

New Data on Anti-Inflammatory and Wound Healing Potential of Transgenic Senna obtusifolia Hairy Roots: In Vitro Studies

 ,
,  , ,
, ,  ,
,  and
and 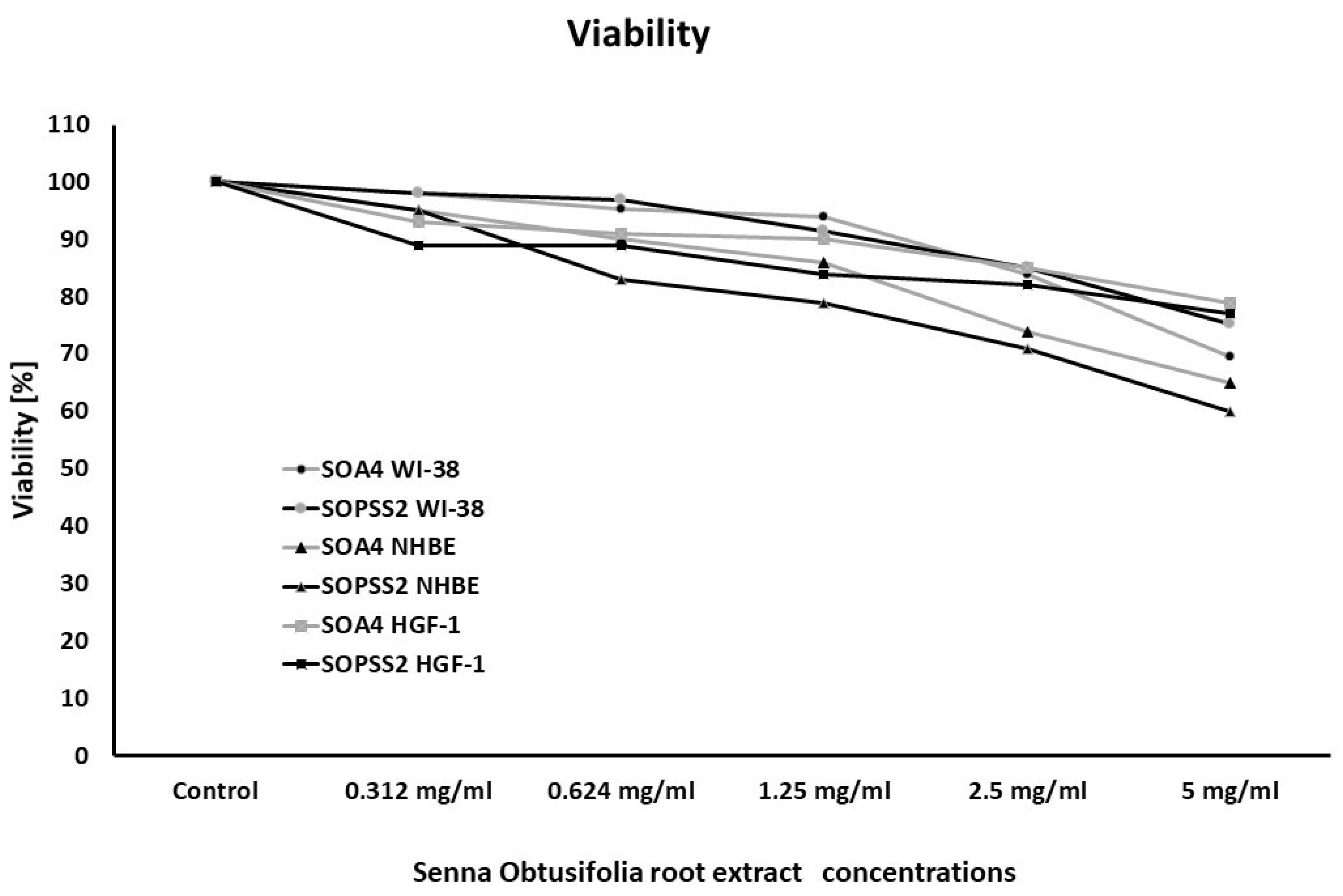{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Phytochemical Analysis
2.2. Cell Viability after Treatment with SOA4 and SOPSS2 Root Extracts from Senna Obtusifolia
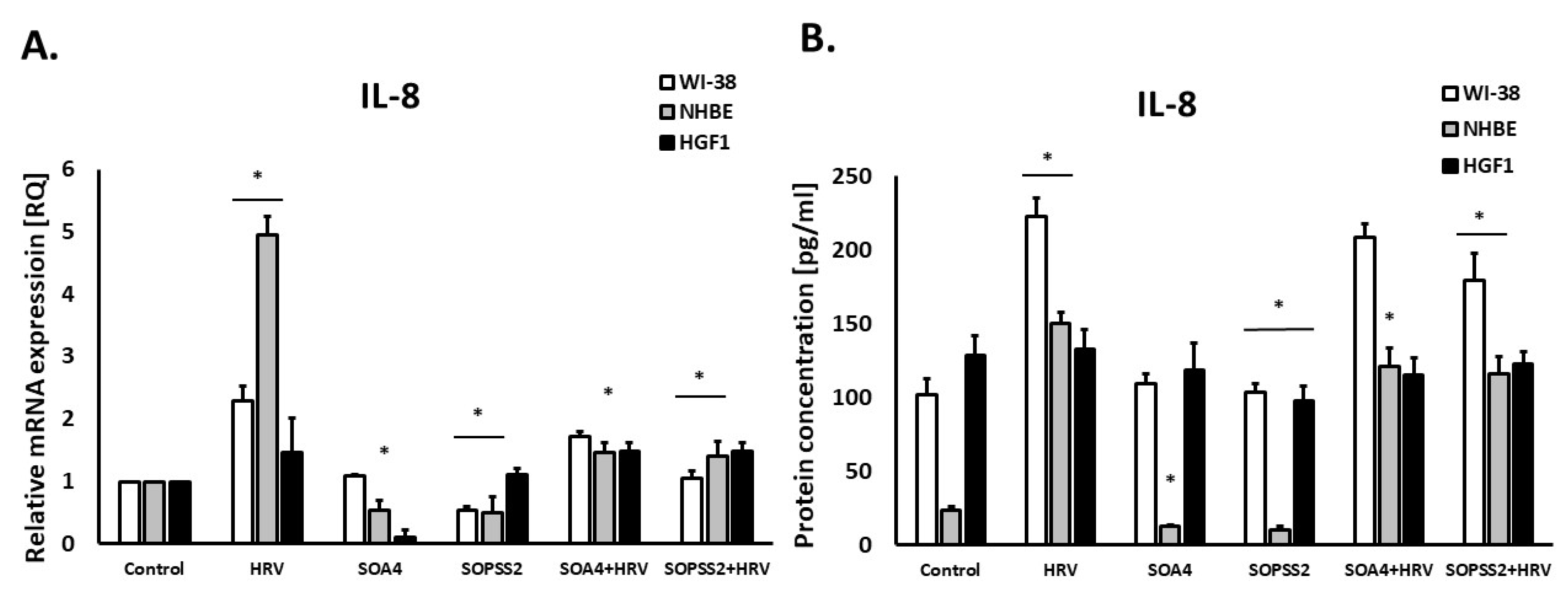
2.3. Immunomodulatory Effects
2.4. Effect of SOPSS2 Hairy Root Extract of Senna obtusifolia on the Migration of Fibroblast Cells
2.5. The Effect of SOA4 and SOPSS2 of Senna obtusifolia Root Extracts on Thiol Groups
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. SOA4 and SOPSS2 of Senna obtusifolia Root Extracts Preparation
4.3. Cell Cultures
4.4. Virus Preparation and Cell Infection
4.5. Experimental Procedure
4.6. RNA Isolation and cDNA Synthesis
4.7. Gene Expression Analysis
4.8. Cytokine Detection in Cells
4.9. Colorimetric Thiol Detection
4.10. In Vitro Wound Healing Potency
4.11. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef]
- Beukes, N.; Levendal, R.A.; Frost, C.L. Selected terpenoids from medicinal plants modulate endoplasmic reticulum stress in metabolic disorders. J. Pharm. Pharmacol. 2014, 66, 1505–1525. [Google Scholar] [CrossRef]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of secondary metabolites in plant defense against pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef] [PubMed]
- WHO Monographs on Selected Medicinal Plants. 1. Plants, Medicinal. 2. Angiosperms. 3. Medicine, Traditional. In WHO Consultation on Selected Medicinal Plants (4th: 2005: Salerno-Paestum, Italy); World Health Organization: Geneva, Switzerland, 2005; Volume 4, ISBN 9789241547055. [Google Scholar]
- Wieczfinska, J.; Sitarek, P.; Kowalczyk, T.; Rieske, P.; Pawliczak, R. Curcumin modulates airway remodelling-contributing genes-the significance of transcription factors. J. Cell. Mol. Med. 2022, 26, 736–749. [Google Scholar] [CrossRef] [PubMed]
- Wieczfinska, J.; Sitarek, P.; Kowalczyk, T.; Pawliczak, R. Leonurus sibiricus root extracts decrease airway remodeling markers expression in fibroblasts. Clin. Exp. Immunol. 2020, 202, 28–46. [Google Scholar] [CrossRef] [PubMed]
- Sitarek, P.; Kowalczyk, T.; Wieczfinska, J.; Merecz-Sadowska, A.; Gorski, K.; Sliwinski, T.; Skala, E. Plant Extracts as a Natural Source of Bioactive Compounds and Potential Remedy for the Treatment of Certain Skin Diseases. Curr. Pharm. Des. 2020, 26, 2859–2875. [Google Scholar] [CrossRef]
- Skala, E.; Makowczynska, J.; Wieczfinska, J.; Kowalczyk, T.; Sitarek, P. Caffeoylquinic Acids with Potential Biological Activity from Plant In vitro Cultures as Alternative Sources of Valuable Natural Products. Curr. Pharm. Des. 2020, 26, 2817–2842. [Google Scholar] [CrossRef]
- Sheludko, Y.V. Recent advances in plant biotechnology and genetic engineering for production of secondary metabolites. Tsitol Genet. 2010, 44, 65–75. [Google Scholar] [CrossRef]
- Georgiev, M.I. Contemporary approaches towards production of phytopharmaceuticals: Plant biotechnology meets medicinal chemistry. Mini Rev. Med. Chem. 2011, 11, 822. [Google Scholar] [CrossRef]
- Mao, R.; Xia, P.; He, Z.; Liu, Y.; Liu, F.; Zhao, H.; Han, R.; Liang, Z. Identification of seeds based on molecular markers and secondary metabolites in Senna obtusifolia and Senna occidentalis. Bot. Stud. 2017, 58, 43. [Google Scholar] [CrossRef]
- Kowalczyk, T.; Sitarek, P.; Toma, M.; Picot, L.; Wielanek, M.; Skala, E.; Sliwinski, T. An Extract of Transgenic Senna obtusifolia L. Hairy Roots with Overexpression of PgSS1 Gene in Combination with Chemotherapeutic Agent Induces Apoptosis in the Leukemia Cell Line. Biomolecules 2020, 10, 510. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.J.; Brightling, C. Pathogenesis of asthma: Implications for precision medicine. Clin. Sci. 2017, 131, 1723–1735. [Google Scholar] [CrossRef]
- Hammad, H.; Lambrecht, B.N. The basic immunology of asthma. Cell 2021, 184, 1469–1485. [Google Scholar] [CrossRef] [PubMed]
- Kirby, T. Christine Jenkins-global advocate for asthma and COPD. Lancet Respir. Med. 2022, 10, 431. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xiao, Z.; Jiang, Z.; Jiang, Y.; Li, W.; Wang, M. Schisandrin B Attenuates Airway Inflammation and Airway Remodeling in Asthma by Inhibiting NLRP3 Inflammasome Activation and Reducing Pyroptosis. Inflammation 2021, 44, 2217–2231. [Google Scholar] [CrossRef]
- Wieczfinska, J.; Sitarek, P.; Skala, E.; Kowalczyk, T.; Pawliczak, R. Inhibition of NADPH Oxidase-Derived Reactive Oxygen Species Decreases Expression of Inflammatory Cytokines in A549 Cells. Inflammation 2019, 42, 2205–2214. [Google Scholar] [CrossRef]
- Luo, J.; Zhang, L.; Zhang, X.; Long, Y.; Zou, F.; Yan, C.; Zou, W. Protective effects and active ingredients of Salvia miltiorrhiza Bunge extracts on airway responsiveness, inflammation and remodeling in mice with ovalbumin-induced allergic asthma. Phytomedicine 2019, 52, 168–177. [Google Scholar] [CrossRef]
- Guan, S.; Yu, P.; Cao, J.; Xi, X.; Zhang, Q.; Zhu, C.; Hu, H.; Gong, X.; Fan, H. Ginsenoside Rg1 protects against cigarette smoke-induced airway remodeling by suppressing the TGF-beta1/Smad3 signaling pathway. Am. J. Transl. Res. 2020, 12, 493–506. [Google Scholar]
- Kowalczyk, T.; Sitarek, P.; Toma, M.; Rijo, P.; Dominguez-Martin, E.; Falco, I.; Sanchez, G.; Sliwinski, T. Enhanced Accumulation of Betulinic Acid in Transgenic Hairy Roots of Senna obtusifolia Growing in the Sprinkle Bioreactor and Evaluation of Their Biological Properties in Various Biological Models. Chem. Biodivers. 2021, 18, e2100455. [Google Scholar] [CrossRef]
- Wirz, O.F.; Jansen, K.; Satitsuksanoa, P.; van de Veen, W.; Tan, G.; Sokolowska, M.; Mirer, D.; Stanic, B.; Message, S.D.; Kebadze, T.; et al. Experimental rhinovirus infection induces an antiviral response in circulating B cells which is dysregulated in patients with asthma. Allergy 2022, 77, 130–142. [Google Scholar] [CrossRef]
- Yang, Z.; Mitlander, H.; Vuorinen, T.; Finotto, S. Mechanism of Rhinovirus Immunity and Asthma. Front. Immunol. 2021, 12, 731846. [Google Scholar] [CrossRef] [PubMed]
- Biagi, C.; Rocca, A.; Poletti, G.; Fabi, M.; Lanari, M. Rhinovirus Infection in Children with Acute Bronchiolitis and Its Impact on Recurrent Wheezing and Asthma Development. Microorganisms 2020, 8, 1620. [Google Scholar] [CrossRef]
- Lambert, K.A.; Prendergast, L.A.; Dharmage, S.C.; Tang, M.; O’Sullivan, M.; Tran, T.; Druce, J.; Bardin, P.; Abramson, M.J.; Erbas, B. The role of human rhinovirus (HRV) species on asthma exacerbation severity in children and adolescents. J. Asthma 2018, 55, 596–602. [Google Scholar] [CrossRef]
- Lukkarinen, M.; Jartti, T. The first rhinovirus-wheeze acts as a marker for later asthma in high-risk children. J. Allergy Clin. Immunol. 2016, 138, 313. [Google Scholar] [CrossRef] [PubMed]
- Teeratakulpisarn, J.; Pientong, C.; Ekalaksananan, T.; Ruangsiripiyakul, H.; Uppala, R. Rhinovirus infection in children hospitalized with acute bronchiolitis and its impact on subsequent wheezing or asthma: A comparison of etiologies. Asian Pac. J. Allergy Immunol. 2014, 32, 226–234. [Google Scholar] [CrossRef]
- Plesa, M.; Gaudet, M.; Mogas, A.; Olivenstein, R.; Al Heialy, S.; Hamid, Q. Action of 1,25(OH)2D3 on Human Asthmatic Bronchial Fibroblasts: Implications for Airway Remodeling in Asthma. J. Asthma Allergy 2020, 13, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Berry, M.; Brightling, C.; Pavord, I.; Wardlaw, A. TNF-alpha in asthma. Curr. Opin. Pharmacol. 2007, 7, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Michalik, M.; Wojcik-Pszczola, K.; Paw, M.; Wnuk, D.; Koczurkiewicz, P.; Sanak, M.; Pekala, E.; Madeja, Z. Fibroblast-to-myofibroblast transition in bronchial asthma. Cell. Mol. Life Sci. 2018, 75, 3943–3961. [Google Scholar] [CrossRef]
- Ali, M.Y.; Park, S.; Chang, M. Phytochemistry, Ethnopharmacological Uses, Biological Activities, and Therapeutic Applications of Cassia obtusifolia L.: A Comprehensive Review. Molecules 2021, 26, 6252. [Google Scholar] [CrossRef]
- Howarth, P.H.; Babu, K.S.; Arshad, H.S.; Lau, L.; Buckley, M.; McConnell, W.; Beckett, P.; Al Ali, M.; Chauhan, A.; Wilson, S.J.; et al. Tumour necrosis factor (TNFalpha) as a novel therapeutic target in symptomatic corticosteroid dependent asthma. Thorax 2005, 60, 1012–1018. [Google Scholar] [CrossRef]
- Majak, P. Tumor necrosis factor alpha as an asthma biomarker in early childhood. Pneumonol. Alergol. Pol. 2016, 84, 143–144. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brown, S.D.; Brown, L.A.; Stephenson, S.; Dodds, J.C.; Douglas, S.L.; Qu, H.; Fitzpatrick, A.M. Characterization of a high TNF-alpha phenotype in children with moderate-to-severe asthma. J. Allergy Clin. Immunol. 2015, 135, 1651–1654. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Huang, Y.; Li, F.; Li, H.; Zhang, B.; Jin, L. MicroRNA-874 inhibits TNF-alpha-induced remodeling in human fetal airway smooth muscle cells by targeting STAT3. Respir. Physiol. Neurobiol. 2018, 251, 34–40. [Google Scholar] [CrossRef]
- Li, D.; Chen, D.; Zhang, X.; Wang, H.; Song, Z.; Xu, W.; He, Y.; Yin, Y.; Cao, J. c-Jun N-terminal kinase and Akt signalling pathways regulating tumour necrosis factor-alpha-induced interleukin-32 expression in human lung fibroblasts: Implications in airway inflammation. Immunology 2015, 144, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Gu, Y.; Zhao, S.; Huo, M.; Wang, S.; Zhang, Y.; Qiao, Y.; Li, X. Anti-Inflammatory Effects of Aurantio-Obtusin from Seed of Cassia obtusifolia L. through Modulation of the NF-kappaB Pathway. Molecules 2018, 23, 3093. [Google Scholar] [CrossRef]
- Kim, S.J.; Kim, K.W.; Kim, D.S.; Kim, M.C.; Jeon, Y.D.; Kim, S.G.; Jung, H.J.; Jang, H.J.; Lee, B.C.; Chung, W.S.; et al. The protective effect of Cassia obtusifolia on DSS-induced colitis. Am. J. Chin. Med. 2011, 39, 565–577. [Google Scholar] [CrossRef]
- Zhu, L.; Yi, X.; Ma, C.; Luo, C.; Kong, L.; Lin, X.; Gao, X.; Yuan, Z.; Wen, L.; Li, R.; et al. Betulinic Acid Attenuates Oxidative Stress in the Thymus Induced by Acute Exposure to T-2 Toxin via Regulation of the MAPK/Nrf2 Signaling Pathway. Toxins 2020, 12, 540. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.; Deng, R.; Gao, S.; Jiang, Q.; Liu, R.; Li, H.; Miao, Y.; Zhai, Y.; Zhang, S.; et al. Betulinic acid attenuated bleomycin-induced pulmonary fibrosis by effectively intervening Wnt/beta-catenin signaling. Phytomedicine 2021, 81, 153428. [Google Scholar] [CrossRef]
- Oliveira-Costa, J.F.; Meira, C.S.; Neves, M.; Dos Reis, B.; Soares, M.B.P. Anti-Inflammatory Activities of Betulinic Acid: A Review. Front. Pharmacol. 2022, 13, 883857. [Google Scholar] [CrossRef]
- Kun, L.; Wang, J.Y.; Zhang, L.; Pan, Y.Y.; Chen, X.Y.; Yuan, Y. Effects of betulinic acid on synovial inflammation in rats with collagen-induced arthritis. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420945078. [Google Scholar] [CrossRef]
- Costa, J.F.; Barbosa-Filho, J.M.; Maia, G.L.; Guimaraes, E.T.; Meira, C.S.; Ribeiro-dos-Santos, R.; de Carvalho, L.C.; Soares, M.B. Potent anti-inflammatory activity of betulinic acid treatment in a model of lethal endotoxemia. Int. Immunopharmacol. 2014, 23, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Lee, D.S.; Kim, D.C.; Yoon, C.S.; Ko, W.; Oh, H.; Kim, Y.C. Anti-Inflammatory Effects and Mechanisms of Action of Coussaric and Betulinic Acids Isolated from Diospyros kaki in Lipopolysaccharide-Stimulated RAW 264.7 Macrophages. Molecules 2016, 21, 1206. [Google Scholar] [CrossRef] [PubMed]
- Meira, C.S.; Espirito Santo, R.F.D.; Dos Santos, T.B.; Orge, I.D.; Silva, D.K.C.; Guimaraes, E.T.; Aragao Franca, L.S.; Barbosa-Filho, J.M.; Moreira, D.R.M.; Soares, M.B.P. Betulinic acid derivative BA5, a dual NF-kB/calcineurin inhibitor, alleviates experimental shock and delayed hypersensitivity. Eur. J. Pharmacol. 2017, 815, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Gong, Z.; Li, X.; Ma, Q.; Wu, M.; Liu, D.; Deng, L.; Pan, D.; Liu, Q.; Wei, Z.; et al. Betulinic acid inhibits the migration and invasion of fibroblast-like synoviocytes from patients with rheumatoid arthritis. Int. Immunopharmacol. 2019, 67, 186–193. [Google Scholar] [CrossRef]
- Karagoz, A.C.; Leidenberger, M.; Hahn, F.; Hampel, F.; Friedrich, O.; Marschall, M.; Kappes, B.; Tsogoeva, S.B. Synthesis of new betulinic acid/betulin-derived dimers and hybrids with potent antimalarial and antiviral activities. Bioorg. Med. Chem. 2019, 27, 110–115. [Google Scholar] [CrossRef]
- Jingbo, W.; Aimin, C.; Qi, W.; Xin, L.; Huaining, L. Betulinic acid inhibits IL-1beta-induced inflammation by activating PPAR-gamma in human osteoarthritis chondrocytes. Int. Immunopharmacol. 2015, 29, 687–692. [Google Scholar] [CrossRef]
- Gonzalez, A.C.; Costa, T.F.; Andrade, Z.A.; Medrado, A.R. Wound healing—A literature review. An. Bras. Dermatol. 2016, 91, 614–620. [Google Scholar] [CrossRef]
- Yazarlu, O.; Iranshahi, M.; Kashani, H.R.K.; Reshadat, S.; Habtemariam, S.; Iranshahy, M.; Hasanpour, M. Perspective on the application of medicinal plants and natural products in wound healing: A mechanistic review. Pharmacol. Res. 2021, 174, 105841. [Google Scholar] [CrossRef]
- Nayak, B.S. Cecropia peltata L(Cecropiaceae) has wound-healing potential: A preclinical study in a Sprague Dawley rat model. Int. J. Low. Extrem. Wounds 2006, 5, 20–26. [Google Scholar] [CrossRef]
- Nayak, B.S.; Vinutha, B.; Geetha, B.; Sudha, B. Experimental evaluation of Pentas lanceolata flowers for wound healing activity in rats. Fitoterapia 2005, 76, 671–675. [Google Scholar] [CrossRef]
- Kviecinski, M.R.; David, I.M.; Fernandes, F.S.; Correa, M.D.; Clarinda, M.M.; Freitas, A.F.; Silva, J.D.; Gava, M.; Muller, S.D.; Florentino, D.; et al. Healing effect of Dillenia indica fruit extracts standardized to betulinic acid on ultraviolet radiation-induced psoriasis-like wounds in rats. Pharm. Biol. 2017, 55, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Ebeling, S.; Naumann, K.; Pollok, S.; Wardecki, T.; Vidal, Y.S.S.; Nascimento, J.M.; Boerries, M.; Schmidt, G.; Brandner, J.M.; Merfort, I. From a traditional medicinal plant to a rational drug: Understanding the clinically proven wound healing efficacy of birch bark extract. PLoS ONE 2014, 9, e86147. [Google Scholar] [CrossRef] [PubMed]
- Armah, F.A.; Annan, K.; Mensah, A.Y.; Amponsah, I.K.; Tocher, D.A.; Habtemariam, S. Erythroivorensin: A novel anti-inflammatory diterpene from the root-bark of Erythrophleum ivorense (A Chev.). Fitoterapia 2015, 105, 37–42. [Google Scholar] [CrossRef]
- Zhang, D.M.; Xu, H.G.; Wang, L.; Li, Y.J.; Sun, P.H.; Wu, X.M.; Wang, G.J.; Chen, W.M.; Ye, W.C. Betulinic Acid and its Derivatives as Potential Antitumor Agents. Med. Res. Rev. 2015, 35, 1127–1155. [Google Scholar] [CrossRef] [PubMed]
- Lingaraju, M.C.; Pathak, N.N.; Begum, J.; Balaganur, V.; Bhat, R.A.; Ram, M.; Kumar, D.; Kumar, D.; Tandan, S.K. Betulinic acid negates oxidative lung injury in surgical sepsis model. J. Surg. Res. 2015, 193, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.D.; Buchta, K.A.; Swenson, C.A.; Gern, J.E.; Busse, W.W. Rhinovirus-induced interferon-gamma and airway responsiveness in asthma. Am. J. Respir. Crit. Care Med. 2003, 168, 1091–1094. [Google Scholar] [CrossRef]
- Abo-Zeid, Y.; Williams, G.R.; Touabi, L.; McLean, G.R. An investigation of rhinovirus infection on cellular uptake of poly (glycerol-adipate) nanoparticles. Int. J. Pharm. 2020, 589, 119826. [Google Scholar] [CrossRef]
- Bartlett, N.W.; Slater, L.; Glanville, N.; Haas, J.J.; Caramori, G.; Casolari, P.; Clarke, D.L.; Message, S.D.; Aniscenko, J.; Kebadze, T.; et al. Defining critical roles for NF-kappaB p65 and type I interferon in innate immunity to rhinovirus. EMBO Mol. Med. 2012, 4, 1244–1260. [Google Scholar] [CrossRef]
- Winther, J.R.; Thorpe, C. Quantification of thiols and disulfides. Biochim. Biophys. Acta 2014, 1840, 838–846. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalczyk, T.; Sitarek, P.; Śliwiński, T.; Hatziantoniou, S.; Soulintzi, N.; Pawliczak, R.; Wieczfinska, J. New Data on Anti-Inflammatory and Wound Healing Potential of Transgenic Senna obtusifolia Hairy Roots: In Vitro Studies. Int. J. Mol. Sci. 2023, 24, 5906. https://doi.org/10.3390/ijms24065906
Kowalczyk T, Sitarek P, Śliwiński T, Hatziantoniou S, Soulintzi N, Pawliczak R, Wieczfinska J. New Data on Anti-Inflammatory and Wound Healing Potential of Transgenic Senna obtusifolia Hairy Roots: In Vitro Studies. International Journal of Molecular Sciences. 2023; 24(6):5906. https://doi.org/10.3390/ijms24065906
Chicago/Turabian StyleKowalczyk, Tomasz, Przemysław Sitarek, Tomasz Śliwiński, Sophia Hatziantoniou, Nikolitsa Soulintzi, Rafal Pawliczak, and Joanna Wieczfinska. 2023. "New Data on Anti-Inflammatory and Wound Healing Potential of Transgenic Senna obtusifolia Hairy Roots: In Vitro Studies" International Journal of Molecular Sciences 24, no. 6: 5906. https://doi.org/10.3390/ijms24065906
APA StyleKowalczyk, T., Sitarek, P., Śliwiński, T., Hatziantoniou, S., Soulintzi, N., Pawliczak, R., & Wieczfinska, J. (2023). New Data on Anti-Inflammatory and Wound Healing Potential of Transgenic Senna obtusifolia Hairy Roots: In Vitro Studies. International Journal of Molecular Sciences, 24(6), 5906. https://doi.org/10.3390/ijms24065906