
Protein Unfolding—Thermodynamic Perspectives and Unfolding Models

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
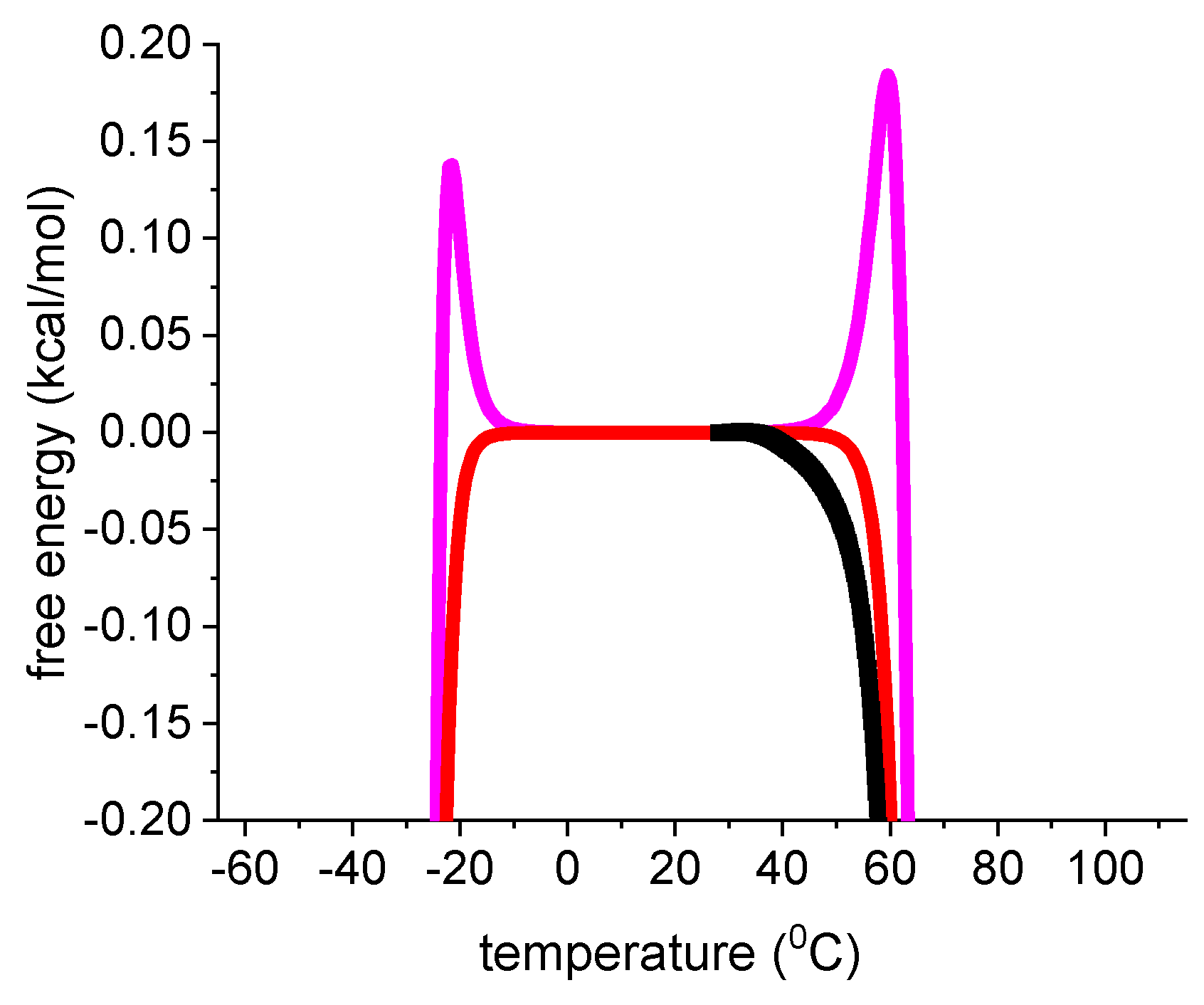{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
2.1. Differential Scanning Calorimetry (DSC). Model-Independent Thermodynamic Analysis of Protein Unfolding Experiments
2.2. Models for Protein Unfolding
2.2.1. Chemical Equilibrium Two-State Models
- (a)
- Van’t Hoff Enthalpy Model:
- (b)
- Free Energy Chemical Equilibrium Two-State Model (“Standard Model”):
- (c)
- ΘU(T)-Weighted Chemical Equilibrium Two-State Model [15]:
2.2.2. Statistical-Mechanical Models
- (a)
- Partition Function Z(T) and Thermodynamic Properties:
- (b)
- Statistical-Mechanical 2-State Unfolding Model [15]:
- (c)
- Multistate Cooperative Unfolding Model [13]:
3. DSC Experiments Compared to the Three New Protein Unfolding Models
3.1. Lysozyme Heat Unfolding
3.2. β-Lactoglobulin Cold and Heat Denaturation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zimm, B.H.; Bragg, J.K. Theory of the Phase Transition between Helix and Random Coil in Polypeptide Chains. J. Chem. Phys. 1959, 31, 526–535. [Google Scholar] [CrossRef]
- Davidson, N. Statistical Mechanics; Mac Graw-Hill: New York, NY, USA, 1962; p. 385. [Google Scholar]
- Schwarz, G.; Seelig, J. Kinetic Properties and Electric Field Effect of Helix-Coil Transition of Poly(Gamma-Benzyl L-Glutamate) Determined from Dielectric Relaxation Measurements. Biopolymers 1968, 6, 1263. [Google Scholar] [CrossRef] [PubMed]
- Ising, E. Report on the theory of ferromagnetism. Z. Fur Phys. 1925, 31, 253–258. [Google Scholar] [CrossRef]
- Brandts, J.F. Thermodynamics of Protein Denaturation. I. Denaturation of Chymotrypsinogen. J. Am. Chem. Soc. 1964, 86, 4291. [Google Scholar] [CrossRef]
- Brandts, J.F. Thermodynamics of Protein Denaturation. 2. Model of Reversible Denaturation + Interpretations Regarding Stability of Chymotrypsinogen. J. Am. Chem. Soc. 1964, 86, 4302. [Google Scholar] [CrossRef]
- Privalov, P.L.; Dragan, A.I. Microcalorimetry of biological macromolecules. Biophys. Chem. 2007, 126, 16–24. [Google Scholar] [CrossRef]
- Ibarra-Molero, B.; N.Naganathan, A.; Sanchez-Ruiz, J.M.; Munoz, V. Modern analysis of protein folding by differential scanning calorimetry. In Methods in enzymology; Elsevier: Amsterdam, The Netherlands, 2016; pp. 281–316. [Google Scholar]
- Jackson, W.M.; Brandts, J.F. Thermodynamics of Protein Denaturation—A Calorimetric Study of Reversible Denaturation of Chymotrypsinogen and Conclusions Regarding Accuracy of 2-State Approximation. Biochemistry 1970, 9, 2294. [Google Scholar] [CrossRef]
- Privalov, P.L. Thermodynamics of protein folding. J. Chem. Thermodyn. 1997, 29, 447–474. [Google Scholar] [CrossRef]
- Ibarra-Molero, B.; Sanchez-Ruiz, J.M. Statistical Differential Scanning Calorimetry: Probing Protein Folding-Unfolding Ensembles. Protein Fold. Misfolding Aggreg. Class. Nov. Approaches 2008, 13, 85–105. [Google Scholar]
- Zhou, Y.; Hall, C.K.; Karplus, M. The calorimetric criterion for a two-state process revisited. Protein Sci. Publ. Protein Soc. 1999, 8, 1064–1074. [Google Scholar] [CrossRef]
- Seelig, J.; Seelig, A. Molecular Understanding of Calorimetric Protein Unfolding Experiments. Biophys. Rep. 2022, 2, 1–11. [Google Scholar] [CrossRef]
- Seelig, J. Free energy in thermal and chemical protein unfolding. In Gibbs energy and Helmholtz energy:Liquids, Solutions and Vapours; Letcher, E.W., Ed.; Royal Society of Chemistry: London, UK, 2022; pp. 363–378. [Google Scholar]
- Seelig, J.; Seelig, A. Protein stability—Analysis of heat and cold denaturation without and with unfolding models. J. Phys. Chem. B 2023, in press. [Google Scholar]
- Gomez, J.; Hilser, V.J.; Xie, D.; Freire, E. The heat capacity of proteins. Proteins 1995, 22, 404–412. [Google Scholar] [CrossRef]
- Privalov, P.L.; Khechinashvili, N.N. Thermodynamic Approach to Problem of Stabilization of Globular Protein Structure—Calorimetric Study. J. Mol. Biol. 1974, 86, 665–684. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.K.; Pace, C.N.; Scholtz, J.M. Denaturant m values and heat capacity changes: Relation to changes in accessible surface areas of protein unfolding. Protein Sci. Publ. Protein Soc. 1995, 4, 2138–2148. [Google Scholar] [CrossRef] [PubMed]
- Durowoju, I.B.; Bhandal, K.S.; Hu, J.; Carpick, B.; Kirkitadze, M. Differential Scanning Calorimetry—A Method for Assessing the Thermal Stability and Conformation of Protein Antigen. Jove-J. Vis. Exp. 2017, 121, e55262. [Google Scholar]
- Seelig, J.; Schönfeld, H.-J. Thermal protein unfolding by differential scanning calorimetry and circular dichroism spectroscopy Two-state model versus sequential unfolding. Q. Rev. Biophys. 2016, 49, e9. [Google Scholar] [CrossRef]
- Li-Blatter, X.; Seelig, J. Thermal and Chemical Unfolding of Lysozyme. Multistate Zimm-Bragg Theory Versus Two-State Model. J. Phys. Chem. B 2019, 123, 10181–10191. [Google Scholar] [CrossRef] [PubMed]
- Privalov, P.L.; Gill, S.J. Stability of Protein-Structure and Hydrophobic Interaction. Adv. Protein Chem. 1988, 39, 191–234. [Google Scholar]
- Privalov, G.; Kavina, V.; Freire, E.; Privalov, P.L. Precise Scanning Calorimeter for Studying Thermal-Properties of Biological Macromolecules in Dilute-Solution. Anal. Biochem. 1995, 232, 79–85. [Google Scholar] [CrossRef]
- Privalov, P.L. Stability of proteins: Small globular proteins. Adv. Protein Chem. 1979, 33, 167–241. [Google Scholar]
- Privalov, P.L.; Griko, Y.V.; Venyaminov, S.Y.; Kutyshenko, V.P. Cold Denaturation of Myoglobin. J. Mol. Biol. 1986, 190, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Privalov, P.L. Cold Denaturation of Proteins. Crit. Rev. Biochem. Mol. Biol. 1990, 25, 281–305. [Google Scholar] [CrossRef] [PubMed]
- Becktel, W.J.; Schellman, J.A. Protein stability curves. Biopolymers 1987, 26, 1859–1877. [Google Scholar] [CrossRef] [PubMed]
- Baumann, R.P. Evaluation of Thermodynamic Properties. In Modern Thermodynamics with Statistical Mechanics; McConnin, R.A., Ed.; Macmillan Publishing Company: New York, NY, USA, 1992; p. 341. [Google Scholar]
- Eisenberg, D.; Crothers, D. Calculation of the energy using the sysem partition function. In Physical Chemistry with Applications to the Life Sciences; Rhame, B., Moore, M., Eds.; The Benjamin/Cummings Publishing Company, Inc.: Menlo Park, CA, USA, 1979; p. 675. [Google Scholar]
- Zimm, B.H.; Doty, P.; Iso, K. Determination of the parameters for helix formation in poly-gamma-benzyl-L-glutamate. Proc. Natl. Acad. Sci. USA 1959, 45, 1601–1607. [Google Scholar] [CrossRef]
- Zimm, B.H.; Rice, S.A. The Helix-Coil Transition in Charged Macromolecules. Mol. Phys. 1960, 3, 391–407. [Google Scholar] [CrossRef]
- Zehender, F.; Ziegler, A.; Schonfeld, H.J.; Seelig, J. Thermodynamics of Protein Self-Association and Unfolding. The Case of Apolipoprotein A-I. Biochem. -US 2012, 51, 1269–1280. [Google Scholar] [CrossRef]
- Schulthess, T.; Schonfeld, H.J.; Seelig, J. Thermal Unfolding of Apolipoprotein A-1. Evaluation of Methods and Models. Biochemistry 2015, 54, 3063–3075. [Google Scholar] [CrossRef]
- Seelig, J. Cooperative protein unfolding. A statistical-mechanical model for the action of denaturants. Biophys. Chem. 2018, 233, 19–25. [Google Scholar] [CrossRef]
- Eckhardt, D.; Li-Blatter, X.; Schonfeld, H.J.; Heerklotz, H.; Seelig, J. Cooperative unfolding of apolipoprotein A-1 induced by chemical denaturation. Biophys. Chem. 2018, 240, 42–49. [Google Scholar] [CrossRef]
- Garidel, P.; Eiperle, A.; Blech, M.; Seelig, J. Thermal and Chemical Unfolding of a Monoclonal IgG1 Antibody: Application of the Multistate Zimm-Bragg Theory. Biophys. J. 2020, 118, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Privalov, P.L.; Makhatadze, G.I. Heat capacity of proteins. II. Partial molar heat capacity of the unfolded polypeptide chain of proteins: Protein unfolding effects. J. Mol. Biol. 1990, 213, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Rosgen, J.; Hinz, H.J. Response functions of proteins. Biophys. Chem. 2000, 83, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Griko, Y.V.; Privalov, P.L. Calorimetric study of the heat and cold denaturation of beta-lactoglobulin. Biochemistry 1992, 31, 8810–8815. [Google Scholar] [CrossRef] [PubMed]
- Meier, M.; Seelig, J. Thermodynamics of the coil <==> beta-sheet transition in a membrane environment. J. Mol. Biol. 2007, 369, 277–289. [Google Scholar] [PubMed]
- Hedoux, A.; Krenzlin, S.; Paccou, L.; Guinet, Y.; Flament, M.P.; Siepmann, J. Influence of urea and guanidine hydrochloride on lysozyme stability and thermal denaturation; a correlation between activity, protein dynamics and conformational changes. Phys. Chem. Chem. Phys. PCCP 2010, 12, 13189–13196. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seelig, J.; Seelig, A. Protein Unfolding—Thermodynamic Perspectives and Unfolding Models. Int. J. Mol. Sci. 2023, 24, 5457. https://doi.org/10.3390/ijms24065457
Seelig J, Seelig A. Protein Unfolding—Thermodynamic Perspectives and Unfolding Models. International Journal of Molecular Sciences. 2023; 24(6):5457. https://doi.org/10.3390/ijms24065457
Chicago/Turabian StyleSeelig, Joachim, and Anna Seelig. 2023. "Protein Unfolding—Thermodynamic Perspectives and Unfolding Models" International Journal of Molecular Sciences 24, no. 6: 5457. https://doi.org/10.3390/ijms24065457
APA StyleSeelig, J., & Seelig, A. (2023). Protein Unfolding—Thermodynamic Perspectives and Unfolding Models. International Journal of Molecular Sciences, 24(6), 5457. https://doi.org/10.3390/ijms24065457