1. Introduction
Rheumatoid arthritis (RA), a common systemic inflammatory autoimmune disease, is characterized by an inflammatory synovium and joint fluid, synovial hyperplasia, and the progressive degradation of cartilage and bone [
1]. Epidemiological data have shown that RA affects an estimated 1% of the world’s adult population [
2]. The pathogenesis of RA infection remains poorly understood and involves a complex interplay between host factors (such as genetic susceptibilities and aberrant immune responses) and environmental triggers (such as bacterial or viral infection) [
3]. Current medical treatments for RA include nonsteroidal anti-inflammatory drugs (NSAIDs), glucocorticoids, nonbiologic disease-modifying antirheumatic drugs (DMARDs), and biologic DMARDs [
4]. Despite all these efforts to provide medical therapy for RA, a significant portion of RA patients remain unremitted despite the progression of treatment [
5,
6]. Therefore, it is crucial to understand the cellular and molecular processes by which the pathology is mediated in order to develop mechanism-driven therapeutic approaches for RA.
Synovial fibroblasts (SFs) in the synovial intimal lining of the joint play a critical role in the pathogenesis of RA through aggressive proliferation and invasion, the secretion of pro-inflammatory cytokines, the formation of pannus, and the production of cartilage-degrading matrix metalloproteases (MMPs) [
7,
8]. More importantly, activated rheumatoid arthritis synovial fibroblasts (RASFs) exhibit tumor-like features, including an increased proliferation rate and pro-migratory and pro-invasive properties that contribute greatly to pannus formation and joint destruction [
9]. Thus, inhibiting the tumor-like features of RASFs may represent potential therapeutic strategies to alleviate RA. Nevertheless, the molecular signatures and biological networks that define the distinct pathophysiologic contributions of SFs in RA have not been systematically clarified.
Circular RNAs (circRNAs), a class of endogenous noncoding RNAs (ncRNAs), are covalently closed−loop molecules without 5′ caps or 3′ poly−A tails, formed via the back−splicing of pre−mRNA [
10]. Compared with linear RNAs, circRNAs exhibit higher stability and are more resistant to RNase R digestion [
11]. Accumulating evidence has demonstrated that circRNAs can competitively bind to microRNA (miRNA) and act as a “miRNA sponge” to release miRNA’s inhibitory effect on its target genes [
12]. Due to their high stability, high conservation, and tissue−specificity, circRNAs have been reported to be implicated in the pathogenesis of various diseases, especially in cancer, and correlated with tumor initiation, progression, and metastasis [
13,
14]. However, the role and underlying mechanisms of circRNAs in the proliferation, migration, and invasion of activated RASFs in RA remain elusive.
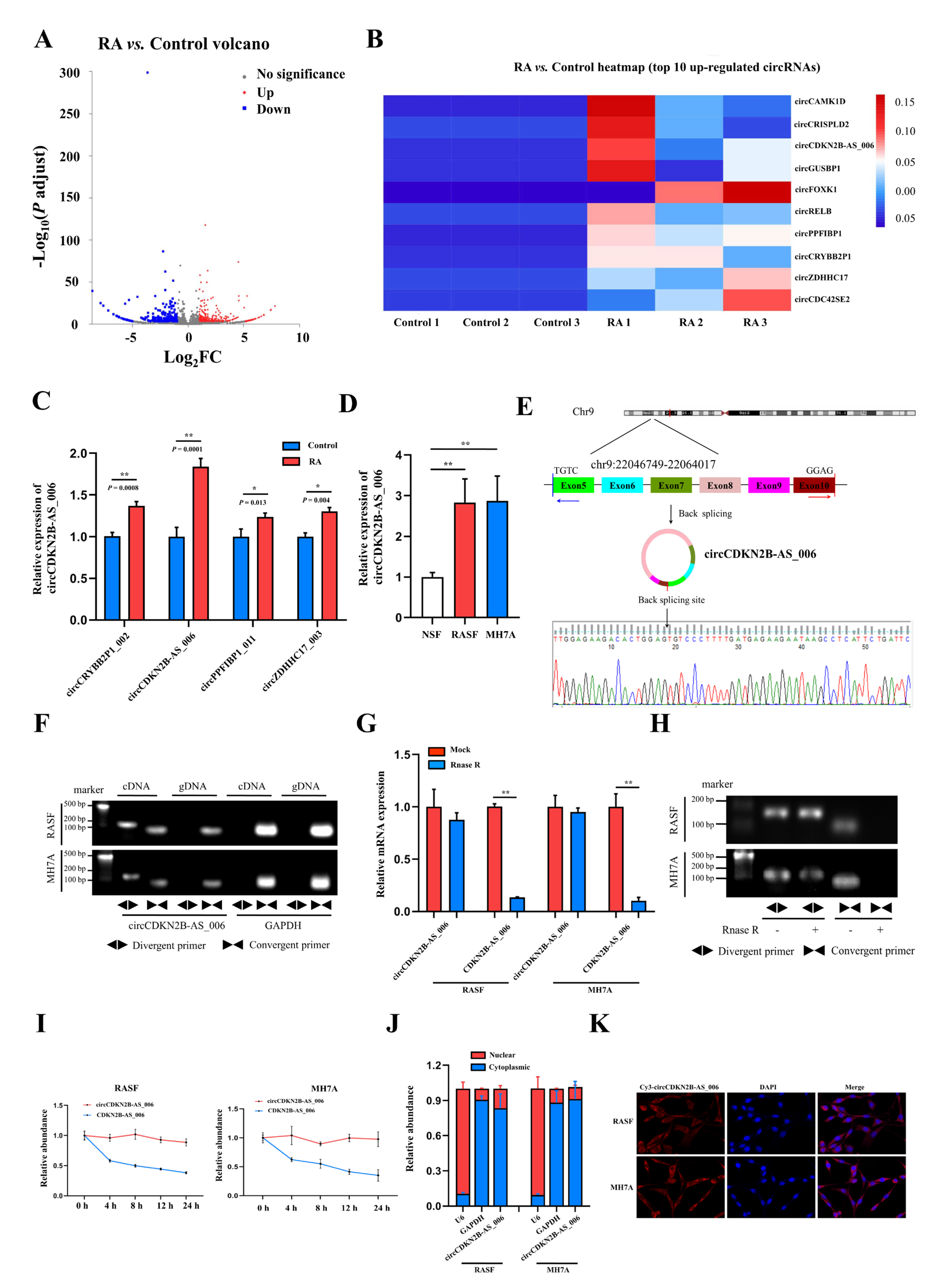
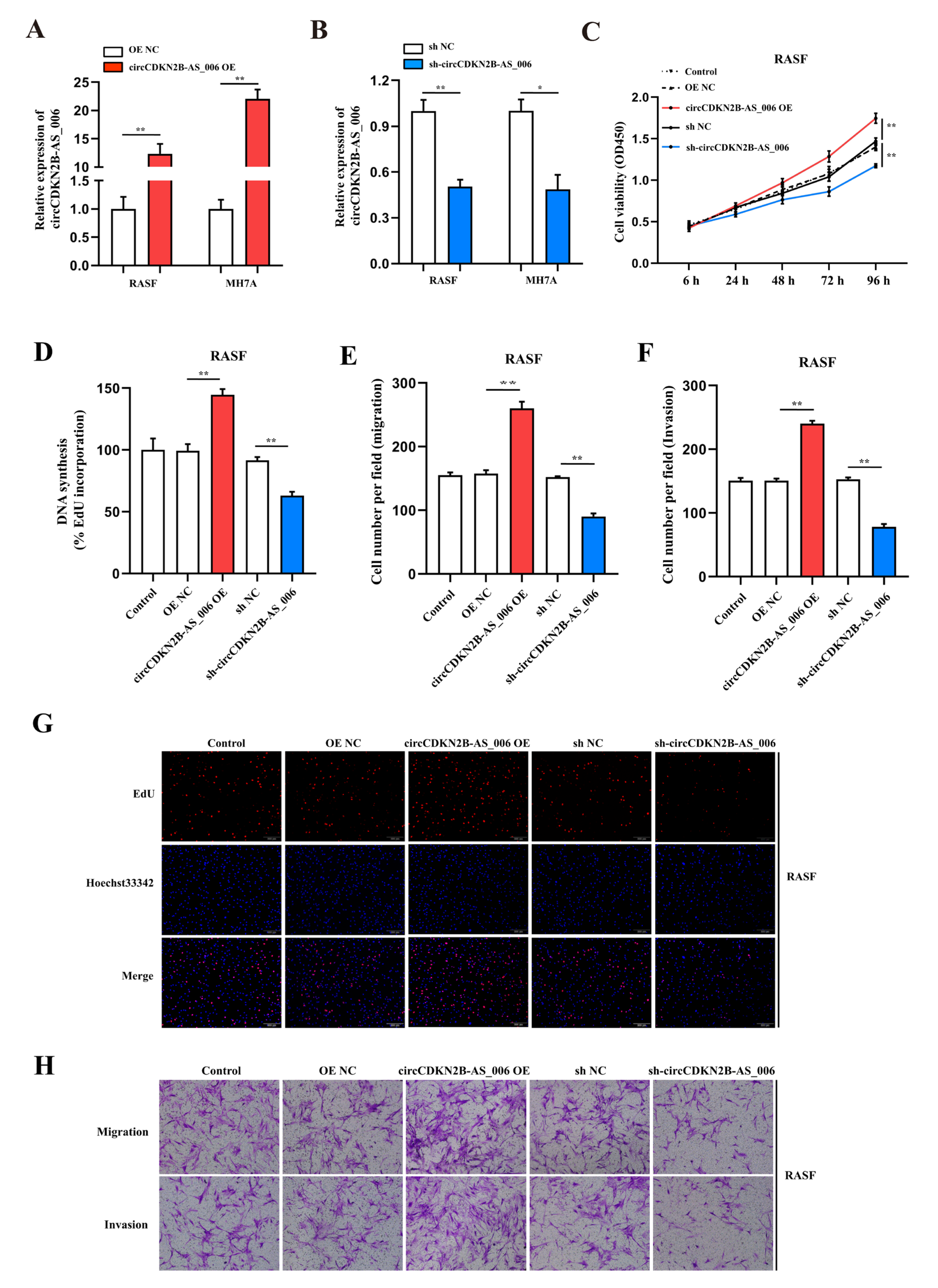
In the present study, we used whole−transcriptome sequencing to screen the differentially expressed circRNAs in the synovia from patients with RA and patients with joint trauma and identified the upregulation of hsa_circCDKN2B−AS_006. We demonstrated that circCDKN2B−AS_006 was notably upregulated in the synovium samples from patients with RA and RASFs. Functionally, circCDKN2B−AS_006 promoted the proliferation, migration, and invasion of RASFs, and circCDKN2B−AS_006 knockdown alleviated arthritis severity in a collagen−induced arthritis (CIA) mouse model. Mechanistically, circCDKN2B−AS_006 was shown to regulate the expression of runt-related transcription factor 1 (RUNX1) by sponging miR−1258, ultimately influencing the Wnt/β−catenin signaling pathway and promoting epithelial−to−mesenchymal transition (EMT) in RASFs. Clinically, circCDKN2B−AS_006, miR−1258, and RUNX1 dysregulation were shown to be related to the clinical indicators of patients with RA. Collectively, these results revealed that the circCDKN2B−AS_006/miR−1258/RUNX1 axis plays a crucial role in RASF proliferation and metastasis and, in particular, enabled us to identify circCDKN2B−AS_006 as a potential biomarker and therapeutic target in RA.
3. Discussion
Advances in high−throughput sequencing and associated bioinformatics tools and databases have extended the recognition and understanding of circRNAs. Recent studies have indicated that circRNAs may play crucial roles in the progression of RA [
22]. It has been reported that the knockdown of circRNA_09505 in macrophages significantly alleviates arthritis and inflammation in vivo in CIA mice [
23]. Another study confirmed that circ_0088036 promoted RA progression by acting as a miR−140−3p sponge to upregulate silent information regulator 1 expression [
24]. However, fewer studies have been conducted on the roles and underlying mechanisms of circRNAs in the proliferation, migration, and invasion of activated RASFs. Through a series of experiments, our investigation was the first to reveal the existence of the circCDKN2B−AS_006/miR−1258/RUNX1 axis in the progression of RA (
Figure 7E).
In the present study, we profiled circRNA expression in synovia from three pairs of RA patients and non−RA patients by means of RNA sequencing, resulting in the identification of 539 differentially expressed circRNAs. For the first time, we identified circCDKN2B−AS_006 as a significantly upregulated circRNA in synovia from RA patients. Loss− and gain−of−function experimental results suggested that circCDKN2B−AS_006 promotes the proliferation, migration, and invasion of RASFs in vitro. Moreover, circCDKN2B−AS_006 knockdown alleviated the severity of arthritis in the CIA mice model. In addition, circCDKN2B−AS_006 expression was positively correlated with ESR, serum RF, and CRP levels in patients with RA. These results indicate the critical role of circCDKN2B−AS_006 in RASF tumor-like growth and metastasis and the prognosis of RA patients.
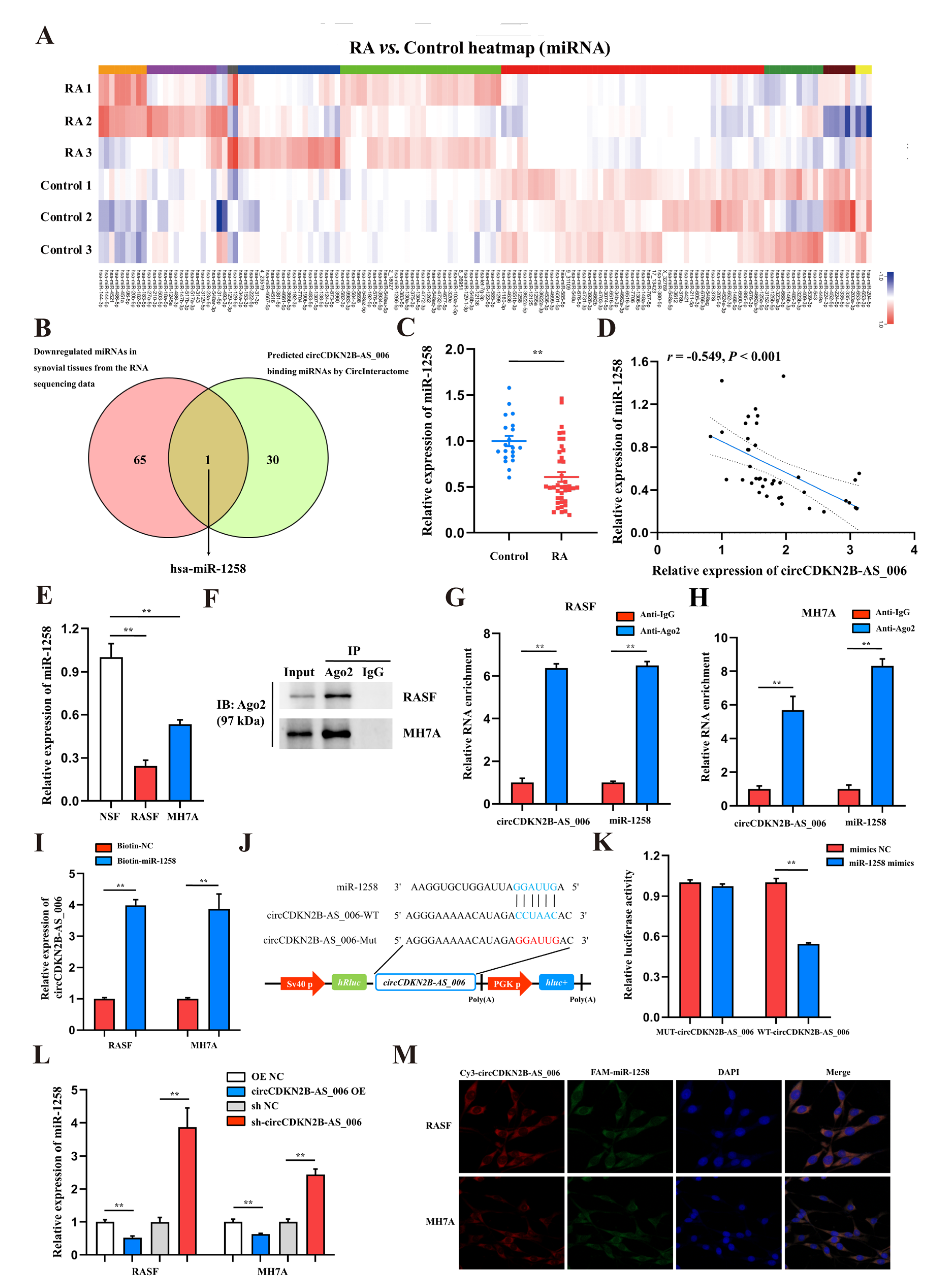
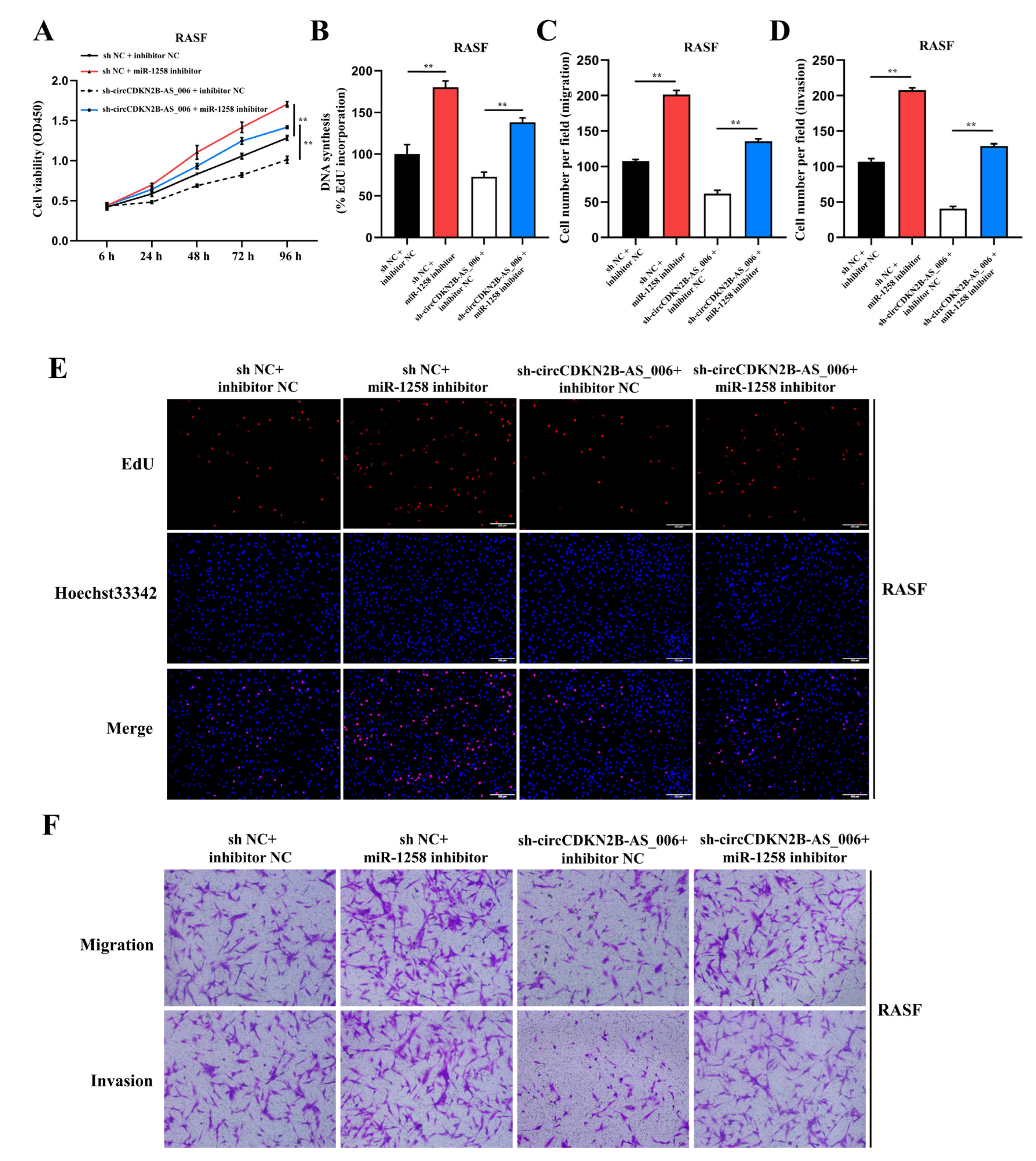
CircRNAs play vital roles in many physiological and pathological processes, including acting as microRNA (miRNA) sponges, interacting with RNA binding proteins (RBPs), and regulating transcription or translation [
25]. Emerging evidence has revealed that circRNAs could function as miRNAs [
26]. It has been reported that circLMTK2 inhibits the proliferation and metastasis of gastric cancer by acting as a competitive endogenous RNA through sponging miR-150-5p [
27]. Similarly, circBCRC−3 suppresses bladder cancer proliferation by acting as an endogenous miR−182−5p sponge, resulting in the upregulation of p27 [
28]. In the present study, through cross-analysis of a miRNA target prediction database (circular RNA interactome) and the results of whole−transcriptome sequencing (down−regulated miRNAs), we initially identified miR−1258 as a candidate target of circCDKN2B−AS_006. Pull−down, anti−Ago2 RIP, dual−luciferase, and FISH assays were conducted to validate the binding between circCDKN2B−AS_006 and miR−1258. The results indicate that, as an RISC complex, circCDKN2B−AS_006 could directly bind to the seed region of miR−1258 in the cytoplasm of RASFs and MH7A cells. Moreover, to validate whether circCDKN2B−AS_006 could exert biological functions via miR−1258, we designed rescue experiments using miR−1258 inhibitors, and the results showed that the circCDKN2B−AS_006 knockdown-induced suppression of cell proliferation, migration, and invasion was reversed through treatment with a miR−1258 inhibitor. Our results provide evidence to support the conjecture that circCDKN2B−AS_006 functions as a miRNA sponge that is important for the progression and metastasis of RASFs.
Previous studies have confirmed miR−1258 as a tumor suppressor in multiple cancers, including breast cancer [
29], hepatocellular carcinoma [
30], non−small−cell lung cancer [
16], and gastric cancer [
31]. Specifically, it has been demonstrated that miR−1258 can target heparanase and subsequently inhibit the cell−aggressive phenotype in breast cancer [
17]. Moreover, miR−1258 may play tumor−suppressive roles by targeting cyclin−dependent kinase regulatory subunit 1B expression in colorectal cancer [
32]. In the present study, we first found that miR−1258 was downregulated in the synovia from RA patients. Gain− and loss−of−function experiments showed that miR−1258 inhibited the proliferation, migration, and invasion of RASFs and MH7A cells, suggesting that miR−1258 functioned as a suppressor in RASF tumor−like growth and metastasis and the prognosis of RA patients.
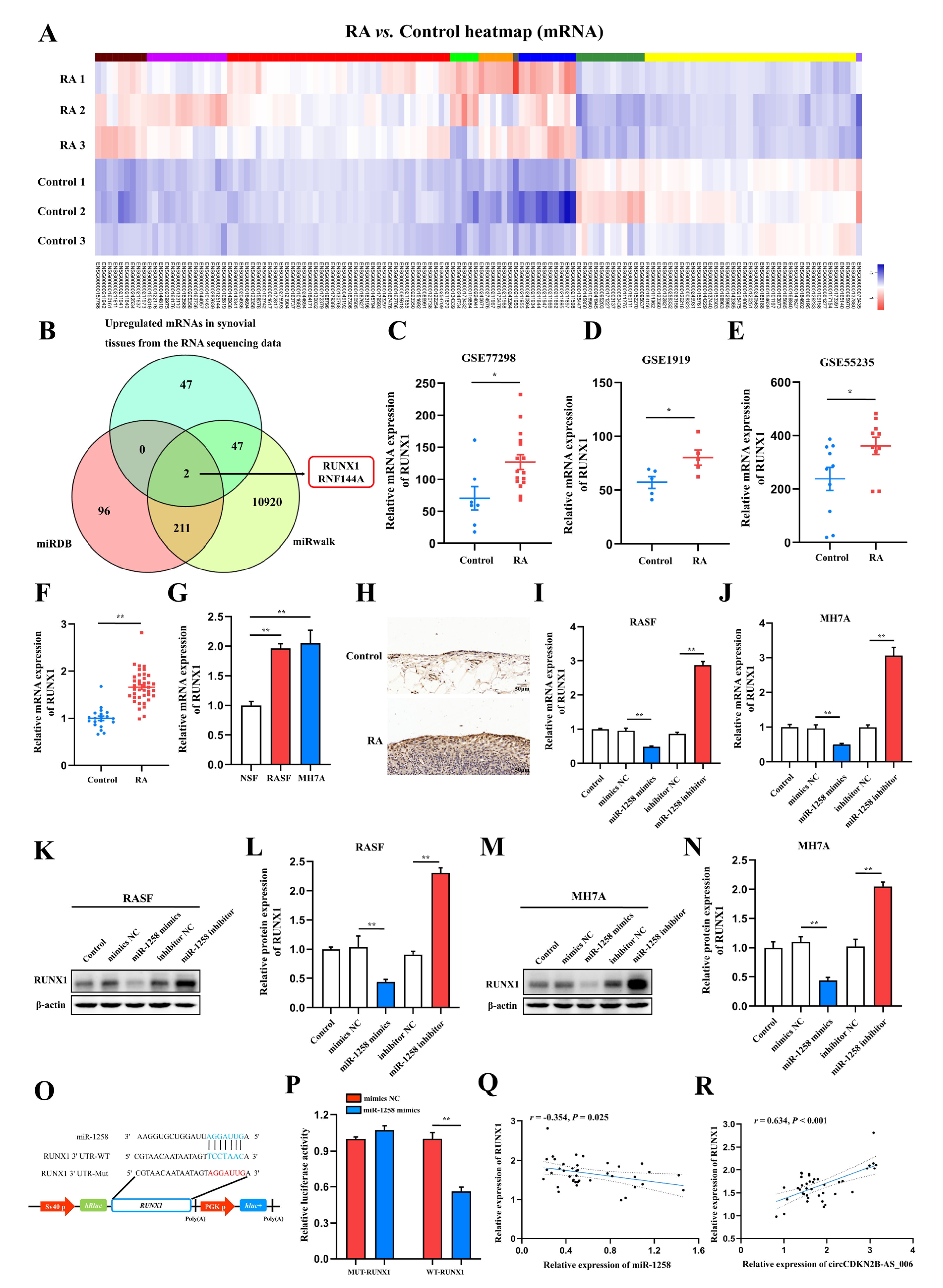
MiRNAs are small RNAs of 21−25 nucleotides that bind to partially complementary sequences in the 3′ UTR of mRNA and negatively affect post−transcriptional regulation [
33]. To further elucidate the underlying mechanism of miR−1258, we conducted bioinformatic, RT−qPCR, Western blot, and dual−luciferase analyses, with the results identifying RUNX1 as the most likely target of miR−1258. RUNX1, also called acute myeloid leukaemia 1, is a member of the RUNX family of transcription factors (RUNX1, RUNX2, and RUNX3), and this family is composed of evolutionarily conserved transcription factors that function as critical lineage determinants in various tissues [
34]. It has been reported that the suppression of RUNX1 inhibited cell proliferation, migration, and invasion in head and neck squamous cell carcinoma (HNSCC) [
35]. On the contrary, RUNX1 activation promoted colorectal cancer cell proliferation [
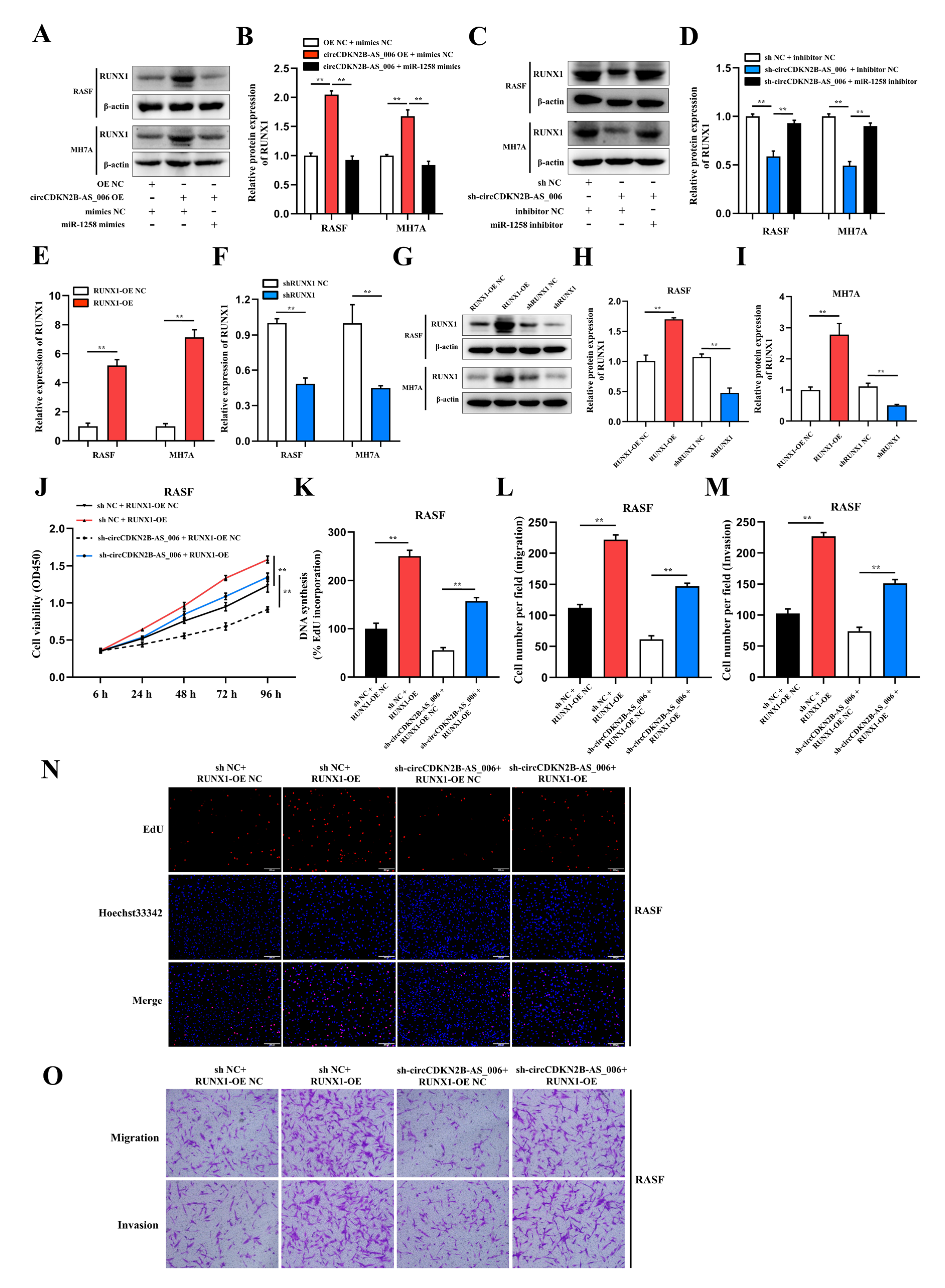
36]. In the present study, after constructing RUNX1 shRNA and overexpression lentiviral constructs, we observed that RUNX1 could remarkably promote the proliferation, migration, and invasion of RASFs and MH7A cells. Moreover, the results of rescue experiments showed that RUNX1 overexpression could mediate the inhibitory effect of circCDKN2B−AS_006 knockdown on cell proliferation, migration, and invasion. Taken together, our results revealed that circCDKN2B−AS_006 promoted RASF and MH7A proliferation, migration, and invasion via the miR−1258/RUNX1 axis.
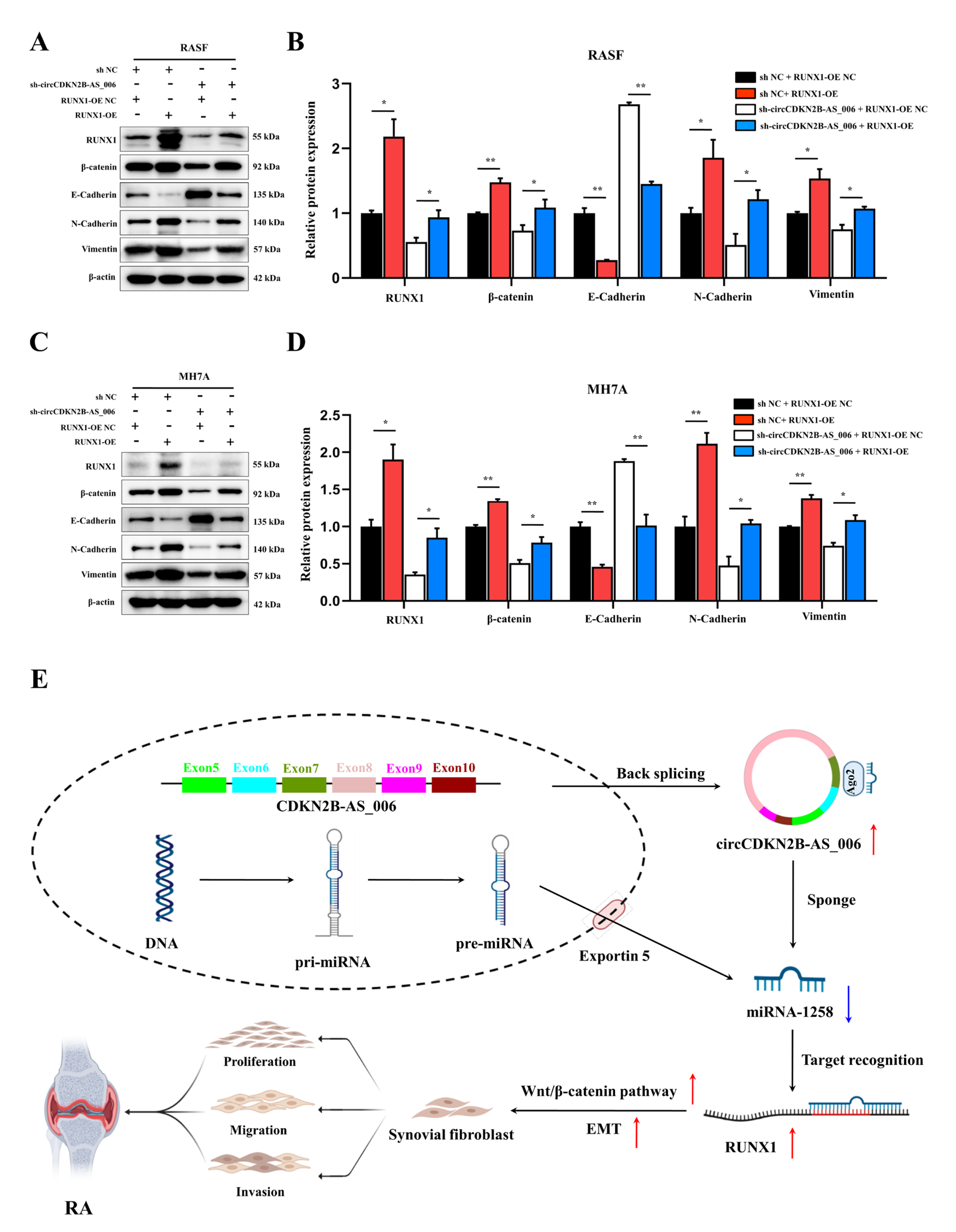
Accumulating evidence has demonstrated that RUNX1 is closely associated with the EMT process and the Wnt/β−catenin signaling pathway. A recent study showed that RUNX1 promotes colorectal cancer metastasis by activating the Wnt/β−catenin signaling pathway and EMT [
23]. Another study showed that RUNX1 promoted renal tubular EMT and kidney fibrosis by regulating TGF−β [
37]. In the present study, through bioinformatic analyses, we initially found that RUNX1 was related to the Wnt/β−catenin signaling pathway and EMT in the synovia from RA patients. Moreover, the Western blot results showed that the overexpression of RUNX1 significantly increased the levels of β−catenin and the EMT−related proteins N−cadherin and vimentin, and decreased the levels of E−cadherin. Furthermore, knockdown of circCDKN2B−AS_006 decreased the levels of β−catenin, N−cadherin, and vimentin, and increased the levels of E−cadherin, which could be reversed via RUNX1 overexpression. Collectively, these data suggest that circCDKN2B−AS_006 promotes RASF and MH7A proliferation, migration, and invasion by activating the Wnt/β−catenin signaling pathway and EMT via the miR−1258/RUNX1 axis.
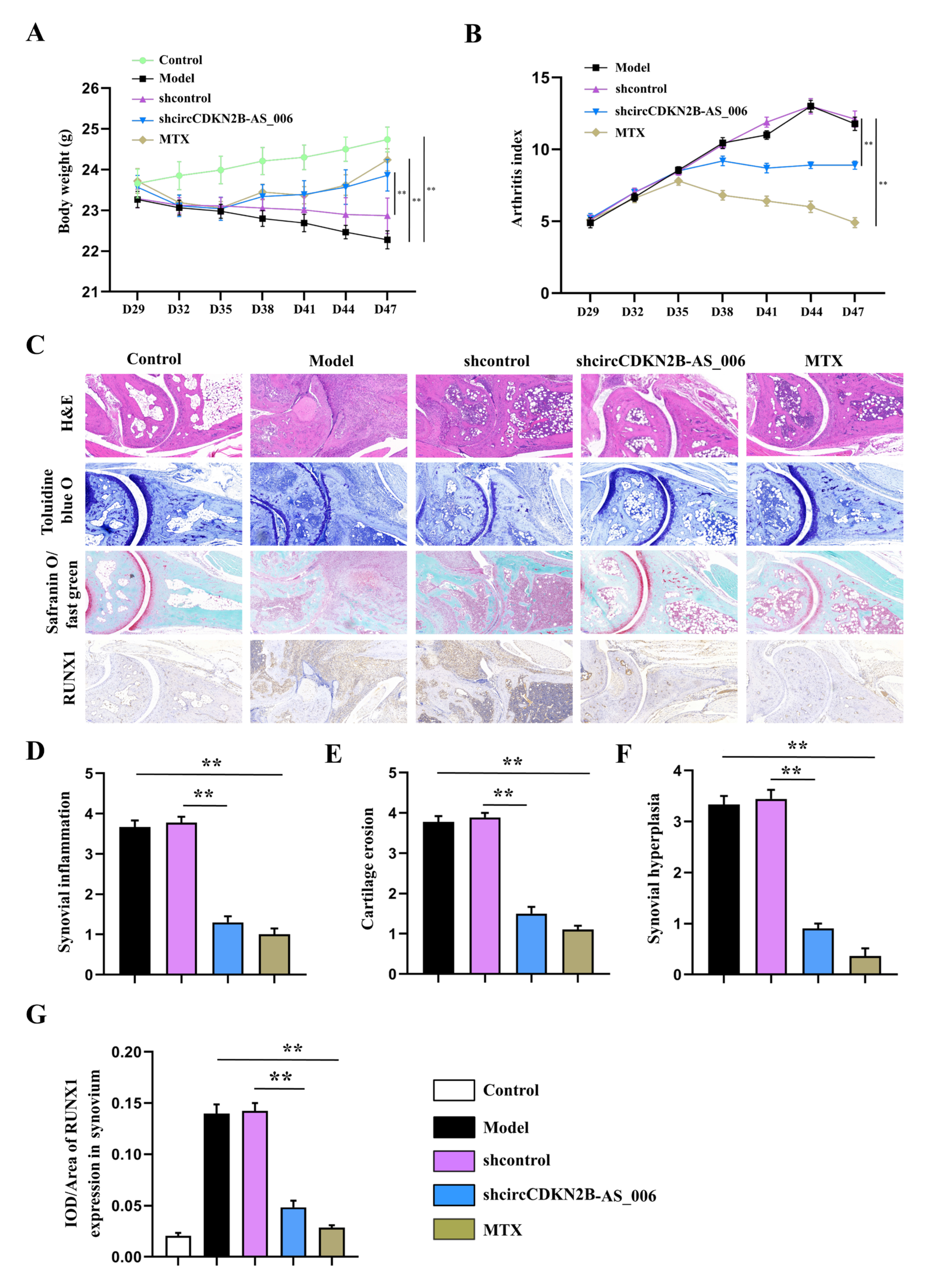
Further results revealed that the knockdown of circCDKN2B−AS_006 inhibited synovial proliferation, inflammatory cell infiltration, and articular cartilage and bone erosion in CIA mice. Immunohistochemistry staining results confirmed that downregulation of circCDKN2B−AS_006 inhibited the protein expressions of RUNX1 and β−catenin. These results indicated that knockdown of circCDKN2B−AS_006 suppressed arthritic progression with significant amelioration of joint damage via the RUNX1/β−catenin axis.
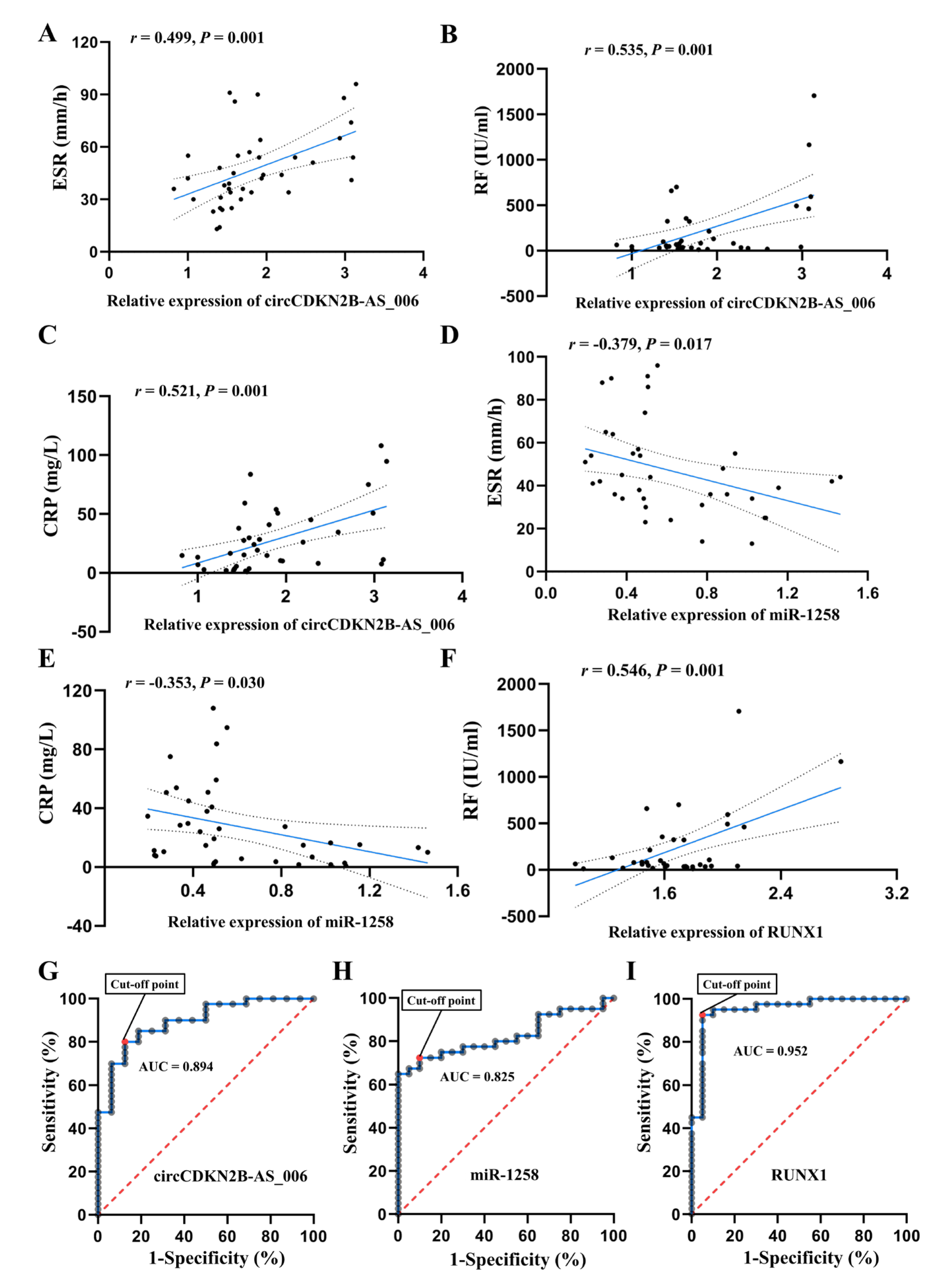
We further explored the relationship between circCDKN2B−AS_006, miR−1258, and RUNX1 expression and the clinical characteristics of RA patients. Blood biomarkers including RF, anti−cyclic citrullinated peptide antibodies (ACPA), antistreptolysin O (ASO), ESR, and CRP have shown good performance in discriminating among a portion of typical RA patients and predicting rapid progression [
38,
39,
40]. The 28−joint−count disease activity score (DAS−28) incorporates one of two inflammatory markers, ESR or CRP [
41], and is an assessment used to measure the level of disease activity in patients with RA [
42]. RF and ACPA are immunological hallmarks of RA, and the presence of these antibodies in RA is associated with higher disease activity and an increased risk of joint destruction [
43]. Therefore, clinical features including X−ray classification, time of pain, ACPA, ASO, RF, ESR, and CRP were used to evaluate the disease severity of RA in the present study. The expression of circCDKN2B−AS_006/miR−1258/RUNX1 was associated with ESR, CRP, and RF levels in serum from RA patients, suggesting a significant relationship between circCDKN2B−AS_006/miR−1258/RUNX1 expression and RA disease activity. In addition, the results of ROC curve analysis showed that the expression of circCDKN2B−AS_006/miR−1258/RUNX1 in synovium samples may represent a useful tool for discriminating patients with RA from patients with joint trauma.
Several limitations should be highlighted. Firstly, the extracellular environment of RASF in the joint cavity of RA patients is inflammatory joint fluid, while the extracellular environment of RASF in the present study is normal cell culture medium. It may be more appropriate to use freshly isolated synovial fluid from patients with RA as a culture medium for RASF. Secondly, only one of the differentially expressed circRNAs was investigated in the present study. Thirdly, patients with joint trauma were selected as control group. Given that these patients with joint trauma may have acute inflammation in the joint, normal subjects (post-mortem joint samples) may be more suitable as control group.
In summary, circCDKN2B−AS_006 was upregulated in the synovia from RA patients and was positively associated with the clinical features of RA. Functionally and mechanistically, the circCDKN2B−AS_006/miR−1258/RUNX1 axis promoted the proliferation, migration, and invasion of RASFs through the activation of the Wnt/β−catenin pathway and EMT, suggesting a potentially promising therapeutic target for RA.
4. Materials and Methods
4.1. Patients and Tissues
Patients who fulfilled the American College of Rheumatology criteria for RA were invited to enroll in the present study. The control group consisted of age− and gender−matched patients with joint trauma. Synovium samples from patients with RA (RA group; n = 40) and patients with joint trauma (control group; n = 16) were collected during joint surgery at the First Affiliated Hospital of Anhui Medical University. Patients who had a history of autoimmune or infectious disease were excluded from the control group. All participants read and signed written informed consent forms before voluntary participation. This study complied with the guidelines of the Declaration of Helsinki and was authorized by the Human Research Ethics Committee of Anhui Medical University (20210331).
4.2. RNA Extraction, Library Construction, and Sequencing
The synovium samples were submitted to the Majorbio Bio−pharm Technology Co., Ltd. (Shanghai, China) for total RNA extraction, mRNA isolation and purification, sequencing library preparation, and sequencing. Briefly, the total RNA was isolated from the synovia of RA patients or control subjects using TRIzol reagent (Invitrogen, Waltham, MA, USA) according to the manufacturer’s instructions. A NanoDrop ND−1000 spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA) was used to determine RNA purity and concentrations. A 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA) and an ND−2000 (NanoDrop Technologies, Wilmington, DE, USA) were used to determine the RNA quality. The ribosomal RNA was digested from the total RNA using the Ribo−Zero Gold kit (Illumina, San Diego, CA, USA). The libraries were then constructed and sequenced on an Illumina HiSeq xten system. Differential mRNA, miRNA, and circRNA expression analysis of the two groups was conducted via DEGseq. Fold change (FC) was regarded as an indicator of differential expression between the two groups. T−tests were utilized to evaluate the statistical significance of differences. p-values < 0.05 were considered to demonstrate differential expression.
4.3. Isolation and Culture of SF
SFs were isolated from the synovia of RA patients (RASF) or control subjects (normal synovial fibroblasts (NSFs)) as described previously [
44]. Briefly, fresh knee synovial tissues were divided into very small pieces (1 mm
3) using scissors or were minced under sterile conditions. The tissues were gently pipetted using a Pasteur pipette and attached to the wall of the cell culture flask and cultured in DMEM/high−glucose medium (Hyclone, Logan, UT, USA) containing 20% (
v/
v) fetal bovine serum (FBS) (Gibco, Grand Island, NY, USA), 100 mg/mL of streptomycin, and 100 U/mL of penicillin (both from Beyotime, Shanghai, China). The culture flask was placed upright in a 37 °C incubator in 5% CO
2 atmosphere for 6 h to allow tissue adhesion. After the formation of fibroblast−like cell colonies, adherent cells were split. All experiments were performed using synoviocyte cultures from the 4th to the 7th passages.
4.4. Cell Culture
RASFs and NSFs were cultured in DMEM/high−glucose medium (Hyclone, Logan, UT, USA) containing 20% (v/v) fetal bovine serum (FBS) (Gibco, Grand Island, NY, USA), 100 mg/mL of streptomycin, and 100 U/mL of penicillin (both from Beyotime, Shanghai, China) at 37 °C in a humidified atmosphere containing 95% air and 5% CO2.
MH7A cells, a human rheumatoid arthritis synovial cell line, were obtained from Jennio Biotech Co., Ltd. (Guangzhou, China). The cells were incubated in DMEM/high-glucose medium (Hyclone, Logan, UT, USA) containing 10% (v/v) fetal bovine serum (FBS) (Gibco, Grand Island, NY, USA), 100 mg/mL of streptomycin, and 100 U/mL of penicillin (both from Beyotime, Shanghai, China) at 37 °C in a humidified atmosphere containing 95% air and 5% CO2.
4.5. Oligonucleotides (Oligos), Plasmids, and Cell Transfection
The circCDKN2B−AS_006 plasmid, circCDKN2B−AS_006 short hairpin RNA (shRNA) plasmid, miR-1258 mimics, miR−1258 inhibitor, RUNX1 plasmid, RUNX1 shRNA plasmid, and the corresponding controls were provided by GenePharma (Shanghai, China). The circCDKN2B−AS_006 overexpression plasmid (circCDKN2B−AS_006 OE) or RUNX1 overexpression plasmid (RUNX1−OE) was generated by inserting the full−length circCDKN2B−AS_006 or RUNX1 sequence into the LV17 (EF−1a/Luciferase17&Puro) lentiviral vector or LV18C (LV18−CMV−T2A−Neo) lentiviral vector, respectively (GenePharma, Shanghai, China). The shRNA plasmid targeting the back−splice junction of circCDKN2B−AS_006 (sh−circCDKN2B−AS_006) or RUNX1 (shRUNX1) was synthesized and cloned into the LV16 (U6/Luciferase17&Puro) lentiviral vector or LV (LV−U6−copGFP−T2A−Neo) lentiviral vector, respectively (GenePharma, Shanghai, China). For the preparation of lentivirus, HEK−293 T cells were co−transfected with a lentiviral vector plus packaging plasmids using Lipofectamine 3000 reagent (Invitrogen, Carlsbad, CA, USA). RASFs or MH7A cells were infected with the packaged lentivirus and then selected with 2 μg/mL puromycin and/or 400 μg/mL geneticin (G418) (Sigma, St. Louis, MO, USA). Cell transfection with the miRNA mimics, miRNA inhibitors, or vectors was performed using Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA). All the sequences used in the present study are listed in
Table S1.
4.6. Quantitative Real−Time Polymerase Chain Reaction (RT−qPCR)
This assay was performed according to a previous study [
45]. Total RNA was obtained from the synovia and cultured cells using the TRIzol reagent (Takara, Shiga, Japan) according to the manufacturer’s instructions. Then, total RNA quantification was performed using Nanodrop 2000 (Thermo Fisher Scientific, Waltham, MA, USA) and reverse−transcribed into complementary DNA (cDNA) using a First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Waltham, MA, USA). The synthesized cDNAs were utilized for RT−qPCR, which was performed using a CFX96 real−time RT−PCR detection system (Bio−Rad, Hercules, CA, USA). The RT−qPCR analyses for mRNAs and circRNAs were conducted with a SYBR Premix Ex Taq kit (TaKaRa Biotechnology, Tokyo, Japan). For the detection of miR-1258, the Bulge−Loop™ hsa−miR−1258 RT−qPCR Primer Set (RiboBio, Guangzhou, China) and the U6 snRNA qPCR Primer Set (RiboBio, Guangzhou, China) were used for the amplification of miR−1258 and U6, respectively. The primers for miR−1258 and U6 were designed by RiboBio Inc. (Guangzhou, China), and the sequences are covered by a patent. GAPDH was used as an internal control for circRNAs and mRNAs, and U6 was employed as an endogenous control for the miRNAs. The Ct values of the samples were calculated and the transcript levels were analyzed using the 2
−ΔΔCt method.
Supplementary Table S2 shows the details of the primers used for RT−qPCR.
4.7. Western Blot Analysis
This assay was performed according to a previous study [
45]. Cells were rinsed with ice−cold phosphate-buffered saline (PBS) and lysed using radioimmunoprecipitation (RIPA) buffer containing protease inhibitors and phosphatase inhibitors. A BCA Protein Assay Kit (Beyotime Biotechnology, Shanghai, China) was used to measure the total protein concentrations. Equal amounts of protein samples were prepared in loading buffer and boiled at 100 °C for 10 min, and then the boiled samples were electrophoretically separated via sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS−PAGE) on a 10% gel and transferred onto polyvinylidene difluoride (PVDF) membranes (Millipore Corp, Billerica, MA, USA). After blocking with 5% nonfat dried milk in Tris−buffered saline (TBS) containing 1% Tween−20 for 2 h at room temperature, the membranes were incubated overnight with specific primary antibodies against Argonaute−2 (Ago2; Abcam, #ab156870), RUNX1 (LS Bio, #LS-C353932), β−catenin (Abcam, #ab223075), E−cadherin (Abcam, #ab40772), N−cadherin (Abcam, #ab76011), vimentin (Abcam, #ab92547), and β−actin (Abcam, #ab8227) at 4 °C overnight. After being washed three times with TBST, the membranes were incubated for 1 h at room temperature with horseradish peroxidase (HRP)−conjugated secondary antibodies (1:10,000 dilution). Protein bands were then detected in the enhanced chemiluminescence (ECL) detection system (Thermo Fisher Scientific Inc, Waltham, MA, USA) and quantification was performed using the ImageJ software.
4.8. RNA Fluorescence In Situ Hybridization (FISH)
The Cy3−labeled circCDKN2B−AS_006 probes and FAM−labeled miR−1258 probes were designed and synthesized by GenePharma Co. Ltd. (Shanghai, China). RASFs or MH7A cells were seeded on round coverslips, fixed with 4% paraformaldehyde for 10 min, permeabilized in PBS with 0.5% Triton X−100, and dehydrated in ethanol. FISH probes were diluted (1:50), denatured, equilibrated, and added to cells overnight at 37 °C. Then, slides were incubated with 4′,6−diamidino−2−phenylindole (DAPI) for 10 min at room temperature in the dark. After that, slides were sealed with rubber cement and air−dried in the dark for 30 min. Finally, a laser confocal microscope (LSM800, Carl Zeiss, Germany) was used to observe the fluorescence results. The probe sequences are listed in
Supplementary Table S1.
4.9. Isolation of Nuclear and Cytoplasmic Fractions
Cytosolic and nuclear fractions of cells were prepared using NE-PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Scientific, Rockford, IL, USA). Briefly, RASFs or MH7A cells were lysed in Lysis Buffer J, supplemented with protease inhibitors and centrifuged at 14,000 g for 3 min. After centrifugation, the cytoplasmic and nuclear fractions were collected from the supernatant and the pellet, respectively. Then, Buffer SK was used to extract RNA from the cytoplasmic and nuclear fraction. Subsequently, RT−qPCR was used to detect the expression of purified RNA.
4.10. Ribonuclease R (RNase R) Digestion
To verify the circCDKN2B−AS_006 characteristics, 3 μg RNA was incubated for 30 min at 37 °C with or without 3 U/μg RNase R (Epicentre Technologies, Madison, WI, USA). Then, the expression levels of circCDKN2B−AS_006 and the linear counterpart mRNA CDKN2B−AS_006 were determined by RT−qPCR and RT−PCR.
4.11. Actinomycin D Assay
RASFs or MH7A cells were seeded in a 6-well plate overnight and then treated with 2 mg/L actinomycin D (Sigma, Saint Louis, MO, USA) at the indicated time. The RT−qPCR assay was performed to assess the stability of circCDKN2B−AS_006 and CDKN2B−AS_006 after cells were harvested.
4.12. Sanger Sequencing
To further ascertain the back−splicing and junction sequence of circCDKN2B−AS_006, Sanger sequencing by Tsingke (Nanjing, China) was applied. The divergent primer (Invitrogen, Shanghai, China) was designed to confirm the back−splice junction of circCDKN2B−AS_006.
4.13. 5−Ethynyl−20−Deoxyuridine (EdU) Incorporation Assay
This assay was performed according to the method of a previous study [
45]. To assess the proliferation viability of cells, the EdU assay was carried out with a BeyoClickTM EdU−555 detection kit (Beyotime, Shanghai, China). Transfected RASFs or MH7A cells were seeded in twelve-well plates and incubated with complete medium for 12 h. After incubation with 50 mM EdU for 6 h, the cells were fixed and stained for 30 min. The nucleic acid was stained with Hoechst 33342. All images were captured with a fluorescent microscope (Olympus Optical Co., Ltd., Tokyo, Japan).
4.14. Transwell Assay
This assay was performed according to the method of a previous study [
45]. The migration and invasion abilities of RASFs or MH7A cells were assessed via transwell assays. Cells were seeded into the upper chamber, which was pre−treated with or without Matrigel (BD Biosciences, Franklin Lakes, NJ, USA), and each chamber was loaded in 100 μL serum-free culture medium and placed in 24−well tissue culture dishes. The lower chambers were filled with 600 μL DMEM containing 10% FBS. After 24 h of incubation, upper-chamber cells were removed and invaded cells were fixed and stained. All images were captured with a microscope (Olympus Optical Co., Ltd., Tokyo, Japan).
4.15. RNA−Binding Protein Immunoprecipitation (RIP) Assay
The EZ-Magna RIP RNA−binding protein immunoprecipitation kit 17−701 (Merck Millipore, Darmstadt, Germany) was used to conduct the RIP assay according to the manufacturer’s instructions. Briefly, RASFs or MH7A cells were washed with PBS and lysed in the RIP lysis buffer containing protease and ribonuclease inhibitors. The cell lysates were incubated with magnetic beads conjugated with Ago2 antibodies (Abcam, #ab156870) or IgG (Millipore, Billerica, MA, USA) at 4 °C overnight. Afterwards, beads were washed with a washing buffer and the complexes were incubated with Proteinase K for 30 min at 55 °C to remove the protein. Finally, the extracted purified RNAs were analyzed via RT−qPCR.
4.16. RNA Pull−Down Assay
GenePharma (Shanghai, China) designed and synthesized the biotinylated miR-1258 probes. The miR-1258 probe was incubated with C−1 magnetic beads (Life Technologies, Waltham, MA, USA) at 25 °C for 2 h to generate probe−coated beads. Afterwards, RASFs or MH7A cells were harvested and lysed, and the lysates were incubated with the miR−1258 probe or the negative control (NC) probes at 4 °C overnight. The RNA complexes combined with the beads were extracted using the RNeasy Mini Kit (QIAGEN). The abundances of circCDKN2B−AS_006 were evaluated via RT−qPCR. The probe sequences are listed in
Supplementary Table S1.
4.17. Luciferase Reporter Gene Assay
Luciferase reporter plasmids were obtained from GenePharma (Shanghai, China). HEK−293T cells were seeded in 24−well plates and then co−transfected with circCDKN2B−AS_006/RUNX1 wild-type or mutant plasmids and miR−1258 mimics or NC mimics using Lipofectamine 2000. At 48 h after transfection, cells were lysed and the luciferase activities were measured using the luciferase reporter assay (Promega, Madison, WI, USA), according to the manufacturer’s instructions.
4.18. Bioinformatics Analysis
Three gene expression profile datasets, GSE77298 (16 RA samples and 7 healthy controls), GSE1919 (5 RA samples and 5 healthy controls), and GSE55235 (10 RA samples and 10 healthy controls), were downloaded from the Gene Expression Omnibus (GEO) database (
https://www.ncbi.nlm.nih.gov/geo; accessed on 19 March 2022). The series matrix and platform TXT files were downloaded from the GEO database. According to the annotation information in the platform, the probes were transformed into corresponding gene symbols. If a gene symbol was recorded with multiple probes, the average value was used as its expression level. The average expression of RUNX1 mRNA in the synovia of patients with RA and normal controls were compared.
Gene set enrichment analysis (GSEA) was conducted to explore the correlation between RUNX1 expression and RA−related gene set enrichment in GSE77298, GSE55235, and GSE55584 (
http://software.broadinstitute.org/gsea/index.jsp; accessed on 6 May 2022).
Potential transcription factor RUNX1 binding sites were predicted using the known transcription factor binding site motifs from the TRANSFAC Predicted Transcription Factor Targets database. A Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis was performed using the clusterProfiler to identify the binding sites of RUNX1−enriched signaling pathways.
4.19. Animal Experiments
DBA/1 male mice (8 weeks, 20–24 g) were purchased from Beijing HFK Bioscience Co. (Beijing, China). All the animal experiments in this study were performed in strict accordance with the guidelines of the University Animal Care and Use Committee and were approved by the Animal Experimental Ethics Review Committee of Anhui Medical University (LLSC20210469).
After a week of adaptation, mice were randomly divided into 5 groups (10 mice/group) including the control group, model group, shcontrol group, shcircCDKN2B−AS_006 group, and MTX group. All mice except those in the control group were injected with complete Freund’s adjuvant (CFA) mixed with bovine collagen II (CII) (Chondrex Inc., Woodinville, WA, USA) and were given a booster injection of incomplete Freund’s adjuvant (IFA) mixed with CII on day 21 to establish the CIA mouse model. From the 28th day, mice in the shcontrol group and the shcircCDKN2B−AS_006 group were injected intra−articularly at the ankle with lentivirus stocks expressing shRNA for control or circCDKN2B−AS_006 at a concentration of 5 × 10
8 virus particles per milliliter every 4 days; mice in the control group and the model group were injected intra−articularly at the ankle with PBS every 4 days; mice in the MTX group were intragastrically administered MTX at a 1 mg/kg dose every 3 days. The severity of arthritis was evaluated as previously described [
46]. The mice were euthanized on Day 47 after the initial immunization. The ankle joints were collected and the soft tissues and muscles around the joints were removed and fixed with 4% paraformaldehyde solution for 48 h, decalcified in 10% ethylenediaminetetraacetic acid (EDTA) for 4 weeks, and embedded in paraffin. Subsequently, paraffin−embedded tissues were cut into 4 mm sections. For immunohistochemical staining, the experiment was performed using SP−9000 Histostain−Plus kits (Zsgb Bio, Beijing, China) according to the manufacturer’s protocols. Hematoxylin and eosin (H&E), toluidine blue O, and safranin O/fast green staining (Beyotime, Beijing, China) were performed according to the manufacturer’s protocols. All sections were scanned with a digital pathology slide scanner (3DHISTECH; The Digital Pathology Company, Budapest, Hungary). The average integrated optical density (IOD) of the immunohistochemical sections was calculated with Image−ProPlus software (Media Cybernetics, Silver Spring, MD, USA).
4.20. Statistical Analysis
The data were analyzed using SPSS (Statistical Package for the Social Sciences) version 17.0.1 (SPSS Inc., Chicago, IL, USA). A p-value < 0.05 was considered statistically significant, and the data are expressed as the mean ± standard error of the mean (SEM). Repeated-measures analysis of variance (ANOVA) followed by a least significant difference (LSD) test was performed to analyze the between-group effects on body weight and the polyarthritis index. Comparisons between the two groups were assessed via Student’s t−test. Comparisons between three or more groups were assessed via one−way ANOVA using an LSD post-hoc test.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








