
Molecular and Clinical Links between Drug-Induced Cholestasis and Familial Intrahepatic Cholestasis

 , , , , and
, , , , and
Abstract
1. Introduction
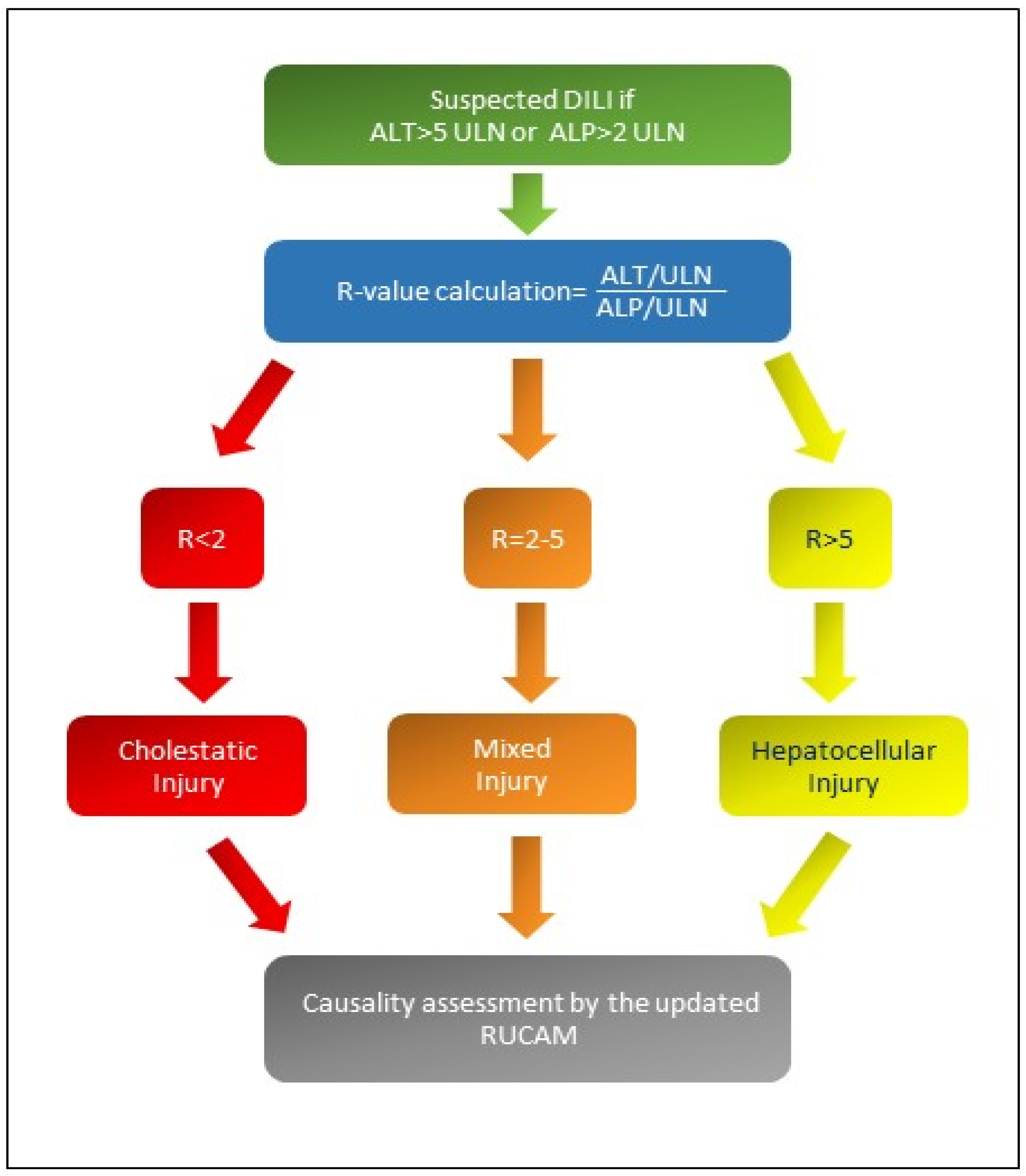
2. Aims and Literature Search
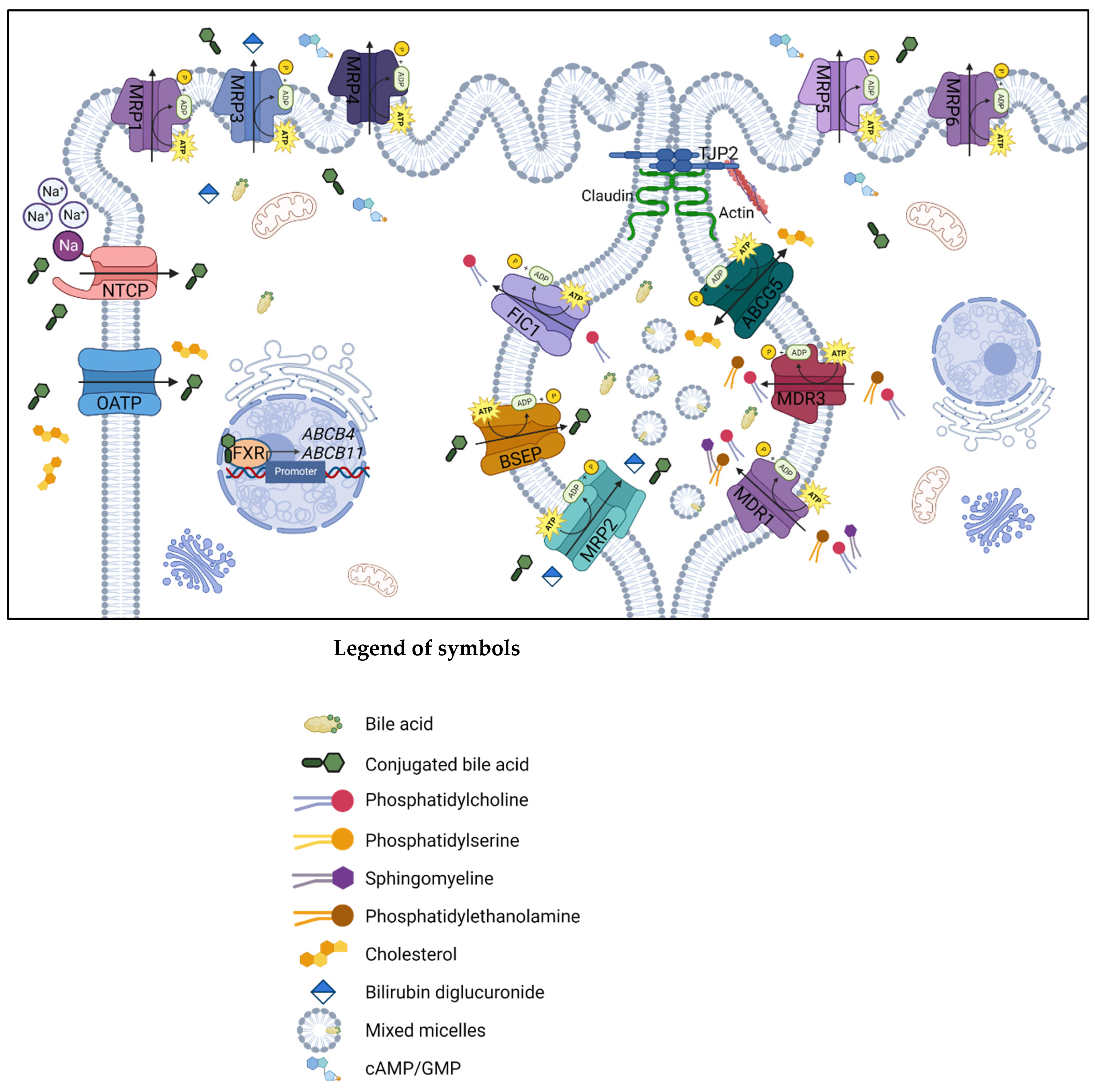
3. Molecular Bile Excretory and Metabolisms Pathways of Canalicular Efflux Transport Proteins in Drug-Induced Cholestasis
3.1. Multidrug Resistance Protein 1
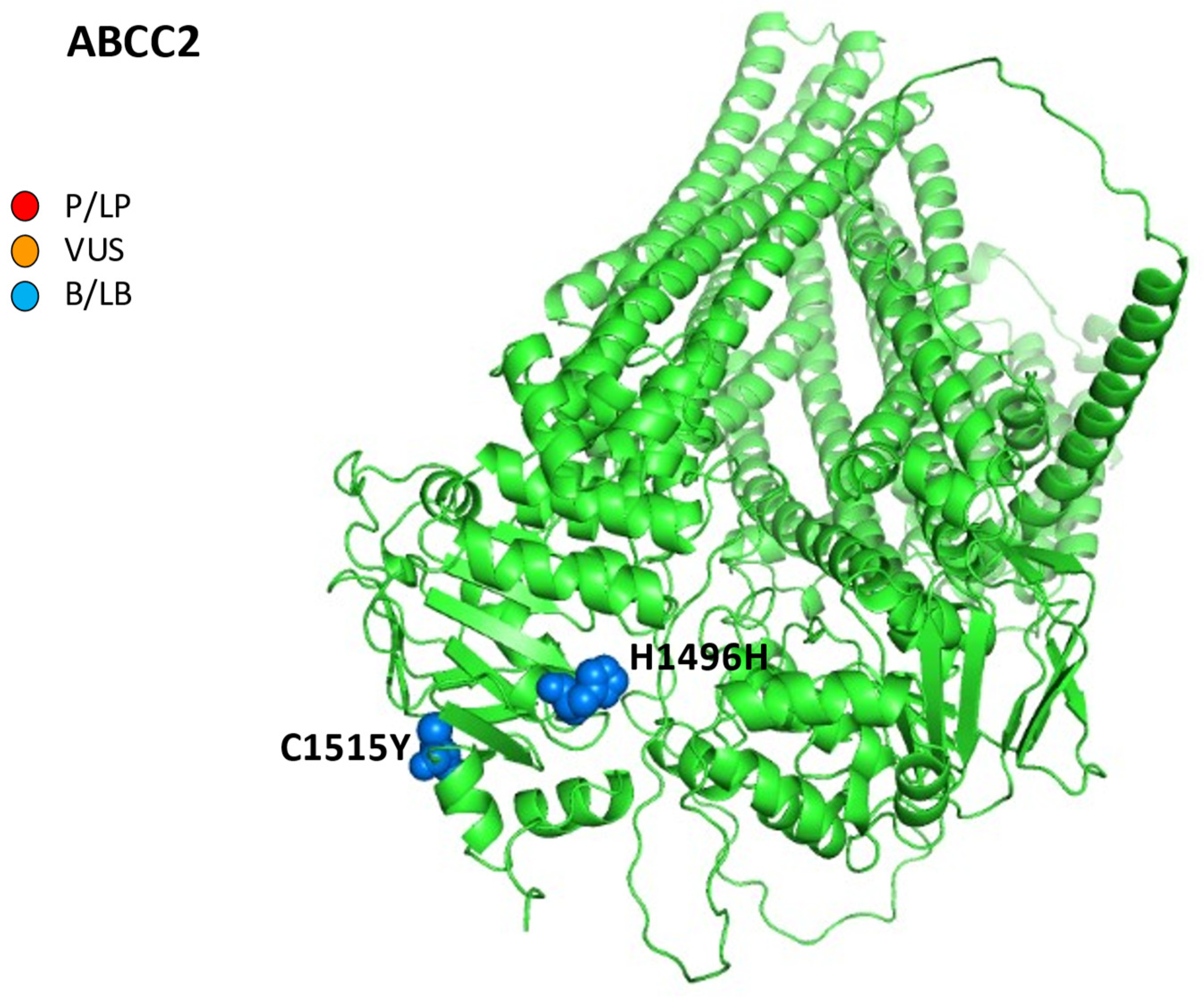
3.2. Multidrug Resistance Protein 2
3.3. Bile Salt Export Pump
3.4. Multidrug Resistance Protein 3
4. Canalicular Efflux Transport Proteins and Compounds Involved in Drug-Induced Cholestasis
4.1. Multidrug Resistance Protein 1
4.2. Multidrug Resistance Protein 2
4.3. Bile Salt Export Pump
- -
- The three causative mutations c.278A>C (p.Y93S), c.1789G>C (p.V597L), and c.3382C>T (p.R1128C) combined in a female patient (29 years) with a history of DIC, ICP, juvenile cholelithiasis, and itching;
- -
- The one homozygous mutation c.1568C>G (p.A523G) in two sisters (7 and 20 years), together with a history of neonatal jaundice, itching, and DIC (amoxicillin/clavulanic and contraceptives, respectively) [19].
4.4. MDR3
- -
- A 36-year-old woman with a background of Marfan syndrome and multiple sclerosis with cholelithiasis worsened cholestatic enzymes during dimethyl fumarate, which improved after drug discontinuation. Liver histology revealed ductopenia and minimal fibrosis without portal inflammation; a genetic test showed the pathogenic ABCB4 variants c.620T>G, p.I207R), which are likely pathogenic in the heterozygous state; a variant c.1228G>C coexisted in the CYP7A1 gene, having a pivotal role in cholesterol metabolism.
- -
- A 46-year-old man with a history of multiple endoscopic retrograde cholangiopancreatography for recurrent cholelithiasis presented jaundice after co-amoxiclav intravenous administration; a liver biopsy suggested acute cholestasis on chronic ductopenia with moderate fibrosis and incomplete septa, while the two ABCB4 variants, the intronic VUS 3507 + 10dupA and the pathogenic c.1769G>A, p.R590Q), were in the heterozygous state.
- -
- A 45-year-old woman with progressive pruritus and a history of ICP, recurrent choledocholithiasis after a cholecystectomy performed at 22 years, received Rifampicin to control her itching; her liver function tests then worsened during the treatment and improved after suspension. Next-generation sequencing revealed the ABCB4 variants: c.959C>T, p.S320F previously reported, likely pathogenic and c.2301dupT, p.T768Yfs*26 previously predicted, likely pathogenic combined to c.3149T>A p.I1050K)—VUS in ATP8B1, responsible for PFIC1. At histology, mild fibrosis, ductular reaction, ductopenia, and cholestatic rosettes were found [102].
5. Discussion
6. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABC | ATP-binding cassette | MSDs | membrane-spanning domains |
| ABCG5 | ATP-binding cassette sub-family G member 5 | NBDs | nucleotide-binding domains |
| ACMG | American College of Medical Genetics and Genomics | LPAC | low-phospholipid-associated cholelithiasis |
| AF | allele frequency | MDR | multidrug resistance protein |
| ALF | acute liver failure | MRP | multidrug resistance protein |
| ALT | alanine aminotransferase | NGS | next-generation sequencing |
| ALP | alkaline phosphatase | ||
| Anti-PD-1 | anti-programmed cell death protein 1 | NIH | National Institutes of Health |
| Anti-PDL-1 | anti-programmed death-ligand 1 | NTCP | sodium taurocholate cotransporting polypeptide |
| AP | amino phospholipids | OATP | organic anion-transporting polypeptide |
| ASBT | apical sodium-dependent bile acid transporter | OMIM | online mendelian inheritance in man |
| ATP | adenosine triphosphate | OR | odds ratio |
| B/LB | benign/likely benign | P/LP | pathogenic/likely pathogenic |
| BAs | bile acids | PBC | primary biliary cholangitis |
| BSEP | bile salt export pump | PSC | primary sclerosing cholangitis |
| BRIC | benign recurrent intrahepatic cholestasis | PDB | protein data bank |
| CCA | cholangiocarcinoma | PE | phosphatidylethanolamine |
| CI | confidence interval | PFIC | progressive familial intrahepatic cholestasis |
| DIC | drug-induced cholestasis | PMID | PubMed-Indexed for MEDLINE |
| iDILI | idiosyncratic drug-induced liver injury | PS | phosphatidylserine |
| ER | endoplasmic reticulum | RXR | retinoid X receptor |
| FIC 1 | familial intrahepatic cholestasis deficiency type 1 protein | RUCAM | Roussel Uclaf Causality Assessment Method scale |
| FXR | farnesoid X receptor | ULN | upper limit of normal |
| GWAS | genome-wide association studies | SNP | single-nucleotide polymorphism |
| HBCs | hepatobiliary cancers | TJP2 | tight junction protein 2 gene |
| HCC | hepatocellular carcinoma | TNC | transient neonatal cholestasis |
| HILI | herb-induced liver injury | VBDS | vanishing bile duct syndrome |
| HLA | human leukocyte antigen | VUS | variant at uncertain significance |
| ICP | intrahepatic cholestasis of pregnancy | ZO2 | zonula occludens-2 |
| LT | liver transplantation |
References
- Reuben, A.; Koch, D.G.; Lee, W.M. Drug-induced acute liver failure: Results of a U.S. multi-center, prospective study. Hepatology 2010, 52, 2065–2076. [Google Scholar] [CrossRef]
- Moreno-Torres, M.; Quintás, G.; Castell, J.V. The Potential Role of Metabolomics in Drug-Induced Liver Injury (DILI) Assessment. Metabolites 2022, 12, 564. [Google Scholar] [CrossRef]
- Benichou, C.; Danan, G.; Flahault, A. Causality assessment of adverse reactions to drugs–II. An original model for validation of drug causality assessment methods: Case reports with positive rechallenge. J. Clin. Epidemiol. 1993, 46, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Danan, G.; Benichou, C. Causality assessment of adverse reactions to drugs–I. A novel method based on the conclusions of international consensus meetings: Application to drug-induced liver injuries. J. Clin. Epidemiol. 1993, 46, 1323–1330. [Google Scholar] [CrossRef]
- Chalasani, N.P.; Maddur, H.; Russo, M.W.; Wong, R.J.; Reddy, K.R. Practice Parameters Committee of the American College of Gastroenterology. ACG Clinical Guideline: Diagnosis and Management of Idiosyncratic Drug-Induced Liver Injury. Am. J. Gastroenterol. 2021, 116, 878–898. [Google Scholar] [CrossRef]
- Danan, G.; Teschke, R. RUCAM in Drug and Herb Induced Liver Injury: The Update. Int. J. Mol. Sci. 2015, 17, 14. [Google Scholar] [CrossRef] [PubMed]
- Sgro, C.; Clinard, F.; Ouazir, K.; Chanay, H.; Allard, C.; Guilleminet, C.; Lenoir, C.; Lemoine, A.; Hillon, P. Incidence of drug-induced hepatic injuries: A French population-based study. Hepatology 2002, 36, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, V.; Björnsson, E.S. Drug-induced cholestasis. Hepatol. Commun. 2017, 1, 726–735. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Chalasani, N.P.; Lee, W.M.; Fontana, R.J.; Bonkovsky, H.L.; Watkins, P.B.; Hayashi, P.H.; Davern, T.J.; Navarro, V.; Reddy, R.; et al. Drug-Induced Liver Injury Network (DILIN). Hepatic histological findings in suspected drug-induced liver injury: Systematic evaluation and clinical associations. Hepatology 2014, 59, 661–670. [Google Scholar] [CrossRef]
- Berry, P.; Kotha, S.; Zen, Y.; Papa, S.; El Menabawey, T.; Webster, G.; Joshi, D.; Heneghan, M. Immune checkpoint inhibitor-related cholangiopathy: Novel clinicopathological de-scription of a multicentre cohort. Liver. Int. 2022. epub ahead of print. [Google Scholar] [CrossRef]
- Vitale, G.; Lamberti, G.; Comito, F.; Di Nunno, V.; Massari, F.; Morelli, M.C.; Ardizzoni, A.; Gelsomino, F. Anti-programmed cell death-1 and anti-programmed cell death ligand-1 immune-related liver diseases: From clinical pivotal studies to real-life experience. Expert. Opin. Biol. Ther. 2020, 20, 1047–1059. [Google Scholar] [CrossRef] [PubMed]
- Invernizzi, P. Drug-induced liver injury: Is it time for genetics to change our clinical practice? J. Hepatol. 2010, 53, 993–994. [Google Scholar] [CrossRef]
- Desmet, V.J. Vanishing bile duct syndrome in drug-induced liver disease. J. Hepatol. 1997, 26 (Suppl. 1), 31–35. [Google Scholar] [CrossRef]
- Pauli-Magnus, C.; Meier, P.J. Hepatobiliary transporters and drug-induced cholestasis. Hepatology 2006, 44, 778–787. [Google Scholar] [CrossRef]
- Vitale, G.; Gitto, S.; Vukotic, R.; Raimondi, F.; Andreone, P. Familial intrahepatic cholestasis: New and wide perspectives. Dig. Liver. Dis. 2019, 51, 922–933. [Google Scholar] [CrossRef]
- Morotti, R.A.; Suchy, F.J.; Magid, M.S. Progressive familial intrahepatic cholestasis (PFIC) type 1, 2, and 3: A review of the liver pathology findings. Semin. Liver. Dis. 2011, 31, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Dröge, C.; Bonus, M.; Baumann, U.; Klindt, C.; Lainka, E.; Kathemann, S.; Brinkert, F.; Grabhorn, E.; Pfister, E.D.; Wenning, D.; et al. Sequencing of FIC1, BSEP and MDR3 in a large cohort of patients with cholestasis revealed a high number of different genetic variants. J. Hepatol. 2017, 67, 1253–1264. [Google Scholar] [CrossRef]
- Nayagam, J.S.; Foskett, P.; Strautnieks, S.; Agarwal, K.; Miquel, R.; Joshi, D.; Thompson, R.J. Clinical phenotype of adult-onset liver disease in patients with variants in ABCB4, ABCB11, and ATP8B1. Hepatol. Commun. 2022, 6, 2654–2664. [Google Scholar] [CrossRef] [PubMed]
- Vitale, G.; Gitto, S.; Raimondi, F.; Mattiaccio, A.; Mantovani, V.; Vukotic, R.; D’Errico, A.; Seri, M.; Russell, R.B.; Andreone, P. Cryptogenic cholestasis in young and adults: ATP8B1, ABCB11, ABCB4, and TJP2 gene variants analysis by high-throughput sequencing. J. Gastroenterol. 2018, 53, 945–958. [Google Scholar] [CrossRef] [PubMed]
- Nayagam, J.S.; Williamson, C.; Joshi, D.; Thompson, R.J. Review article: Liver disease in adults with variants in the cholestasis-related genes ABCB11, ABCB4 and ATP8B1. Aliment. Pharmacol. Ther. 2020, 52, 1628–1639. [Google Scholar] [CrossRef]
- Vitale, G.; Mattiaccio, A.; Conti, A.; Turco, L.; Seri, M.; Piscaglia, F.; Morelli, M.C. Genetics in Familial Intrahepatic Cholestasis: Clinical Patterns and Development of Liver and Biliary Cancers: A Review of the Literature. Cancers 2022, 14, 3421. [Google Scholar] [CrossRef] [PubMed]
- Stättermayer, A.F.; Halilbasic, E.; Wrba, F.; Ferenci, P.; Trauner, M. Variants in ABCB4 (MDR3) across the spectrum of cholestatic liver diseases in adults. J. Hepatol. 2020, 73, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Ahn, B.M.; Yi, J.; Lee, J.H.; Lee, J.H.; Nam, S.W.; Chon, C.Y.; Han, K.H.; Ahn, S.H.; Jang, I.J.; et al. MRP2 haplotypes confer differential susceptibility to toxic liver injury. Pharmacogenetics Genom. 2007, 17, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Kodan, A.; Futamata, R.; Kimura, Y.; Kioka, N.; Nakatsu, T.; Kato, H.; Ueda, K. ABCB1/MDR1/P-gp employs an ATP-dependent twist-and-squeeze mechanism to export hydrophobic drugs. FEBS Lett. 2021, 595, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hou, W.T.; Wang, J.; Xu, D.; Guo, C.; Sun, L.; Ruan, K.; Zhou, C.Z.; Chen, Y. Structures of human bile acid exporter ABCB11 reveal a transport mechanism facilitated by two tandem substrate-binding pockets. Cell Res. 2022, 32, 501–504. [Google Scholar] [CrossRef]
- Olsen, J.A.; Alam, A.; Kowal, J.; Stieger, B.; Locher, K.P. Structure of the human lipid exporter ABCB4 in a lipid environment. Nat. Struct. Mol. Biol. 2020, 27, 62–70. [Google Scholar] [CrossRef]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. ACMG Laboratory Quality Assurance Committee. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef]
- Huang, Y.S.; Chang, T.E.; Perng, C.L.; Huang, Y.H. The association of transporter ABCC2 (MRP2) genetic variation and drug-induced hyperbilirubinemia. J. Chin. Med. Assoc. 2021, 84, 129–135. [Google Scholar] [CrossRef]
- Wang, R.B.; Kuo, C.L.; Lien, L.L.; Lien, E.J. Structure-activity relationship: Analyses of p-glycoprotein substrates and inhibitors. J. Clin. Pharm. Ther. 2003, 28, 203–228. [Google Scholar] [CrossRef]
- Ambudkar, S.V.; Dey, S.; Hrycyna, C.A.; Ramachandra, M.; Pastan, I.; Gottesman, M.M. Biochemical, cellular, and pharmacological aspects of the multidrug transporter. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 361–398. [Google Scholar] [CrossRef]
- Cascorbi, I. P-glycoprotein: Tissue distribution, substrates, and functional consequences of genetic variations. Handb. Exp. Pharmacol. 2011, 201, 261–283. [Google Scholar] [CrossRef]
- Haas, D.W.; Bartlett, J.A.; Andersen, J.W.; Sanne, I.; Wilkinson, G.R.; Hinkle, J.; Rousseau, F.; Ingram, C.D.; Shaw, A.; Lederman, M.M.; et al. Adult AIDS Clinical Trials Group. Pharmacogenetics of nevirapine-associated hepatotoxicity: An Adult AIDS Clinical Trials Group collaboration. Clin. Infect. Dis. 2006, 43, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.D.; Haas, D.W.; Motsinger, A.A.; Donahue, J.P.; Erdem, H.; Raffanti, S.; Rebeiro, P.; George, A.L.; Kim, R.B.; Haines, J.L.; et al. Drug transporter and metabolizing enzyme gene variants and nonnucleoside reverse-transcriptase inhibitor hepatotoxicity. Clin. Infect. Dis. 2006, 43, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Ciccacci, C.; Borgiani, P.; Ceffa, S.; Sirianni, E.; Marazzi, M.C.; Altan, A.M.; Paturzo, G.; Bramanti, P.; Novelli, G.; Palombi, L. Nevirapine-induced hepatotoxicity and pharmacogenetics: A retrospective study in a population from Mozambique. Pharmacogenomics 2010, 11, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Borst, P.; Zelcer, N.; van de Wetering, K. MRP2 and 3 in health and disease. Cancer Lett. 2006, 234, 51–61. [Google Scholar] [CrossRef]
- Alnouti, Y. Bile Acid sulfation: A pathway of bile acid elimination and detoxification. Toxicol. Sci. 2009, 108, 225–246. [Google Scholar] [CrossRef]
- Zollner, G.; Trauner, M. Molecular mechanisms of cholestasis. Wien. Med. Wochenschr. 2006, 156, 380–385. [Google Scholar] [CrossRef]
- Nies, A.T.; Keppler, D. The apical conjugate efflux pump ABCC2 (MRP2). Pflugers. Arch. 2007, 453, 643–659. [Google Scholar] [CrossRef]
- Geier, A.; Wagner, M.; Dietrich, C.G.; Trauner, M. Principles of hepatic organic anion transporter regulation during cholestasis, inflammation and liver regeneration. Biochim. Biophys. Acta 2007, 1773, 283–308. [Google Scholar] [CrossRef]
- Paulusma, C.C.; Kool, M.; Bosma, P.J.; Scheffer, G.L.; ter Borg, F.; Scheper, R.J.; Tytgat, G.N.; Borst, P.; Baas, F.; Oude Elferink, R.P. A mutation in the human canalicular multispecific organic anion transporter gene causes the Dubin-Johnson syndrome. Hepatology 1997, 25, 1539–1542. [Google Scholar] [CrossRef] [PubMed]
- Dawson, P.A.; Lan, T.; Rao, A. Bile acid transporters. J. Lipid. Res. 2009, 50, 2340–2357. [Google Scholar] [CrossRef] [PubMed]
- Stieger, B.; Meier, Y.; Meier, P.J. The bile salt export pump. Pflugers. Arch. 2007, 453, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Stieger, B. Recent insights into the function and regulation of the bile salt export pump (ABCB11). Curr. Opin. Lipidol. 2009, 20, 176–181. [Google Scholar] [CrossRef]
- Soroka, C.J.; Boyer, J.L. Biosynthesis and trafficking of the bile salt export pump, BSEP: Therapeutic implications of BSEP mutations. Mol. Aspects. Med. 2014, 37, 3–14. [Google Scholar] [CrossRef]
- Baker, A.; Kerkar, N.; Todorova, L.; Kamath, B.M.; Houwen, R.H.J. Systematic review of progressive familial intrahepatic cholestasis. Clin. Res. Hepatol. Gastroenterol. 2019, 43, 20–36. [Google Scholar] [CrossRef]
- Pauli-Magnus, C.; Stieger, B.; Meier, Y.; Kullak-Ublick, G.A.; Meier, P.J. Enterohepatic transport of bile salts and genetics of cholestasis. J. Hepatol. 2005, 43, 342–357. [Google Scholar] [CrossRef] [PubMed]
- Noe, J.; Kullak-Ublick, G.A.; Jochum, W.; Stieger, B.; Kerb, R.; Haberl, M.; Müllhaupt, B.; Meier, P.J.; Pauli-Magnus, C. Impaired expression and function of the bile salt export pump due to three novel ABCB11 mutations in intrahepatic cholestasis. J. Hepatol. 2005, 43, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Morgan, R.E.; Trauner, M.; van Staden, C.J.; Lee, P.H.; Ramachandran, B.; Eschenberg, M.; Afshari, C.A.; Qualls, C.W., Jr.; Lightfoot-Dunn, R.; Hamadeh, H.K. Interference with bile salt export pump function is a susceptibility factor for human liver injury in drug development. Toxicol. Sci. 2010, 118, 485–500. [Google Scholar] [CrossRef]
- Morgan, R.E.; van Staden, C.J.; Chen, Y.; Kalyanaraman, N.; Kalanzi, J.; Dunn RT 2nd Afshari, C.A.; Hamadeh, H.K. A multifactorial approach to hepatobiliary transporter assessment enables improved therapeutic compound development. Toxicol. Sci. 2013, 136, 216–241. [Google Scholar] [CrossRef]
- Prince, M.I.; Burt, A.D.; Jones, D.E. Hepatitis and liver dysfunction with rifampicin therapy for pruritus in primary biliary cirrhosis. Gut 2002, 50, 436–439. [Google Scholar] [CrossRef]
- Lang, C.; Meier, Y.; Stieger, B.; Beuers, U.; Lang, T.; Kerb, R.; Kullak-Ublick, G.A.; Meier, P.J.; Pauli-Magnus, C. Mutations and polymorphisms in the bile salt export pump and the multidrug resistance protein 3 associated with drug-induced liver injury. Pharm. Genom. 2007, 17, 47–60. [Google Scholar] [CrossRef]
- Fattinger, K.; Funk, C.; Pantze, M.; Weber, C.; Reichen, J.; Stieger, B.; Meier, P.J. The endothelin antagonist bosentan inhibits the canalicular bile salt export pump: A potential mechanism for hepatic adverse reactions. Clin. Pharmacol. Ther. 2001, 69, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Funk, C.; Pantze, M.; Jehle, L.; Ponelle, C.; Scheuermann, G.; Lazendic, M.; Gasser, R. Troglitazone-induced intrahepatic cholestasis by an interference with the hepatobiliary export of bile acids in male and female rats. Correlation with the gender difference in troglitazone sulfate formation and the inhibition of the canalicular bile salt export pump (Bsep) by troglitazone and troglitazone sulfate. Toxicology 2001, 167, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Stieger, B.; Fattinger, K.; Madon, J.; Kullak-Ublick, G.A.; Meier, P.J. Drug- and estrogen-induced cholestasis through inhibition of the hepatocellular bile salt export pump (Bsep) of rat liver. Gastroenterology 2000, 118, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Dean, M.; Rzhetsky, A.; Allikmets, R. The human ATP-binding cassette (ABC) transporter superfamily. Genome. Res. 2001, 11, 1156–1166. [Google Scholar] [CrossRef]
- Zolnerciks, J.K.; Andress, E.J.; Nicolaou, M.; Linton, K.J. Structure of ABC transporters. Essays. Biochem. 2011, 50, 43–61. [Google Scholar] [CrossRef]
- Morita, S.Y.; Terada, T. Molecular mechanisms for biliary phospholipid and drug efflux mediated by ABCB4 and bile salts. Biomed. Res. Int. 2014, 2014, 954781. [Google Scholar] [CrossRef] [PubMed]
- Groen, A.; Romero, M.R.; Kunne, C.; Hoosdally, S.J.; Dixon, P.H.; Wooding, C.; Williamson, C.; Seppen, J.; Van den Oever, K.; Mok, K.S.; et al. Complementary functions of the flippase ATP8B1 and the floppase ABCB4 in maintaining canalicular membrane integrity. Gastroenterology 2011, 141, 1927–1937.e1-4. [Google Scholar] [CrossRef]
- Schatz, S.B.; Jüngst, C.; Keitel-Anselmo, V.; Kubitz, R.; Becker, C.; Gerner, P.; Pfister, E.D.; Goldschmidt, I.; Junge, N.; Wenning, D.; et al. Phenotypic spectrum and diagnostic pitfalls of ABCB4 deficiency depending on age of onset. Hepatol. Commun. 2018, 2, 504–514. [Google Scholar] [CrossRef]
- Delaunay, J.L.; Durand-Schneider, A.M.; Dossier, C.; Falguières, T.; Gautherot, J.; Davit-Spraul, A.; Aït-Slimane, T.; Housset, C.; Jacquemin, E.; Maurice, M. A functional classification of ABCB4 variations causing progressive familial intrahepatic cholestasis type 3. Hepatology 2016, 63, 1620–1631. [Google Scholar] [CrossRef]
- Kubitz, R.; Bode, J.; Erhardt, A.; Graf, D.; Kircheis, G.; Müller-Stöver, I.; Reinehr, R.; Reuter, S.; Richter, J.; Sagir, A.; et al. Cholestatic liver diseases from child to adult: The diversity of MDR3 disease. Z Gastroenterol. 2011, 49, 728–736. [Google Scholar] [CrossRef]
- Gudbjartsson, D.F.; Helgason, H.; Gudjonsson, S.A.; Zink, F.; Oddson, A.; Gylfason, A. Large-scale whole-genome sequencing of the Icelandic population. Nat. Genet. 2015, 47, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Mhatre, S.; Wang, Z.; Nagrani, R.; Badwe, R.; Chiplunkar, S.; Mittal, B.; Yadav, S.; Zhang, H.; Chung, C.C.; Patil, P.; et al. Common genetic variation and risk of gallbladder cancer in India: A case-control genome-wide association study. Lancet. Oncol. 2017, 18, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Reichert, M.C.; Lammert, F. ABCB4 gene aberrations in human liver disease: An evolving spectrum. Semin. Liver. Dis. 2018, 38, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Zollner, G.; Thueringer, A.; Lackner, C.; Fickert, P.; Trauner, M. Alterations of canalicular ATP-binding cassette transporter expression in drug-induced liver injury. Digestion 2014, 90, 81–88. [Google Scholar] [CrossRef]
- Ros, J.E.; Libbrecht, L.; Geuken, M.; Jansen, P.L.; Roskams, T.A. High expression of MDR1, MRP1, and MRP3 in the hepatic progenitor cell compartment and hepatocytes in severe human liver disease. J. Pathol. 2003, 200, 553–560. [Google Scholar] [CrossRef]
- Saab, L.; Peluso, J.; Muller, C.D.; Ubeaud-Sequier, G. Implication of hepatic transporters (MDR1 and MRP2) in inflammation-associated idiosyncratic drug-induced hepatotoxicity investigated by microvolume cytometry. Cytom. A 2013, 83, 403–408. [Google Scholar] [CrossRef]
- Fukunaga, K.; Nakagawa, H.; Ishikawa, T.; Kubo, M.; Mushiroda, T. ABCB1 polymorphism is associated with atorvastatin-induced liver injury in Japanese population. BMC Genet. 2016, 17, 79. [Google Scholar] [CrossRef]
- Fromm, M.F. Importance of P-glycoprotein at blood-tissue barriers. Trends. Pharmacol. Sci. 2004, 25, 423–429. [Google Scholar] [CrossRef]
- Kimura, Y.; Morita, S.Y.; Matsuo, M.; Ueda, K. Mechanism of multidrug recognition by MDR1/ABCB1. Cancer Sci. 2007, 98, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kim, S.H.; Lee, J.H.; Lee, B.H.; Kim, Y.S.; Park, J.S.; Jee, Y.K. Polymorphisms in drug transporter genes (ABCB1, SLCO1B1 and ABCC2) and hepatitis induced by antituberculosis drugs. Tuberculosis (Edinb.) 2012, 92, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Wu, T.; Jiao, L.; Wu, Q.; Zhao, Z.; Song, J.; Liu, T.; Lv, Y.; Lu, X.; Ying, B. Association of ABCC Gene Polymorphism With Susceptibility to Antituberculosis Drug-Induced Hepatotoxicity in Western Han Patients With Tuberculosis. J. Clin. Pharmacol. 2020, 60, 361–368. [Google Scholar] [CrossRef]
- Yuan, J.; Guo, S.; Hall, D.; Cammett, A.M.; Jayadev, S.; Distel, M.; Storfer, S.; Huang, Z.; Mootsikapun, P.; Ruxrungtham, K.; et al. Nevirapine Toxicogenomics Study Team. Toxicogenomics of nevirapine-associated cutaneous and hepatic adverse events among populations of African, Asian, and European descent. AIDS 2011, 25, 1271–1280. [Google Scholar] [CrossRef]
- Nicolaou, M.; Andress, E.J.; Zolnerciks, J.K.; Dixon, P.H.; Williamson, C.; Linton, K.J. Canalicular ABC transporters and liver disease. J. Pathol. 2012, 226, 300–315. [Google Scholar] [CrossRef]
- Daly, A.K.; Aithal, G.P.; Leathart, J.B.; Swainsbury, R.A.; Dang, T.S.; Day, C.P. Genetic susceptibility to diclofenac-induced hepatotoxicity: Contribution of UGT2B7, CYP2C8, and ABCC2 genotypes. Gastroenterology 2007, 132, 272–281. [Google Scholar] [CrossRef]
- Fouassier, L.; Kinnman, N.; Lefèvre, G.; Lasnier, E.; Rey, C.; Poupon, R.; Elferink, R.P.; Housset, C. Contribution of mrp2 in alterations of canalicular bile formation by the endothelin antagonist bosentan. J. Hepatol. 2002, 37, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.Y.; Tang, X.D.; Chen, J.; Wu, H.B.; Chen, W.S.; Chen, L. Rifampicin induces clathrin-dependent endocytosis and ubiquitin-proteasome degradation of MRP2 via oxidative stress-activated PKC-ERK/JNK/p38 and PI3K signaling pathways in HepG2 cells. Acta Pharmacol. Sin. 2020, 41, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Bozaykut, P.; Sahin, A.; Karademir, B.; Ozer, N.K. Endoplasmic reticulum stress related molecular mechanisms in nonalcoholic steatohepatitis. Mech. Ageing Dev. 2016, 157, 17–29. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, C.; Wang, H.; Xu, J.; Duan, Z.H.; Zhang, Y.; Yu, T.; Wei, W.; Xu, D.X.; Xu, J.M. Altered integrity and decreased expression of hepatocyte tight junctions in rifampicin-induced cholestasis in mice. Toxicol. Appl. Pharmacol. 2009, 240, 26–36. [Google Scholar] [CrossRef]
- Fu, D.; Cardona, P.; Ho, H.; Watkins, P.B.; Brouwer, K.L.R. Novel Mechanisms of Valproate Hepatotoxicity: Impaired Mrp2 Trafficking and Hepatocyte Depolarization. Toxicol. Sci. 2019, 171, 431–442. [Google Scholar] [CrossRef]
- Laurenty, A.P.; Thomas, F.; Chatelut, E.; Bétrian, S.; Le Guellec, C.; Hennebelle, I.; Le Guellec, S.; Chevreau, C. Irreversible hepatotoxicity after administration of trabectedin to a pleiomorphic sarcoma patient with a rare ABCC2 polymorphism: A case report. Pharmacogenomics 2013, 14, 1389–1396. [Google Scholar] [CrossRef]
- Hafey, M.J.; Houle, R.; Tanis, K.Q.; Knemeyer, I.; Shang, J.; Chen, Q.; Baudy, A.; Monroe, J.; Sistare, F.D.; Evers, R. A Two-Tiered In Vitro Approach to De-Risk Drug Candidates for Potential Bile Salt Export Pump Inhibition Liabilities in Drug Discovery. Drug. Metab. Dispos. 2020, 48, 1147–1160. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.X.; Xu, X.F.; Zhang, Q.Z.; Li, C.; Deng, Y.; Jiang, P.; He, L.Y.; Peng, W.X. The inhibition of hepatic bile acids transporters Ntcp and Bsep is involved in the pathogenesis of isoniazid/rifampicin-induced hepatotoxicity. Toxicol. Mech. Methods 2015, 25, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Meier, Y.; Zodan, T.; Lang, C.; Zimmermann, R.; Kullak-Ublick, G.A.; Meier, P.J.; Stieger, B.; Pauli-Magnus, C. Increased susceptibility for intrahepatic cholestasis of pregnancy and contraceptive-induced cholestasis in carriers of the 1331T>C polymorphism in the bile salt export pump. World J. Gastroenterol. 2008, 14, 38–45. [Google Scholar] [CrossRef]
- Kagawa, T.; Hirose, S.; Arase, Y.; Oka, A.; Anzai, K.; Tsuruya, K.; Shiraishi, K.; Orii, R.; Ieda, S.; Nakazawa, T.; et al. No contribution of the ABCB11 p.444A polymorphism in Japanese patients with drug-induced cholestasis. Drug. Metab. Dispos. 2015, 43, 691–697. [Google Scholar] [CrossRef]
- Ulzurrun, E.; Stephens, C.; Crespo, E.; Ruiz-Cabello, F.; Ruiz-Nuñez, J.; Saenz-López, P.; Moreno-Herrera, I.; Robles-Díaz, M.; Hallal, H.; Moreno-Planas, J.M.; et al. Role of chemical structures and the 1331T>C bile salt export pump polymorphism in idiosyncratic drug-induced liver injury. Liver. Int. 2013, 33, 1378–1385. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Wang, J.; Tang, S.; Zhang, Y.; Lv, X.; Wu, S.; Yang, Z.; Xia, Y.; Chen, D.; Zhan, S. Role of polymorphic bile salt export pump (BSEP, ABCB11) transporters in anti-tuberculosis drug-induced liver injury in a Chinese cohort. Sci. Rep. 2016, 6, 27750. [Google Scholar] [CrossRef]
- Aleo, M.D.; Luo, Y.; Swiss, R.; Bonin, P.D.; Potter, D.M.; Will, Y. Human drug-induced liver in-jury severity is highly associated with dual inhibition of liver mitochondrial function and bile salt export pump. Hepatology 2014, 60, 1015–1022. [Google Scholar] [CrossRef]
- Stieger, B. Role of the bile salt export pump, BSEP, in acquired forms of cholestasis. Drug. Metab. Rev. 2010, 42, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Padda, M.S.; Sanchez, M.; Akhtar, A.J.; Boyer, J.L. Drug-induced cholestasis. Hepatology 2011, 53, 1377–1387. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver. Electronic address: Easloffice@easloffice.eu; Clinical Practice Guideline Panel: Chair; Panel members; EASL Governing Board representative: EASL Clinical Practice Guidelines: Drug-induced liver injury. J. Hepatol. 2019, 70, 1222–1261. [Google Scholar] [CrossRef] [PubMed]
- Fotoulaki, M.; Giza, S.; Jirsa, M.; Grammatikopoulos, T.; Miquel, R.; Hytiroglou, P.; Tsitouridis, I.; Knisely, A.S. Beyond an Obvious Cause of Cholestasis in a Toddler: Compound Heterozygosity for ABCB11 Mutations. Pediatrics 2019, 143, e20182146. [Google Scholar] [CrossRef]
- Vitale, G.; Pirillo, M.; Mantovani, V.; Marasco, E.; Aquilano, A.; Gamal, N.; Francalanci, P.; Conti, F.; Andreone, P. Bile salt export pump deficiency disease: Two novel, late onset, ABCB11 mutations identified by next generation sequencing. Ann. Hepatol. 2016, 15, 795–800. [Google Scholar] [CrossRef]
- Perrault, F.; Echelard, P.; Viens, D.; Borduas, M. Contraceptive vaginal ring-induced cholestasis in a patient with a history of intrahepatic cholestasis of pregnancy. Clin. Res. Hepatol. Gastroenterol. 2021, 45, 101475. [Google Scholar] [CrossRef]
- Aleo, M.D.; Shah, F.; He, K.; Bonin, P.D.; Rodrigues, A.D. Evaluating the Role of Multidrug Resistance Protein 3 (MDR3) Inhibition in Predicting Drug-Induced Liver Injury Using 125 Pharmaceuticals. Chem. Res. Toxicol. 2017, 30, 1219–1229, Erratum in Chem. Res. Toxicol. 2017, 30, 1365. [Google Scholar] [CrossRef]
- Mahdi, Z.M.; Synal-Hermanns, U.; Yoker, A.; Locher, K.P.; Stieger, B. Role of Multidrug Resistance Protein 3 in Antifungal-Induced Cholestasis. Mol. Pharmacol. 2016, 90, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Kolarić, T.O.; Ninčević, V.; Smolić, R.; Smolić, M.; Wu, G.Y. Mechanisms of Hepatic Cholestatic Drug Injury. J. Clin. Transl. Hepatol. 2019, 7, 86–92. [Google Scholar] [CrossRef]
- Dixon, P.H.; Williamson, C. The pathophysiology of intrahepatic cholestasis of pregnancy. Clin. Res. Hepatol. Gastroenterol. 2016, 40, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Azzaroli, F.; Turco, L.; Lisotti, A.; Calvanese, C.; Mazzella, G. The pharmacological management of intrahepatic cholestasis of pregnancy. Curr. Clin. Pharmacol. 2011, 6, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Falcão, D.; Pedroto, I.; Moreira, T. The wide phenotypic and genetic spectrum of ABCB4 gene deficiency: A case series. Dig. Liver. Dis. 2022, 54, 221–227. [Google Scholar] [CrossRef]
- Cardoso, M.F.; EBranco, J.C.; Anapaz, V.; Rodrigues, C.G.; Carvalho, R.; Horta, D.; Martins, A.; Reis, J. A Complex Case of Cholestasis in a Patient with ABCB4 and ABCB11 Mutations. GE Port. J. Gastroenterol. 2018, 25, 189–194. [Google Scholar] [CrossRef]
- Sinha, A.; Bhuva, M.; Grant, C.; Gimson, A.E.; Thompson, E.; Duckworth, A.; Davies, S.E.; Aithal, G.; Griffiths, W.J. ABCB4 Mutations in Adults Cause a Spectrum Cholestatic Disorder Histologically Distinct from Other Biliary Disease. Dig. Dis. Sci. 2022, 67, 5551–5561. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.J.; van Helvoort, A.; van Meer, G.; Szabo, K.; Welker, E.; Szakacs, G.; Varadi, A.; Sarkadi, B.; Borst, P. MDR3 P-glycoprotein, a phosphatidylcholine translocase, transports several cytotoxic drugs and directly interacts with drugs as judged by interference with nucleotide trapping. J. Biol. Chem. 2000, 275, 23530–23539. [Google Scholar] [CrossRef]
- Hooiveld, G.J.; van Montfoort, J.E.; Meijer, D.K.; Müller, M. Function and regulation of ATP-binding cassette transport proteins involved in hepatobiliary transport. Eur. J. Pharm. Sci. 2000, 12, 13–30, Corrected and republished in: Eur. J. Pharm. Sci. 2001, 12, 525–543. [Google Scholar] [CrossRef] [PubMed]
- Bonin, S.; Pascolo, L.; Crocé, L.S.; Stanta, G.; Tiribelli, C. Gene expression of ABC proteins in hepatocellular carcinoma, perineoplastic tissue, and liver diseases. Mol. Med. 2002, 8, 318–325. [Google Scholar] [CrossRef]
- Björnsson, E.S.; Hoofnagle, J.H. Categorization of drugs implicated in causing liver injury: Critical assessment based on published case reports. Hepatology 2016, 63, 590–603. [Google Scholar] [CrossRef] [PubMed]
- European Association for the Study of the Liver. Electronic address: Easloffice@easloffice.eu; European Association for the Study of the Liver: EASL Clinical Practice Guidelines: The diagnosis and management of patients with primary biliary cholangitis. J. Hepatol. 2017, 67, 145–172. [Google Scholar] [CrossRef]
- Kruk, B.; Milkiewicz, M.; Raszeja-Wyszomirska, J.; Milkiewicz, P.; Krawczyk, M. A common variant in the hepatobiliary phospholipid transporter ABCB4 modulates liver injury in PBC but not in PSC: Prospective analysis in 867 patients. Orphanet. J. Rare Dis. 2022, 17, 419. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of DILI | ||||
|---|---|---|---|---|
| Items | Hepatocellular | Score | Mixed/Cholestatic | Score |
| (1) Time to onset from the beginning of the drug/herb |
| +2 +1 |
| +2 +1 |
| Alternative: Time to onset from cessation of the drug/herb |
| +1 |
| +1 |
| (2) Course of liver exams after cessation of the drug/herb |
| +3 +2 0 0 −2 |
| +2 +1 0 |
| (3) Risk factors |
| +1 0 +1 +1 0 |
| +1 0 +1 +1 0 |
| (4) Concomitant drug(s)/herb(s) |
| 0 |
| 0 |
| 0 |
| 0 | |
| −1 |
| −1 | |
| −2 |
| −2 | |
| −3 |
| −3 | |
| (5) Search for alternative causes | Group I (7 causes)
Group II (5 causes)
EBV HSV VZV | Group I (7 causes)
Group II (5 causes)
EBV HSV VZV | ||
Evaluation of groups I and II
| +2 +1 0 −2 −3 | Evaluation of groups I and II
| +2 +1 0 −2 −3 | |
| (6) Previous hepatotoxicity of the drug/herb |
| +2 +1 0 |
| +2 +1 0 |
| (7) Response to unintentional reexposure |
| +3 +1 −2 0 |
| +3 +1 −2 0 |
| Genes, Protein, Cell Position | Function |
In Vitro
Inhibitors of ABC Transporters and In Vivo Liver Injury-Related Drugs |
Variants, Prediction, Transcript |
|---|---|---|---|
| ABCB1 MDR1 Canalicular membrane | It works both as a functional barrier and as an efflux transporter. In addition, it drives the excretion of multiple drugs, especially of quaternary cationic amines, converging them into bile | Cyclosporine, Erythromycin, Chlorpromazine, Anticancer drugs, HIV protease inhibitors, Amiodarone, Antituberculosis Drugs, Nefazodone, Telithromycin, Carvedilol, Clarithromycin, Itraconazole, Lapatinib, Verapamil | c.2677T>G p.S893A (LB) c.3435T>G p.I1145M (VUS) Transcript: NM_000927.4 |
| ABCC2 MRP2 Canalicular membrane | ATP-dependent efflux of numerous drugs, organic anions, such as glutathione, bilirubin, leukotrienes, and sulfated or glucuronidated BAs | Anticancer drugs (Trabectedin) Benzbromarone, Bosentan, Cyclosporine, Diclofenac, Efavirenz, Probenecid, Rifampicin, Valproate | c.-24C>T in 5’UTR # c.4146+154A>G # c.4488C>T p.H1496H (B) c.1249G>A p.V417I (B) c.3972C>T p.I1324 (B) c.4544G>A p.C1515Y (B) Transcript: NM_000392.5 |
| ABCB11 BSEP Canalicular membrane | It eliminates unconjugated and conjugated BAs from hepatocytes into the bile against a concentration gradient. This function is essential for keeping the potentially cytotoxic BAs at a low intracellular level in hepatocytes | Benzbromarone, Bosentan, Cyclosporine, Isoniazid, Rifampicin, Glitazones, Statins, Sulindac, Erythromycin, Glibenclamide, Nefazodone, Progesterone metabolites, Estradiol 17b-Glucuronide, Taxol, Cefprozil combined with homoeopathic preparation of cantharidin, Antibacterials with b-lactam ring | c.1331T>C p.V444A (B) c.2026G>T p.D676Y § (LB) c.278A>C p.Y93S (VUS) c.1568C>G p.A523G (VUS) c.1789G>C p.V597L (VUS) c.2563G>A p.G855R (LP) c.3449_3440delGTinsTA p.V1147* (LP) c.3148C>T p.R1050C (P) c.3382C>T p.R1128C (P) c.343 c.3904G>T p.E1302* (P) Transcript: NM_003742.4 |
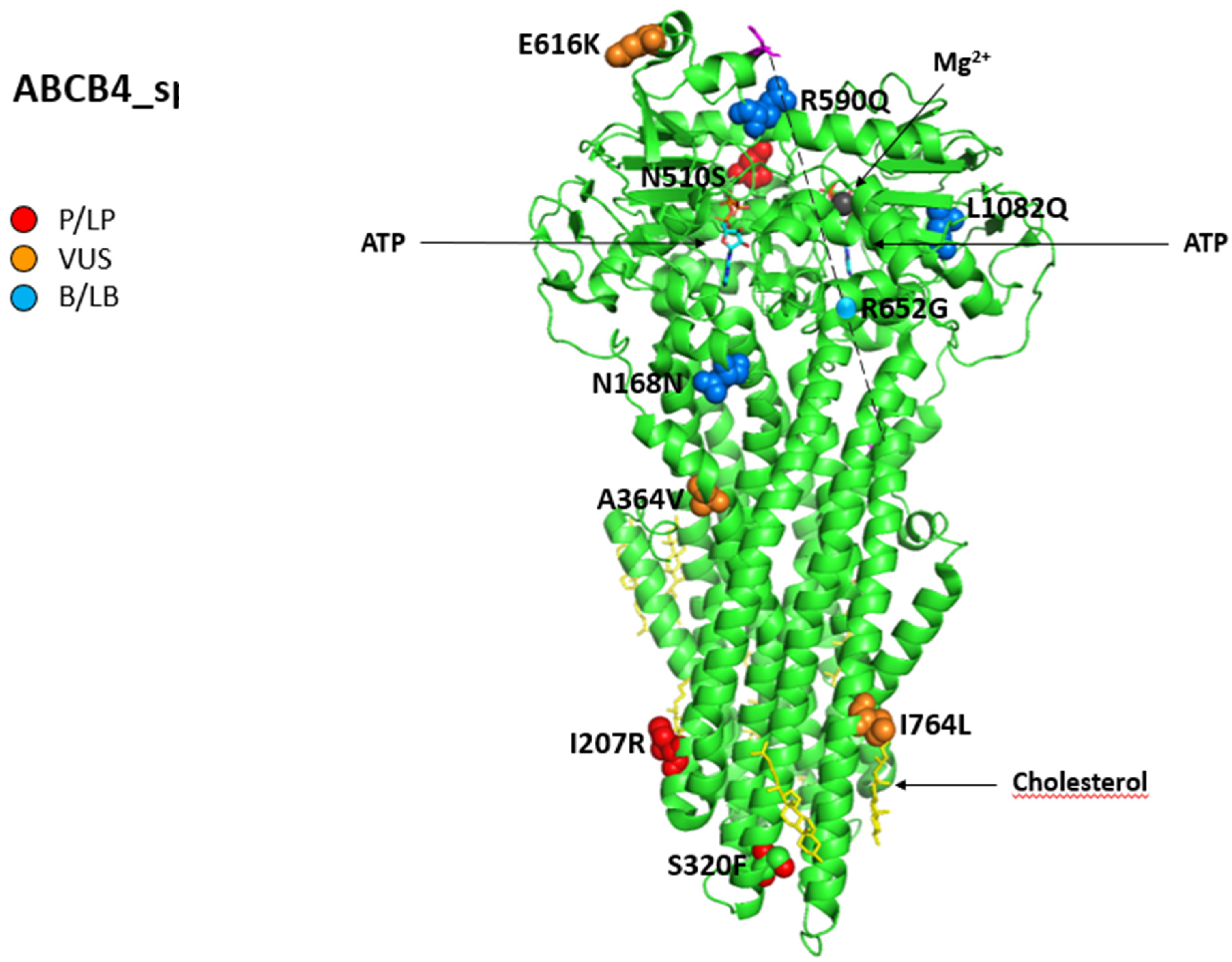
| ABCB4 MDR3 Canalicular membrane | It works as “floppase”, translocating PL of the PC family from the inner to the outer side of the lipid bilayer of the canalicular membrane of hepatocytes; it is responsible for PL secretion into the bile, regulating the development of mixed micelles | Itraconazole, Chlorpromazine, Imipramine, Haloperidol, Ketoconazole, Clotrimazole, Troglitazone, Sirolimus, Cyclosporine, Verapamil, Vinblastine, Atenolol, Losartan Risperidone, Proton pump inhibitors, Antibacterials with b-lactam ring, Oral contraceptives, Ciprofloxacin | c.504C>T p.N168N (B) c.1769G>A p.R590Q (B) c.1954A>G p.R652G (B) c.3245T>A p.L1082Q (LB) c.1091C>T p.A364V (VUS) c.1529A>G p.N510S (VUS) c.1846G>A p.E616K (VUS) c.2290A>C p.I764L (VUS) c.620T>G p.I207R (LP) c.2014A>T p.K672* (LP) c.959C>T p.S320F (P) c.2301dupT p.T768Yfs*26 § (P) Transcript: NM_000443.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vitale, G.; Mattiaccio, A.; Conti, A.; Berardi, S.; Vero, V.; Turco, L.; Seri, M.; Morelli, M.C. Molecular and Clinical Links between Drug-Induced Cholestasis and Familial Intrahepatic Cholestasis. Int. J. Mol. Sci. 2023, 24, 5823. https://doi.org/10.3390/ijms24065823
Vitale G, Mattiaccio A, Conti A, Berardi S, Vero V, Turco L, Seri M, Morelli MC. Molecular and Clinical Links between Drug-Induced Cholestasis and Familial Intrahepatic Cholestasis. International Journal of Molecular Sciences. 2023; 24(6):5823. https://doi.org/10.3390/ijms24065823
Chicago/Turabian StyleVitale, Giovanni, Alessandro Mattiaccio, Amalia Conti, Sonia Berardi, Vittoria Vero, Laura Turco, Marco Seri, and Maria Cristina Morelli. 2023. "Molecular and Clinical Links between Drug-Induced Cholestasis and Familial Intrahepatic Cholestasis" International Journal of Molecular Sciences 24, no. 6: 5823. https://doi.org/10.3390/ijms24065823
APA StyleVitale, G., Mattiaccio, A., Conti, A., Berardi, S., Vero, V., Turco, L., Seri, M., & Morelli, M. C. (2023). Molecular and Clinical Links between Drug-Induced Cholestasis and Familial Intrahepatic Cholestasis. International Journal of Molecular Sciences, 24(6), 5823. https://doi.org/10.3390/ijms24065823