Battaglia-Hsu, S.F.; Ewu, E.; Alberto, J.M.; Helle, D.; Weryha, G.; Guéant, J.L. Methyl donor deficiency impairs bone development via peroxisome proliferator-activated receptor-gamma coactivator-1alpha-dependent vitamin D receptor pathway. FASEB J. 2016, 30, 3598–35612. [Google Scholar] [CrossRef] [PubMed]
- Marampon, F.; Gravina, G.; Festuccia, C.; Colapietro, A.; Di Cesare, E.; Tombolini, V. EP-2041: Vitamin D protects HUVEC from RT-induced senescence and apoptosis by modulating MAPK/SirT1 axis. Radiother. Oncol. 2016, 119, S963–S964. [Google Scholar] [CrossRef]
- Marampon, F.; Gravina, G.L.; Festuccia, C.; Popov, V.M.; Colapietro, A.; Sanita, P.; Musio, D.; De Felice, F.; Lenzi, A.; Jannini, E.A.; et al. Vitamin D protects endothelial cells from irradiation-induced senescence and apoptosis by modulating MAPK/SirT1 axis. J. Endocrinol. Investig. 2020, 43, 411–422. [Google Scholar] [CrossRef]
- Institute of Medicine (US) Committee; Ross, A.C.; Taylor, C.L.; Yaktine, A.L.; Del Valle, H.B. (Eds.) Dietary Reference Intakes for Calcium and Vitamin D; The National Academies Collection; National Academic Press: Washington, DC, USA, 2011. [Google Scholar]
- Institute of Medicine (US) Committee; Ross, A.C.; Taylor, C.L.; Yaktine, A.L.; Del Valle, H.B. (Eds.) Tolerable Upper Intake Levels: Calcium and Vitamin D. In Dietary Reference Intakes for Calcium and Vitamin D; The National Academies Collection; National Academic Press: Washington, DC, USA, 2011. [Google Scholar]
- Chauhan, K.; Shahrokhi, M.; Huecker, M. Vitamin D; [Updated 2023 Marc 3]. In StatPearls [Internet]; StatPerls Publishing: Treasure Island, FL, USA, January 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK441912/ (accessed on 16 January 2023).
- Pludowski, P.; Takacs, I.; Boyanov, M.; Belaya, Z.; Diaconu, C.C.; Mokhort, T.; Zherdova, N.; Rasa, I.; Payer, J.; Pilz, S. Clinical Practice in the Prevention, Diagnosis and Treatment of Vitamin D Deficiency: A Central and Eastern European Expert Consensus Statement. Nutrients 2022, 14, 1483. [Google Scholar] [CrossRef]
- Takacs, I.; Dank, M.; Majnik, J.; Nagy, G.; Szabo, A.; Szabo, B.; Zoltan, S.; Istvan, S.; Erzsebet, T.; Andras, T.; et al. Hungarian consensus recommendation on the role of vitamin D in disease prevention and treatment. Orv. Hetil. 2022, 163, 575–584. [Google Scholar]
- Kuznia, S.; Czock, D.; Kopp-Schneider, A.; Caspari, R.; Fischer, H.; Laetsch, D.C.; Slavic, M.; Brenner, H.; Schöttker, B. Efficacy and Safety of a Personalized Vitamin D3 Loading Dose Followed by Daily 2000 IU in Colorectal Cancer Patients with Vitamin D Insufficiency: Interim Analysis of a Randomized Controlled Trial. Nutrients 2022, 14, 4546. [Google Scholar] [CrossRef]
- Wimalawansa, S.J. Rapidly Increasing Serum 25(OH)D Boosts the Immune System, against Infections—Sepsis and COVID-19. Nutrients 2022, 14, 2997. [Google Scholar] [CrossRef]
- Schloss, J. Nutritional Deficiencies That May Predispose to Long COVID; Hungarian Academy of Science (MTA): Budapest, Hungary, 2022; Available online: https://mta.hu/esemenynaptar/2022-05-05-az-mta-195-kozgyulese-the-long-covid-syndrome-lingering-illness-afterthe-pandemic-4332 (accessed on 2 June 2022).
- Kiss, E.; Hajdu, A.; Forika, G.; Dank, M.; Krenacs, T.; Nemeth, Z. The Effect of Dietary Methyl-Donor Intake and Other Lifestyle Factors on Cancer Patients in Hungary. Cancers 2022, 14, 4432. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Number | Key Vitamin D Target Genes | Location | Function | Processes | Ref |
|---|---|---|---|---|---|
| 1 | ACVRL1 | plasma membrane | TGF-β receptor//serine/threonine kinases | signaling | [77] |
| 2 | CD14 | plasma membrane | TLR coreceptor | innate immunity | |
| 3 | CD93 | plasma membrane | intercellular adhesion, clearance of apoptotic cells | innate immunity | |
| 4 | LILRB4 | plasma membrane | leukocyte immunglobulin-like receptor (LIR) | inhibiting immune response | |
| 5 | LRRC25 | membrane bound in the cytoplasm | inhibition of IFN and NFκB signaling pathways | inhibiting immune response | |
| 6 | NINJ1 | plasma membrane | adhesion molecule | inflammation, cell death, axonal growth, cell chemotaxis, and angiogenesis | |
| 7 | SEMA6B | plasma membrane | axon guidance | NS development | |
| 8 | THBD | plasma membrane | thrombin receptor | coagulation | |
| 9 | TREM1 | plasma membrane | receptor (Ig superfamily member) | amplifies neutrophil and monocyte-mediated inflammatory responses | |
| 10 | CAMP | secreted | antimicrobial peptid | innate immunity | |
| 11 | FN1 | plasma membrane, extracellular matrix, secreted into the plasma (blood) | glycoprotein | cell adhesion and migration processes (embryogenesis, wound healing, blood coagulation, host defense, and metastasis) | |
| 12 | SRGN | secretory granules in hemapoetic cells | secretory granule formation, mediator of cytotoxic cell granule-mediated apoptosis, regulates MMP2 and TNF-α secretion, inhibits bone mineralization | innate and adaptive immunity, Ca2+ homesotasis | |
| 13 | CEBPB | nucleus | transcription factor, regulation of immune and inflammatory response, adipogenesis, gluconeogenesis, liver regeneration, hematopoesis, osteoblast differentiation, osteoclastogenesis | innate and adaptive immunity, metabolism, bone hoeostasis | |
| 14 | THEMIS2 | cytoplasm and nucleus | T cell receptor signaling, regulation of B cell activation, macrophage inflammatory response, promotes LPS-induced TLR4-mediated TNF production | innate and adaptive immunity | |
| 15 | MAPK13 | cytoplasm (member of MAP kinase family, a p38 MAP kinase) | mediates extracellular stimuli; activating ELK1, ATF2 trascription factors, prolactin signaling, | innate and adaptive immunity, proliferation, differentiation, transcription regulation, development | |
| 1 | CYP24A1 | mitochondria | degradation of 1,25-dihydroxyvitamin D3, calcium homeostasis, | vitamin D endocrine system | [83] |
| 2 | G0S2 | mitochondria | positive regulation of extrinsic apoptotic pathway, binding to BCL2 | regulation of apoptosis | |
| 3 | HBEGF | secreted, extracellular space | growth factor | SMC proliferation, cardiac valve formation, normal heart function | |
| 4 | SEMA6B | type I membrane protein | cell surface repellent | axon guidance | |
| 5 | THBD | endothelial specific type I membrane receptor | receptor of thrombin | bind thrombin, conversion of protein C to activated protein C | |
| 6 | AQP9 | plasma membrane, multi pass membrane protein | water channel | permeable for glyerol and urea, mediate passage of small, non-charged solutes (carbamides, polyols, purines, pyrimidines) | |
| 7 | CCL7 | secreted | chemotactic factor, attracts moncytes and eosinophils but not neutrophils, binds heparin, CCR1/2/3 | attracts macrophages during inflammation and metastasis, augments monocytes antitumor activity, induces release of gelatinase B | |
| 8 | PVALB | cytoplasm, nucleoplasm | high-affinity calcium binding | muscle relaxation in muscle cells, calcium binding in GABAergic cells | |
| 9 | CD1E | plasma membrane, MHC-type protein | form heterodimers with beta-2-microglobulin | presentation of lipid and glycolipid antigens to T cells | |
| 10 | NRG1 | plasma membrane, single-pass type I membrane protein | ligand of ERBB3/4 tyrosine kinase receptors | growth, differentiation of epithelial, glial, neuronal and skeletal muscle cells, stimulating milk production, mammary tumor cell differentiation | |
| 1 | CD14 | plasma membrane | TLR co-receptor | innate immunity (independent of the size of the tested target gene set, gene ontology analysis indicates that the modulation of innate immunity is the main physiological outcome of a vitamin D stimulation of human monocytes) | [84] |
| 2 | ORM1 | vesicles in the cytoplasm | acute phase plasma protein | ||
| 3 | CAMP | secreted | antimicrobial peptid | ||
| 4 | FBP1 | mitochondria | glucose metabolizing enzyme | ||
| 5 | CYP26B1 | endoplasmatic reticulum | Drug- and retinoid metabolism, and synthesis of cholesterol, steroids and other lipids | ||
| 6 | TSPAN18 | plasma membrane | unknown | ||
| 7–19 | +13 other genes | top five biological pathways: neutrophil activation, positive regulation of TNF production, inflammatory response, neutrophil degranulation, negative regulation of T cell proliferation, positive regulation of cytokine secretion | |||
| 25(OH)D3 Level (ng/mL) | Recommendation of IOM | Recommendation of ES |
|---|---|---|
| lower than 12 | deficiency | deficiency |
| 12–19 | inadequacy | deficiency |
| 20–29 | sufficiency | insufficiency |
| 30–49 | sufficiency | sufficiency |
| higher than 50 | reason for concern | sufficiency |
| Vitamin D | SIRT1 | Tissue/Organ Type | Diseases | Signal Pathway/Targeted Molecules | Prevention/Complementary Treatment Options |
|---|---|---|---|---|---|
| decrease: BMI, body fat, TGs, CRP, TNFα. increase: SIRT1/6, HDL-C, apo-AI, ABCA1, FFA oxidation | decrease: adipocyte differentiation-PPARγ, inflammation, leptin increase: activates PPAR-α, lipolysis, FFA oxidation, PGC-1α, insulin/mTOR/cell growth, leptin/PI3K-Akt-mTORC1 | adipose (white/brown) | T2DM, obesity, insulin resistance | adiponectin/NFĸB, AMPK, PPAR-α | • weight loss • exercise/physical activity • emotional/mental support • nutrition factors (vitamin D, polyphenols, carotinoids, omega-3 FA) |
| increase: glucose uptake and utilization | increase: FFA oxidation, PGC-1α | skeltal muscle | insulin resistance | adiponectin/NFĸB, AMPK, PPAR-α | |
| increase: NO production, fat oxidation decrease: blood pressure, cardiac hypertophy, foam cell formation, macrophage activation, vascular inflammation, oxidative stress | decrease: cardiac hypertophy, proinflammatory macrophage activity increase: PPAR-α, eNOS/NO, FOXOs | endothel | cardiovascular abnormalities | adiponectin/NFĸB, AMPK, PPAR-α | |
| decrease: renin-angiotensin-aldosterone axis | kidney | CKD, hypertension | |||
| decrease: inflammation, proinflammatory cytokines, TNFα and CRP, mTOR, ROS/RNS increase: IL-10, IL-1RA, BCAA catabolism, antioxidant enzymes | decrease: TLR4, IFNγ, IL-1β, IL-6, IL-8 and TNFα levels, oxidative stress, ROS, iNOS, COX-2 | immune system | TLR4 Akt/NFĸB/COX-2, Nrf2 | ||
| decrease: β-catenin, WNT/β-catenin pathway, c-Myc, TCF1, LEF1, antiapoptotic proteins, HIF-1, VEGF, IL-8 increase: p21WAF/CIP and p27KIP, E-cadherin, DKK-1, proapoptotic proteins, Nrf2 | decrease: NFκB, MMP-9, insulin/mTOR/cell growth, leptin/PI3K-Akt-mTORC1, TNF-α, IL-1β, iNOS, COX-2; initiation, promotion, and progression of cancer development increase: ROS, apoptosis, cell cycle arrest | PC, BC, CRC, SkinC | cancer | insulin/mTOR/cell growth, proliferation of leptin/PI3K-Akt-mTORC1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nemeth, Z.; Patonai, A.; Simon-Szabó, L.; Takács, I. Interplay of Vitamin D and SIRT1 in Tissue-Specific Metabolism—Potential Roles in Prevention and Treatment of Non-Communicable Diseases Including Cancer. Int. J. Mol. Sci. 2023, 24, 6154. https://doi.org/10.3390/ijms24076154
Nemeth Z, Patonai A, Simon-Szabó L, Takács I. Interplay of Vitamin D and SIRT1 in Tissue-Specific Metabolism—Potential Roles in Prevention and Treatment of Non-Communicable Diseases Including Cancer. International Journal of Molecular Sciences. 2023; 24(7):6154. https://doi.org/10.3390/ijms24076154
Chicago/Turabian StyleNemeth, Zsuzsanna, Attila Patonai, Laura Simon-Szabó, and István Takács. 2023. "Interplay of Vitamin D and SIRT1 in Tissue-Specific Metabolism—Potential Roles in Prevention and Treatment of Non-Communicable Diseases Including Cancer" International Journal of Molecular Sciences 24, no. 7: 6154. https://doi.org/10.3390/ijms24076154
APA StyleNemeth, Z., Patonai, A., Simon-Szabó, L., & Takács, I. (2023). Interplay of Vitamin D and SIRT1 in Tissue-Specific Metabolism—Potential Roles in Prevention and Treatment of Non-Communicable Diseases Including Cancer. International Journal of Molecular Sciences, 24(7), 6154. https://doi.org/10.3390/ijms24076154