
Comparing Vaginal and Endometrial Microbiota Using Culturomics: Proof of Concept

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
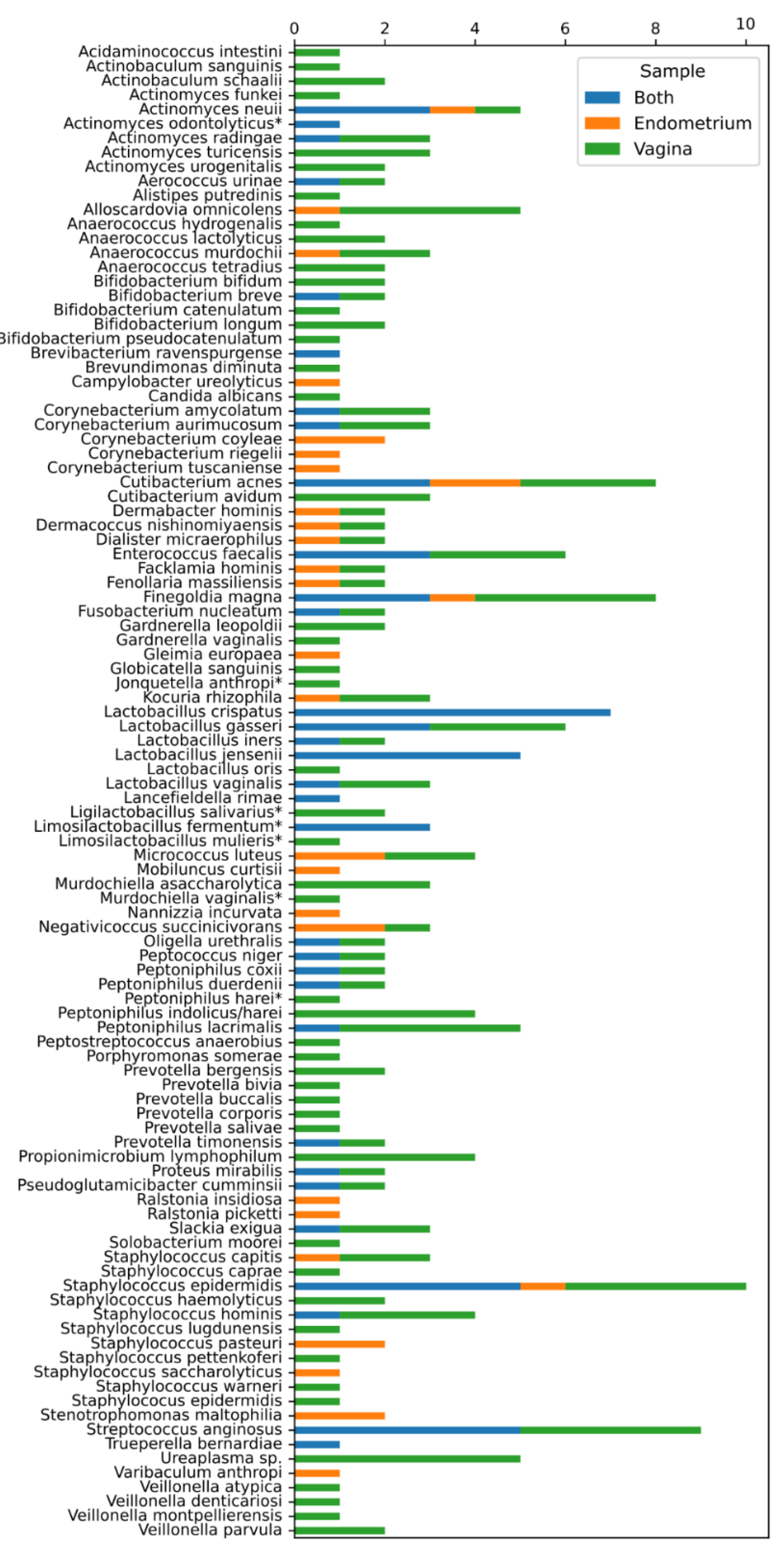
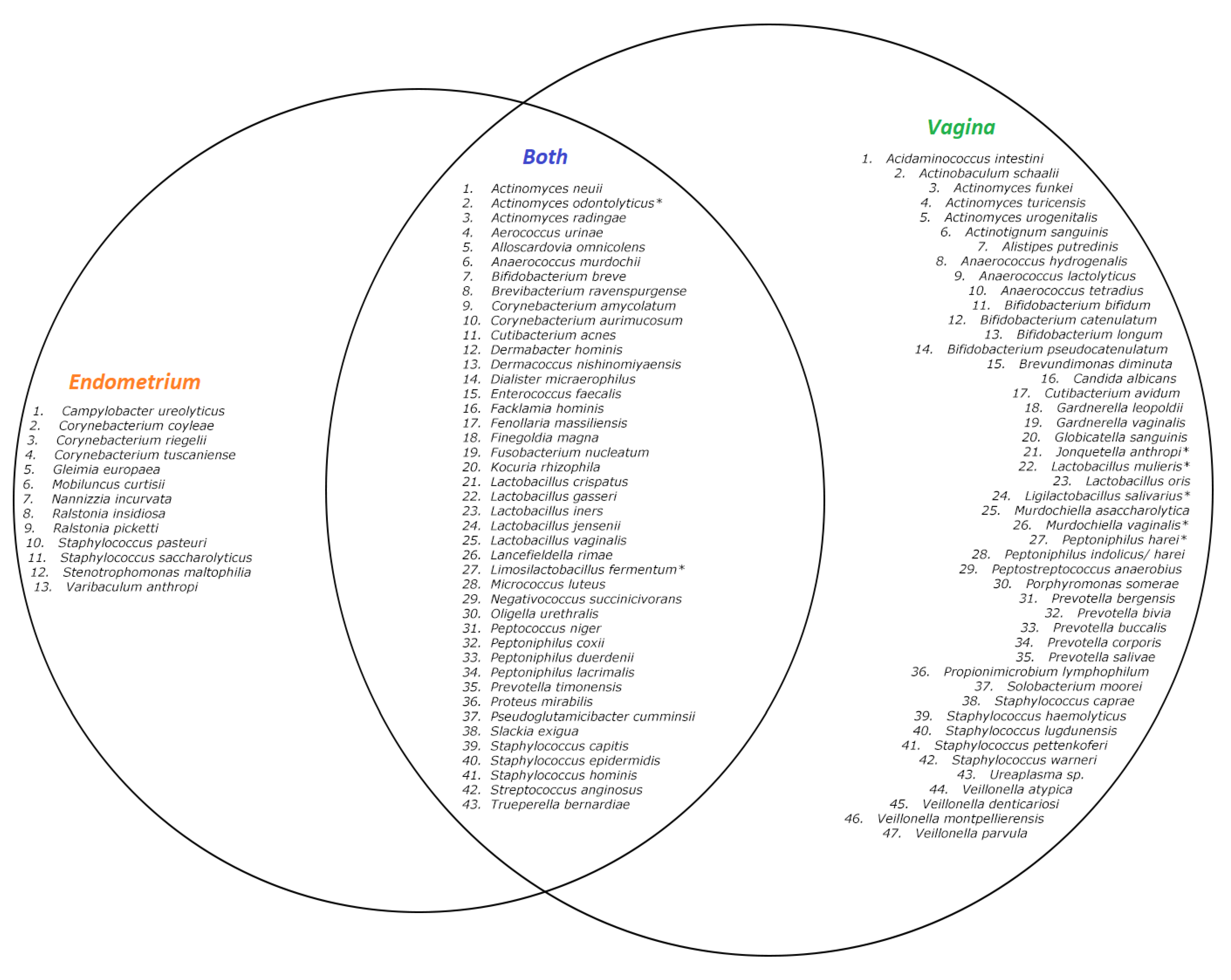
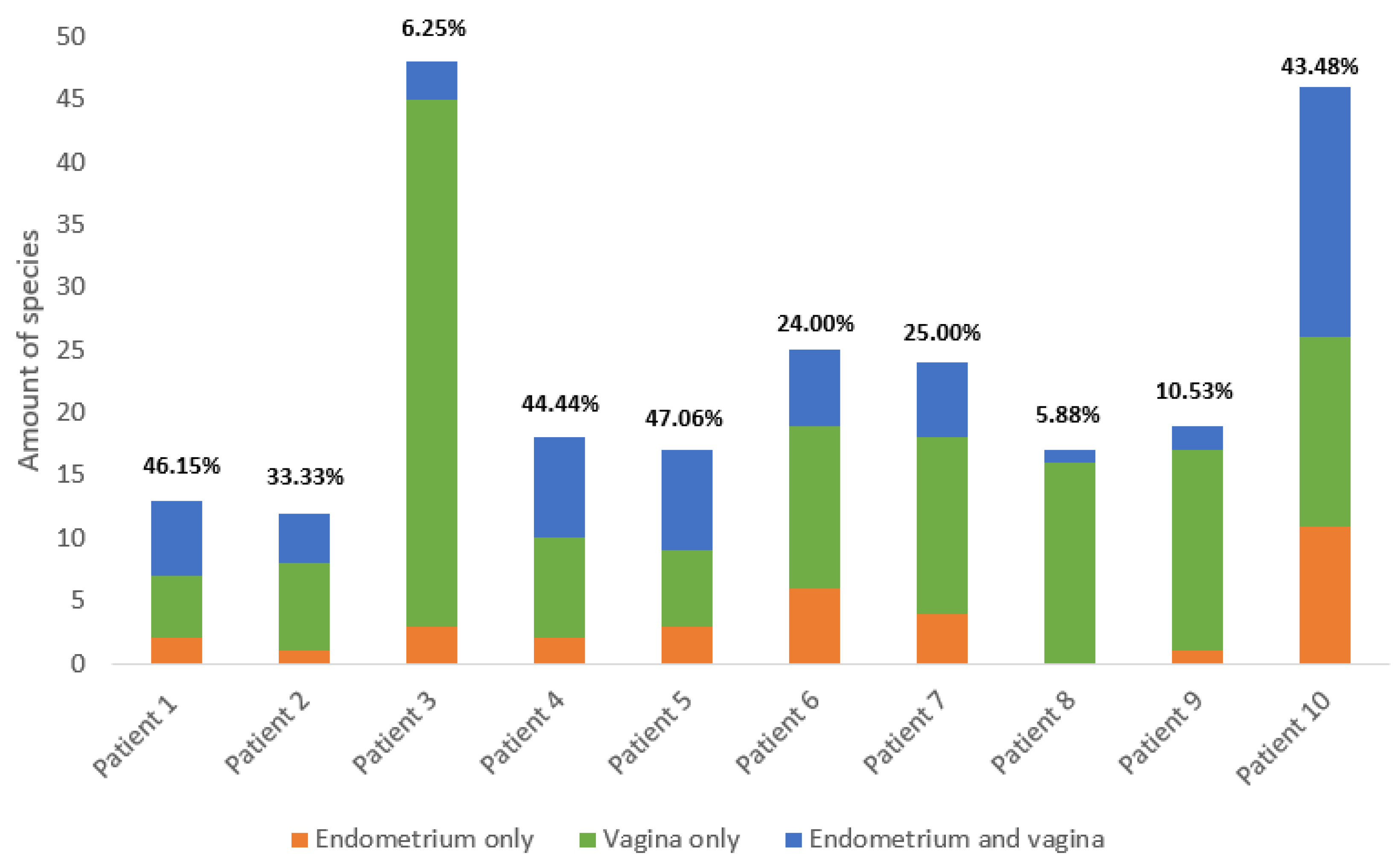
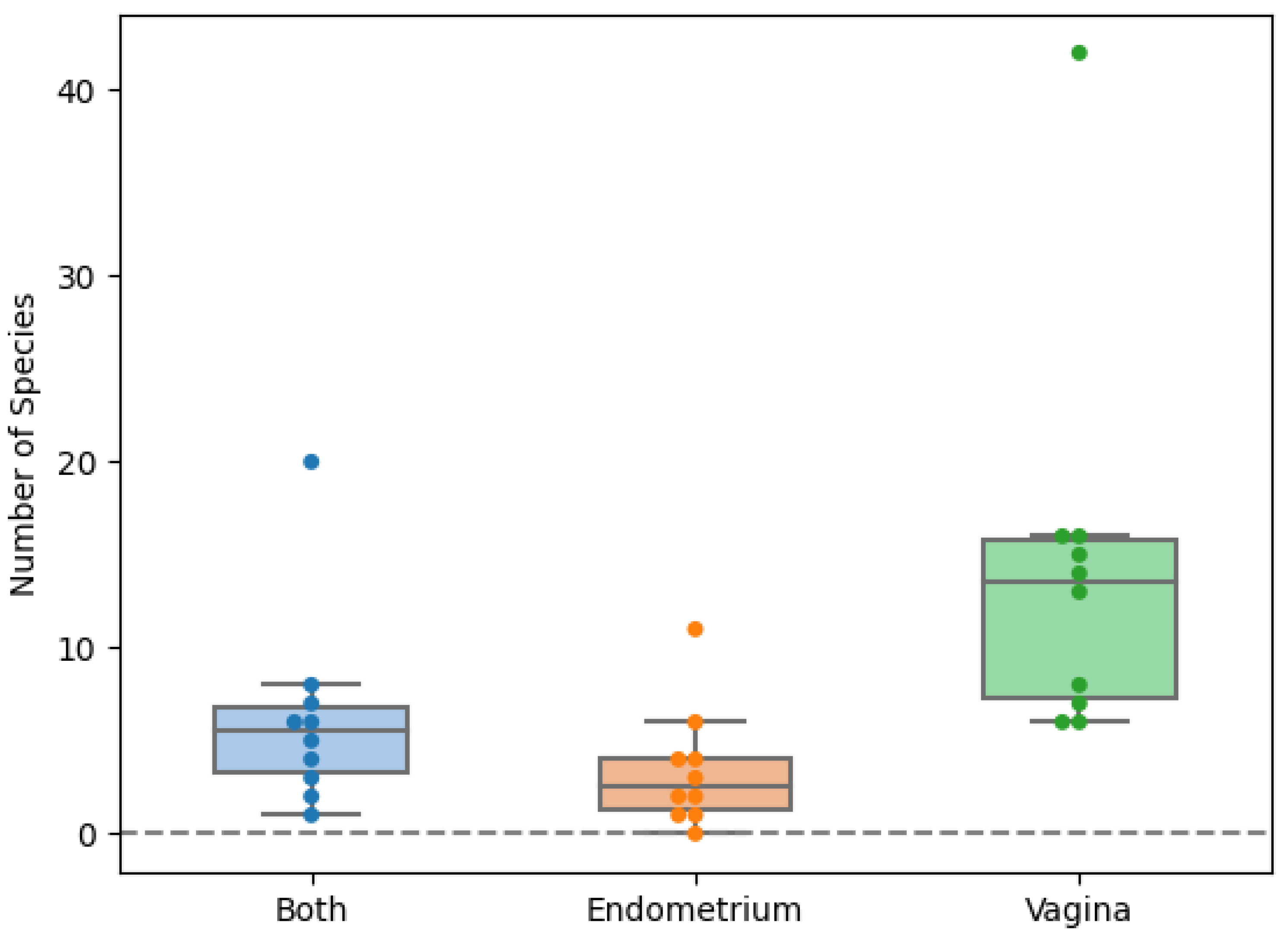
2. Results
2.1. Comparing the Endometrial and Vaginal Microbiota
2.2. Lactobacillus Species
3. Discussion
4. Materials and Methods
4.1. Setting and Study Design
4.2. Culturomics
4.2.1. Direct Inoculation
4.2.2. Pre-Incubation
4.2.3. Identification of the Colonies
4.3. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, C.; Song, X.; Wei, W.; Zhong, H.; Dai, J.; Lan, Z.; Li, F.; Yu, X.; Feng, Q.; Wang, Z.; et al. The microbiota continuum along the female reproductive tract and its relation to uterine-related diseases. Nat. Commun. 2017, 8, 875. [Google Scholar] [CrossRef]
- Punzón-Jiménez, P.; Labarta, E. The impact of the female genital tract microbiome in women health and reproduction: A review. J. Assist. Reprod. Genet. 2021, 38, 2519–2541. [Google Scholar] [CrossRef]
- Aroutcheva, A.; Gariti, D.; Simon, M.; Shott, S.; Faro, J.; Simoes, J.A.; Gurguis, A.; Faro, S. Defense factors of vaginal lactobacilli. Am. J. Obstet. Gynecol. 2001, 185, 375–379. [Google Scholar] [CrossRef]
- Boskey, E.R.; Cone, R.A.; Whaley, K.J.; Moench, T.R. Origins of vaginal acidity: High D/L lactate ratio is consistent with bacteria being the primary source. Hum. Reprod. 2001, 16, 1809–1813. [Google Scholar] [CrossRef]
- Gupta, S.; Kakkar, V.; Bhushan, I. Crosstalk between Vaginal Microbiome and Female Health: A review. Microb. Pathog. 2019, 136, 103696. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Ahannach, S.; Wittouck, S.; Gehrmann, T.; Eilers, T.; Oerlemans, E.; Condori, S.; Dillen, J.; Spacova, I.; Vander Donck, L.; et al. Citizen-science map of the vaginal microbiome. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Oerlemans, E.F.M.; Wuyts, S.; Bellen, G.; Wittouck, S.; De Boeck, I.; Ruban, K.; Allonsius, C.N.; van den Broek, M.F.L.; Donders, G.G.G.; Lebeer, S. The Dwindling Microbiota of Aerobic Vaginitis, an Inflammatory State Enriched in Pathobionts with Limited TLR Stimulation. Diagnostics 2020, 10, 879. [Google Scholar] [CrossRef] [PubMed]
- Campisciano, G.; Florian, F.; D’Eustacchio, A.; Stanković, D.; Ricci, G.; De Seta, F.; Comar, M. Subclinical alteration of the cervical-vaginal microbiome in women with idiopathic infertility. J. Cell. Physiol. 2017, 232, 1681–1688. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.; Machado, D.; Cerca, N. Unveiling the role of Gardnerella vaginalis in polymicrobial Bacterial Vaginosis biofilms: The impact of other vaginal pathogens living as neighbors. ISME J. 2019, 13, 1306–1317. [Google Scholar] [CrossRef]
- Harwich, M.D.; Alves, J.M.; A Buck, G.; Strauss, J.F.; Patterson, J.L.; Oki, A.T.; Girerd, P.H.; Jefferson, K.K. Drawing the line between commensal and pathogenic Gardnerella vaginalis through genome analysis and virulence studies. BMC Genom. 2010, 11, 375. [Google Scholar] [CrossRef]
- Toson, B.; Simon, C.; Moreno, I. The Endometrial Microbiome and Its Impact on Human Conception. Int. J. Mol. Sci. 2022, 23, 485. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.M.; Chase, D.M.; Herbst-Kralovetz, M.M. Uterine Microbiota: Residents, Tourists, or Invaders? Front. Immunol. 2018, 2, 208. [Google Scholar] [CrossRef] [PubMed]
- Moreno, I.; Franasiak, J.M. Endometrial microbiota-new player in town. Fertil. Steril. 2017, 108, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Moreno, I.; Codoñer, F.M.; Vilella, F.; Valbuena, D.; Martinez-Blanch, J.F.; Jimenez-Almazán, J.; Alonso, R.; Alamá, P.; Remohí, J.; Pellicer, A.; et al. Evidence that the endometrial microbiota has an effect on implantation success or failure. Am. J. Obstet. Gynecol. 2016, 215, 684–703. [Google Scholar] [CrossRef]
- Hashimoto, T.; Kyono, K. Does dysbiotic endometrium affect blastocyst implantation in IVF patients? J. Assist. Reprod. Genet. 2019, 36, 2471–2479. [Google Scholar] [CrossRef]
- Salter, S.J.; Cox, M.J.; Turek, E.M.; Calus, S.T.; Cookson, W.O.; Moffatt, M.F.; Turner, P.; Parkhill, J.; Loman, N.J.; Walker, A.W. Reagent and laboratory contamination can critically impact sequence-based microbiome analyses. BMC Biol. 2014, 12, 87. [Google Scholar] [CrossRef]
- Brooks, J.P.; Edwards, D.J.; Harwich, M.D., Jr.; Rivera, M.C.; Fettweis, J.M.; Serrano, M.G.; Reris, R.A.; Sheth, N.U.; Huang, B.; Girerd, P.; et al. The truth about metagenomics: Quantifying and counteracting bias in 16S rRNA studies. BMC Microbiol. 2015, 15, 66. [Google Scholar] [CrossRef]
- Kennedy, K.M.; de Goffau, M.C.; Perez-Muñoz, M.E.; Arrieta, M.-C.; Bäckhed, F.; Bork, P.; Braun, T.; Bushman, F.D.; Dore, J.; de Vos, W.M.; et al. Questioning the fetal microbiome illustrates pitfalls of low-biomass microbial studies. Nature 2023, 613, 639–649. [Google Scholar] [CrossRef]
- Lagier, J.C.; Armougom, F.; Million, M.; Hugon, P.; Pagnier, I.; Robert, C.; Bittar, F.; Fournous, G.; Gimenez, G.; Maraninchi, M.; et al. Endometrial microbiome: Sampling, assessment, and possible impact on embryo implantation. Sci. Rep. 2022, 12, 8467. [Google Scholar]
- Lagier, J.C.; Armougom, F.; Million, M.; Hugon, P.; Pagnier, I.; Robert, C.; Bittar, F.; Fournous, G.; Gimenez, G.; Maraninchi, M.; et al. Microbial culturomics: Paradigm shift in the human gut microbiome study. Clin. Microbiol. Infect. 2012, 18, 1185–1193. [Google Scholar] [CrossRef]
- Vanstokstraeten, R.; Mackens, S.; Callewaert, E.; Blotwijk, S.; Emmerechts, K.; Crombé, F.; Soetens, O.; Wybo, I.; Vandoorslaer, K.; Mostert, L.; et al. Culturomics to Investigate the Endometrial Microbiome: Proof-of-Concept. Int. J. Mol. Sci. 2022, 23, 12212. [Google Scholar] [CrossRef] [PubMed]
- Lagier, J.C.; Dubourg, G.; Million, M.; Cadoret, F.; Bilen, M.; Fenollar, F.; Levasseur, A.; Rolain, J.M.; Fournier, P.E.; Raoult, D. Culturing the human microbiota and culturomics. Nat. Rev. Microbiol. 2018, 16, 540–550. [Google Scholar] [CrossRef]
- Smolnikova, V.; Keburiya, L.; Iteimowei, M.; Priputnevich, T.; Muravieva, V.; Kalinina, E.; Sukhikh, G. Influence of Endometrial Microbiota on Reproductive Outcomes in IVF Programs. Am. J. Biomed. Sci. Res. 2019, 4, 197–200. [Google Scholar] [CrossRef]
- Srinivasan, S.; Munch, M.M.; Sizova, M.V.; Fiedler, T.L.; Kohler, C.M.; Hoffman, N.G.; Liu, C.; Agnew, K.J.; Marrazzo, J.M.; Epstein, S.S.; et al. More Easily Cultivated Than Identified: Classical Isolation with Molecular Identification of Vaginal Bacteria. J. Infect. Dis. 2016, 214 (Suppl. S1), S21–S28. [Google Scholar] [CrossRef]
- Liang, G.; Bushman, F.D. The human virome: Assembly, composition and host interactions. Nat. Rev. Microbiol. 2021, 19, 514–527. [Google Scholar] [CrossRef]
- Lykke, M.R.; Becher, N.; Haahr, T.; Boedtkjer, E.; Jensen, J.S.; Uldbjerg, N. Vaginal, Cervical and Uterine pH in Women with Normal and Abnormal Vaginal Microbiota. Pathogens 2021, 10, 90. [Google Scholar] [CrossRef]
- Svenstrup, H.F.; Fedder, J.; Abraham-Peskir, J.; Birkelund, S.; Christiansen, G. Mycoplasma genitalium attaches to human spermatozoa. Hum. Reprod. 2003, 18, 2103–2109. [Google Scholar] [CrossRef] [PubMed]
- Sparks, R.A.; Purrier, B.G.; Watt, P.J.; Elstein, M. Bacteriological colonisation of uterine cavity: Role of tailed intrauterine contraceptive device. Br. Med. J. 1981, 282, 1189–1191. [Google Scholar] [CrossRef]
- Castillo, D.J.; Rifkin, R.F.; Cowan, D.A.; Potgieter, M. The Healthy Human Blood Microbiome: Fact or Fiction? Front. Cell. Infect. Microbiol. 2019, 9, 148. [Google Scholar] [CrossRef]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4680–4687. [Google Scholar] [CrossRef]
- Demuyser, T.; De Geyter, D.; Van Dorpe, D.; Vandoorslaer, K.; Wybo, I. Extensive evaluation of fastidious anaerobic bacteria recovery from the Copan eSwab® transport system. J. Microbiol. Methods 2018, 144, 73–78. [Google Scholar] [CrossRef] [PubMed]
- MM18. Interprative Criteria for Identification of bacteria and Fungi by targeted DNA Sequencing. Inst. Clin. Lab. Stand. 2018, 25–117.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Age (Years) | Ethnicity | Vagina | Cervix | Cavum | Left Ostium | Right Ostium | Anatomopathology |
|---|---|---|---|---|---|---|---|---|
| 1 | 44 | Caucasian | Normal | Normal | Normal | Normal | Normal | Normal histology |
| 2 | 30 | Caucasian | Normal | Normal | Normal | Normal | Normal | Normal histology |
| 3 | 34 | Caucasian | Normal | Normal | Normal | Normal | Normal | Normal histology |
| 4 | 44 | Caucasian | Normal | Normal | Normal | Normal | Normal | Isolated plasmacells |
| 5 | 31 | Caucasian | Normal | Normal | Inflammatory | Normal | Normal | Isolated plasmacells |
| 6 | 35 | Caucasian | Normal | Normal | Normal | Normal | Normal | Normal histology |
| 7 | 41 | Caucasian | Normal | Normal | Atrophic | Normal | Normal | Normal histology |
| 8 | 34 | Caucasian | Normal | Normal | Normal | Normal | Normal | Isolated plasmacells |
| 9 | 36 | Caucasian | Normal | Normal | Normal | Normal | Normal | Isolated plasmacells |
| 10 | 43 | Caucasian | Normal | Normal | Normal | Normal | Normal | Isolated plasmacells |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vanstokstraeten, R.; Callewaert, E.; Blotwijk, S.; Rombauts, E.; Crombé, F.; Emmerechts, K.; Soetens, O.; Vandoorslaer, K.; De Geyter, D.; Allonsius, C.; et al. Comparing Vaginal and Endometrial Microbiota Using Culturomics: Proof of Concept. Int. J. Mol. Sci. 2023, 24, 5947. https://doi.org/10.3390/ijms24065947
Vanstokstraeten R, Callewaert E, Blotwijk S, Rombauts E, Crombé F, Emmerechts K, Soetens O, Vandoorslaer K, De Geyter D, Allonsius C, et al. Comparing Vaginal and Endometrial Microbiota Using Culturomics: Proof of Concept. International Journal of Molecular Sciences. 2023; 24(6):5947. https://doi.org/10.3390/ijms24065947
Chicago/Turabian StyleVanstokstraeten, Robin, Ellen Callewaert, Susanne Blotwijk, Eleni Rombauts, Florence Crombé, Kristof Emmerechts, Oriane Soetens, Kristof Vandoorslaer, Deborah De Geyter, Camille Allonsius, and et al. 2023. "Comparing Vaginal and Endometrial Microbiota Using Culturomics: Proof of Concept" International Journal of Molecular Sciences 24, no. 6: 5947. https://doi.org/10.3390/ijms24065947
APA StyleVanstokstraeten, R., Callewaert, E., Blotwijk, S., Rombauts, E., Crombé, F., Emmerechts, K., Soetens, O., Vandoorslaer, K., De Geyter, D., Allonsius, C., Vander Donck, L., Blockeel, C., Wybo, I., Piérard, D., Demuyser, T., & Mackens, S. (2023). Comparing Vaginal and Endometrial Microbiota Using Culturomics: Proof of Concept. International Journal of Molecular Sciences, 24(6), 5947. https://doi.org/10.3390/ijms24065947