
Temporins: Multifunctional Peptides from Frog Skin

Abstract
1. Introduction
2. Temporins from Rana
2.1. Temporin 1CE Family
2.1.1. Temporin 1 CEa
2.1.2. Temporins 1-CEb, 1-CEc, 1-CEh

{kind=link}
{kind=link}
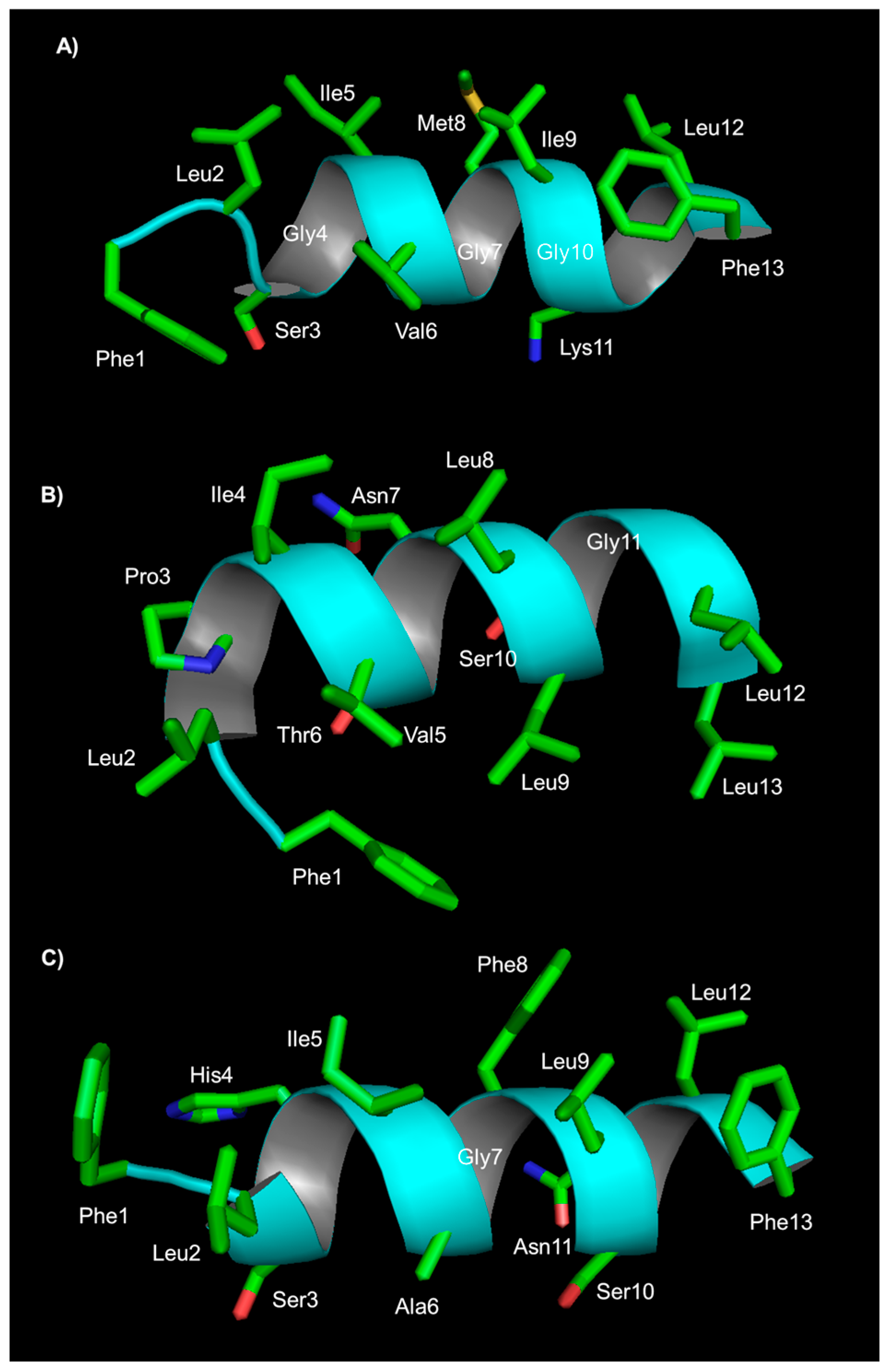{kind=link}
| Peptide | Antimicrobial Activity as MIC (µM) | |||
|---|---|---|---|---|
| S. aureus 22401 | Ref | E. coli 44102 | Ref | |
| Temporin 1CEa | 14.4 | [18] | >100 | [19] |
| Temporin 1CEb | 41 | [18] | >100 | [18] |
| Temporin 1CEc | >100 | [18] | >100 | [18] |
| Temporin 1CEe | 6.250 | [19] | 50.0 | [19] |
| Temporin 1CEh | 8 | [24] | 128 | [24] |
2.1.3. Temporin 1 CEa Analogues
2.1.4. Temporin 1CEb Analogues
2.1.5. Temporin 1CEc Analogues
2.2. Temporin A
2.2.1. Temporin A Biological Activity
2.2.2. Temporin A Structure and Mechanism of Action
2.2.3. Temporin A Analogues
2.3. Temporin B
2.3.1. Temporin B Biological Activity
2.3.2. Temporin B Structure and Mechanism of Action
2.3.3. Temporin B Analogues
2.4. Temporin G
2.5. Temporin L
2.5.1. Temporin L Biological Activity
2.5.2. Temporin L Structure and Mechanisms of Action
2.5.3. Temporin L Analogues
3. Temporins from Hylarana
Temporin GHa Analogues
4. Temporins from Lithobates
5. Temporins from Odorrana
6. Temporins from Pelophylax
6.1. Temporin SH Family
6.1.1. Temporin SHa Biological Activity
6.1.2. Temporin SHa Structure and Mechanism of Action
6.1.3. Temporin SHf Biological Activity
6.1.4. Temporin SHa Structure and Mechanism of Action
6.1.5. Temporin SH Analogues
7. Biomedical/Biotechnological Applications of Temporins
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Konig, E.; Bininda-Emonds, O.R.; Shaw, C. The diversity and evolution of anuran skin peptides. Peptides 2015, 63, 96–117. [Google Scholar] [CrossRef] [PubMed]
- Erspamer, V.; Ottolenghi, A. Preliminary researches on the mechanism of the antidiuretic action of enteramine. Experientia 1951, 7, 191–192. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Iwamuro, S.; King, J.D. Dermal cytolytic peptides and the system of innate immunity in anurans. Ann. N. Y. Acad. Sci. 2009, 1163, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M. Structural diversity and species distribution of host-defense peptides in frog skin secretions. Cell. Mol. Life Sci. 2011, 68, 2303–2315. [Google Scholar] [CrossRef]
- Patocka, J.; Nepovimova, E.; Klimova, B.; Wu, Q.; Kuca, K. Antimicrobial Peptides: Amphibian Host Defense Peptides. Curr. Med. Chem. 2019, 26, 5924–5946. [Google Scholar] [CrossRef] [PubMed]
- Ladram, A.; Nicolas, P. Antimicrobial peptides from frog skin: Biodiversity and therapeutic promises. Front. Biosci. (Landmark Ed.) 2016, 21, 1341–1371. [Google Scholar] [CrossRef]
- Vanhoye, D.; Bruston, F.; Nicolas, P.; Amiche, M. Antimicrobial peptides from hylid and ranin frogs originated from a 150-million-year-old ancestral precursor with a conserved signal peptide but a hypermutable antimicrobial domain. Eur. J. BioChem. 2003, 270, 2068–2081. [Google Scholar] [CrossRef]
- Samgina, T.Y.; Vasileva, I.D.; Trebse, P.; Torkar, G.; Surin, A.K.; Meng, Z.; Zubarev, R.A.; Lebedev, A.T. Mass Spectrometry Differentiation between Rana arvalis Populations Based on Their Skin Peptidome Composition. J. Am. Soc. Mass Spectrom. 2022, 33, 1480–1491. [Google Scholar] [CrossRef] [PubMed]
- Ohnuma, A.; Conlon, J.M.; Kawasaki, H.; Iwamuro, S. Developmental and triiodothyronine-induced expression of genes encoding preprotemporins in the skin of Tago’s brown frog Rana tagoi. Gen. Comp. Endocrinol. 2006, 146, 242–250. [Google Scholar] [CrossRef]
- Matutte, B.; Storey, K.B.; Knoop, F.C.; Conlon, J.M. Induction of synthesis of an antimicrobial peptide in the skin of the freeze-tolerant frog, Rana sylvatica, in response to environmental stimuli. FEBS Lett. 2000, 483, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Wang, G. Bioinformatic Analysis of 1000 Amphibian Antimicrobial Peptides Uncovers Multiple Length-Dependent Correlations for Peptide Design and Prediction. Antibiotics 2020, 9, 491. [Google Scholar] [CrossRef] [PubMed]
- Cortazar-Chinarro, M.; Meyer-Lucht, Y.; Van der Valk, T.; Richter-Boix, A.; Laurila, A.; Hoglund, J. Antimicrobial peptide and sequence variation along a latitudinal gradient in two anurans. BMC Genet. 2020, 21, 38. [Google Scholar] [CrossRef]
- Samgina, T.Y.; Vasileva, I.D.; Kovalev, S.V.; Trebse, P.; Torkar, G.; Surin, A.K.; Zubarev, R.A.; Lebedev, A.T. Differentiation of Central Slovenian and Moscow populations of Rana temporaria frogs using peptide biomarkers of temporins family. Anal. Bioanal. Chem. 2021, 413, 5333–5347. [Google Scholar] [CrossRef] [PubMed]
- Simmaco, M.; Mignogna, G.; Barra, D.; Bossa, F. Antimicrobial peptides from skin secretions of Rana esculenta. Molecular cloning of cDNAs encoding esculentin and brevinins and isolation of new active peptides. J. Biol. Chem. 1994, 269, 11956–11961. [Google Scholar] [CrossRef]
- Romero, S.M.; Cardillo, A.B.; Martinez Ceron, M.C.; Camperi, S.A.; Giudicessi, S.L. Temporins: An Approach of Potential Pharmaceutic Candidates. Surg. Infect. (Larchmt) 2020, 21, 309–322. [Google Scholar] [CrossRef]
- Conlon, J.M. Reflections on a systematic nomenclature for antimicrobial peptides from the skins of frogs of the family Ranidae. Peptides 2008, 29, 1815–1819. [Google Scholar] [CrossRef] [PubMed]
- Pirtskhalava, M.; Amstrong, A.A.; Grigolava, M.; Chubinidze, M.; Alimbarashvili, E.; Vishnepolsky, B.; Gabrielian, A.; Rosenthal, A.; Hurt, D.E.; Tartakovsky, M. DBAASP v3: Database of antimicrobial/cytotoxic activity and structure of peptides as a resource for development of new therapeutics. Nucleic Acids Res. 2021, 49, D288–D297. [Google Scholar] [CrossRef]
- Shang, D.; Yu, F.; Li, J.; Zheng, J.; Zhang, L.; Li, Y. Molecular cloning of cDNAs encoding antimicrobial peptide precursors from the skin of the Chinese brown frog, Rana chensinensis. Zool. Sci. 2009, 26, 220–226. [Google Scholar] [CrossRef]
- Zhao, J.; Sun, Y.; Li, Z.; Su, Q. Molecular cloning of novel antimicrobial peptide genes from the skin of the Chinese brown frog, Rana chensinensis. Zool. Sci. 2011, 28, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, Y.; Kang, Y.; Shang, D. Antimicrobial peptide temporin-1CEa isolated from frog skin secretions inhibits the proinflammatory response in lipopolysaccharide-stimulated RAW264.7 murine macrophages through the MyD88-dependent signaling pathway. Mol. Immunol. 2021, 132, 227–235. [Google Scholar] [CrossRef]
- Wang, C.; Li, H.B.; Li, S.; Tian, L.L.; Shang, D.J. Antitumor effects and cell selectivity of temporin-1CEa, an antimicrobial peptide from the skin secretions of the Chinese brown frog (Rana chensinensis). Biochimie 2012, 94, 434–441. [Google Scholar] [CrossRef]
- Wang, C.; Chen, Y.W.; Zhang, L.; Gong, X.G.; Zhou, Y.; Shang, D.J. Melanoma cell surface-expressed phosphatidylserine as a therapeutic target for cationic anticancer peptide, temporin-1CEa. J. Drug Target. 2016, 24, 548–556. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, Y.; Li, S.; Li, H.; Tian, L.; Wang, H.; Shang, D. Anticancer mechanisms of temporin-1CEa, an amphipathic alpha-helical antimicrobial peptide, in Bcap-37 human breast cancer cells. Life Sci. 2013, 92, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Zhou, X.; Xi, X.; Zai, Y.; Zhou, M.; Chen, X.; Ma, C.; Chen, T.; Wang, L.; Kwok, H.F. In Vitro & In Vivo Studies on Identifying and Designing Temporin-1CEh from the Skin Secretion of Rana chensinensis as the Optimised Antibacterial Prototype Drug. Pharmaceutics 2022, 14, 604. [Google Scholar] [PubMed]
- Yang, Q.Z.; Wang, C.; Lang, L.; Zhou, Y.; Wang, H.; Shang, D.J. Design of potent, non-toxic anticancer peptides based on the structure of the antimicrobial peptide, temporin-1CEa. Arch. Pharm. Res. 2013, 36, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Shang, D.; Li, X.; Sun, Y.; Wang, C.; Sun, L.; Wei, S.; Gou, M. Design of potent, non-toxic antimicrobial agents based upon the structure of the frog skin peptide, temporin-1CEb from Chinese brown frog, Rana chensinensis. Chem. Biol. Drug Des. 2012, 79, 653–662. [Google Scholar] [CrossRef]
- Shang, D.; Liang, H.; Wei, S.; Yan, X.; Yang, Q.; Sun, Y. Effects of antimicrobial peptide L-K6, a temporin-1CEb analog on oral pathogen growth, Streptococcus mutans biofilm formation, and anti-inflammatory activity. Appl. Microbiol. Biotechnol. 2014, 98, 8685–8695. [Google Scholar] [CrossRef]
- Dong, W.; Zhu, X.; Zhou, X.; Yang, Y.; Yan, X.; Sun, L.; Shang, D. Potential role of a series of lysine-/leucine-rich antimicrobial peptide in inhibiting lipopolysaccharide-induced inflammation. Biochem. J. 2018, 475, 3687–3706. [Google Scholar] [CrossRef]
- Bi, X.; Wang, C.; Ma, L.; Sun, Y.; Shang, D. Investigation of the role of tryptophan residues in cationic antimicrobial peptides to determine the mechanism of antimicrobial action. J. Appl. Microbiol. 2013, 115, 663–672. [Google Scholar] [CrossRef]
- Bi, X.; Wang, C.; Dong, W.; Zhu, W.; Shang, D. Antimicrobial properties and interaction of two Trp-substituted cationic antimicrobial peptides with a lipid bilayer. J. Antibiot. (Tokyo) 2014, 67, 361–368. [Google Scholar] [CrossRef]
- Wang, C.; Dong, S.; Zhang, L.; Zhao, Y.; Huang, L.; Gong, X.; Wang, H.; Shang, D. Cell surface binding, uptaking and anticancer activity of L-K6, a lysine/leucine-rich peptide, on human breast cancer MCF-7 cells. Sci. Rep. 2017, 7, 8293. [Google Scholar] [CrossRef]
- Wang, C.; Huang, L.L.; Li, R.J.; Wang, Y.; Wu, X.X.; Shang, D.J. Synergistic Therapy of Doxorubicin with Cationic Anticancer Peptide L-K6 Reverses Multidrug Resistance in MCF-7/ADR Cancer Cells In Vitro via P-glycoprotein Inhibition. Int. J. Pept. Res. Ther. 2021, 27, 2291–2301. [Google Scholar] [CrossRef]
- Kosikowska, P.; Pikula, M.; Langa, P.; Trzonkowski, P.; Obuchowski, M.; Lesner, A. Synthesis and Evaluation of Biological Activity of Antimicrobial--Pro-Proliferative Peptide Conjugates. PLoS ONE 2015, 10, e0140377. [Google Scholar] [CrossRef]
- Golda, A.; Kosikowska-Adamus, P.; Babyak, O.; Lech, M.; Wysocka, M.; Lesner, A.; Potempa, J.; Koziel, J. Conjugate of Enkephalin and Temporin Peptides as a Novel Therapeutic Agent for Sepsis. Bioconjug. Chem. 2018, 29, 4127–4139. [Google Scholar] [CrossRef] [PubMed]
- Golda, A.; Kosikowska-Adamus, P.; Kret, A.; Babyak, O.; Wojcik, K.; Dobosz, E.; Potempa, J.; Lesner, A.; Koziel, J. The Bactericidal Activity of Temporin Analogues Against Methicillin Resistant. Staphylococcus aureus. Int. J. Mol. Sci. 2019, 20, 4761. [Google Scholar] [CrossRef]
- Ji, F.; Zhao, Y.; Jiang, F.; Shang, D. Membrane mechanism of temporin-1CEc, an antimicrobial peptide isolated from the skin secretions of Rana chensinensis, and its systemic analogs. Bioorg. Chem. 2022, 119, 105544. [Google Scholar] [CrossRef]
- Simmaco, M.; Mignogna, G.; Canofeni, S.; Miele, R.; Mangoni, M.L.; Barra, D. Temporins, antimicrobial peptides from the European red frog Rana temporaria. Eur. J. Biochem. 1996, 242, 788–792. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.L. Temporins, anti-infective peptides with expanding properties. Cell. Mol. Life Sci. 2006, 63, 1060–1069. [Google Scholar] [CrossRef] [PubMed]
- Wade, D.; Silberring, J.; Soliymani, R.; Heikkinen, S.; Kilpelainen, I.; Lankinen, H.; Kuusela, P. Antibacterial activities of temporin A analogs. Febs. Lett. 2000, 479, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Di Grazia, A.; Luca, V.; Segev-Zarko, L.A.; Shai, Y.; Mangoni, M.L. Temporins A and B stimulate migration of HaCaT keratinocytes and kill intracellular Staphylococcus aureus. Antimicrob. Agents Chemother. 2014, 58, 2520–2527. [Google Scholar] [CrossRef]
- Simonetti, O.; Cirioni, O.; Goteri, G.; Ghiselli, R.; Kamysz, W.; Kamysz, E.; Silvestri, C.; Orlando, F.; Barucca, C.; Scalise, A.; et al. Temporin A is effective in MRSA-infected wounds through bactericidal activity and acceleration of wound repair in a murine model. Peptides 2008, 29, 520–528. [Google Scholar] [CrossRef]
- Ghiselli, R.; Giacometti, A.; Cirioni, O.; Mocchegiani, F.; Orlando, F.; Kamysz, W.; Del Prete, M.S.; Lukasiak, J.; Scalise, G.; Saba, V. Temporin A as a prophylactic agent against methicillin sodium-susceptible and methicillin sodium-resistant Staphylococcus epidermidis vascular graft infection. J. Vasc. Surg. 2002, 36, 1027–1030. [Google Scholar] [CrossRef]
- Ciandrini, E.; Morroni, G.; Cirioni, O.; Kamysz, W.; Kamysz, E.; Brescini, L.; Baffone, W.; Campana, R. Synergistic combinations of antimicrobial peptides against biofilms of methicillin-resistant Staphylococcus aureus (MRSA) on polystyrene and medical devices. J. Glob. Antimicrob. Resist. 2020, 21, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Giacometti, A.; Cirioni, O.; Kamysz, W.; Silvestri, C.; Licci, A.; Riva, A.; Lukasiak, J.; Scalise, G. In vitro activity of amphibian peptides alone and in combination with antimicrobial agents against multidrug-resistant pathogens isolated from surgical wound infection. Peptides 2005, 26, 2111–2116. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Ghiselli, R.; Orlando, F.; D’Amato, G.; Kamysz, W.; Mocchegiani, F.; Sisti, V.; Silvestri, C.; Lukasiak, J.; et al. Temporin A soaking in combination with intraperitoneal linezolid prevents vascular graft infection in a subcutaneous rat pouch model of infection with Staphylococcus epidermidis with intermediate resistance to glycopeptides. Antimicrob. Agents Chemother. 2004, 48, 3162–3164. [Google Scholar] [CrossRef]
- Cirioni, O.; Giacometti, A.; Ghiselli, R.; Kamysz, W.; Orlando, F.; Mocchegiani, F.; Silvestri, C.; Licci, A.; Lukasiak, J.; Saba, V.; et al. Temporin A alone and in combination with imipenem reduces lethality in a mouse model of staphylococcal sepsis. J. Infect. Dis. 2005, 192, 1613–1620. [Google Scholar] [CrossRef] [PubMed]
- Paduszynska, M.A.; Greber, K.E.; Paduszynski, W.; Sawicki, W.; Kamysz, W. Activity of Temporin A and Short Lipopeptides Combined with Gentamicin against Biofilm Formed by Staphylococcus aureus and Pseudomonas aeruginosa. Antibiotics 2020, 9, 566. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Shai, Y. Temporins and their synergism against Gram-negative bacteria and in lipopolysaccharide detoxification. Biochim. Biophys. Acta 2009, 1788, 1610–1619. [Google Scholar] [CrossRef]
- Capparelli, R.; Romanelli, A.; Iannaccone, M.; Nocerino, N.; Ripa, R.; Pensato, S.; Pedone, C.; Iannelli, D. Synergistic antibacterial and anti-inflammatory activity of temporin A and modified temporin B in vivo. PLoS ONE 2009, 4, e7191. [Google Scholar] [CrossRef] [PubMed]
- Chinchar, V.G.; Bryan, L.; Silphadaung, U.; Noga, E.; Wade, D.; Rollins-Smith, L. Inactivation of viruses infecting ectothermic animals by amphibian and piscine antimicrobial peptides. Virology 2004, 323, 268–275. [Google Scholar] [CrossRef]
- Marimuthu, S.K.; Nagarajan, K.; Perumal, S.K.; Palanisamy, S.; Subbiah, L. Insilico Alpha-Helical Structural Recognition of Temporin Antimicrobial Peptides and Its Interactions with Middle East Respiratory Syndrome-Coronavirus. Int. J. Pept. Res. Ther. 2020, 26, 1473–1483. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.L.; Saugar, J.M.; Dellisanti, M.; Barra, D.; Simmaco, M.; Rivas, L. Temporins, small antimicrobial peptides with leishmanicidal activity. J. Biol. Chem. 2005, 280, 984–990. [Google Scholar] [CrossRef]
- Chadbourne, F.L.; Raleigh, C.; Ali, H.Z.; Denny, P.W.; Cobb, S.L. Studies on the antileishmanial properties of the antimicrobial peptides temporin A, B and 1Sa. J. Pept. Sci. 2011, 17, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wade, D.; Kurosaka, K.; Wang, Z.Y.; Oppenheim, J.J.; Yang, D. Temporin A and related frog antimicrobial peptides use formyl peptide receptor-like 1 as a receptor to chemoattract phagocytes. J. Immunol. 2004, 173, 2652–2659. [Google Scholar] [CrossRef]
- Swithenbank, L.; Cox, P.; Harris, L.G.; Dudley, E.; Sinclair, K.; Lewis, P.; Cappiello, F.; Morgan, C. Temporin A and Bombinin H2 Antimicrobial Peptides Exhibit Selective Cytotoxicity to Lung Cancer Cells. Scientificn (Cairo) 2020, 2020, 3526286. [Google Scholar] [CrossRef]
- Musale, V.; Casciaro, B.; Mangoni, M.L.; Abdel-Wahab, Y.H.A.; Flatt, P.R.; Conlon, J.M. Assessment of the potential of temporin peptides from the frog Rana temporaria (Ranidae) as anti-diabetic agents. J. Pept. Sci. 2018, 24, e3065. [Google Scholar] [CrossRef]
- Xu, L.F.; Li, Y.; Jin, R.; Jiang, D.P.; Jiang, D.C. High spatial resolution observation of Temporin A at cell membranes using scanning ion conductive microscopy. Electrochem. Commun. 2022, 134, 107181. [Google Scholar] [CrossRef]
- Carotenuto, A.; Malfi, S.; Saviello, M.R.; Campiglia, P.; Gomez-Monterrey, I.; Mangoni, M.L.; Gaddi, L.M.; Novellino, E.; Grieco, P. A different molecular mechanism underlying antimicrobial and hemolytic actions of temporins A and L. J. Med. Chem. 2008, 51, 2354–2362. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, R.; Joshi, M.; Mohanram, H.; Bhunia, A.; Mangoni, M.L.; Bhattacharjya, S. NMR structure of temporin-1 ta in lipopolysaccharide micelles: Mechanistic insight into inactivation by outer membrane. PLoS ONE 2013, 8, e72718. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, Y.; Barra, D.; Simmaco, M.; Shai, Y.; Mangoni, M.L. A synergism between temporins toward Gram-negative bacteria overcomes resistance imposed by the lipopolysaccharide protective layer. J. Biol. Chem. 2006, 281, 28565–28574. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Rinaldi, A.C.; Di Giulio, A.; Mignogna, G.; Bozzi, A.; Barra, D.; Simmaco, M. Structure-function relationships of temporins, small antimicrobial peptides from amphibian skin. Eur. J. Biochem. 2000, 267, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Uccelletti, D.; Zanni, E.; Marcellini, L.; Palleschi, C.; Barra, D.; Mangoni, M.L. Anti-Pseudomonas activity of frog skin antimicrobial peptides in a Caenorhabditis elegans infection model: A plausible mode of action in vitro and in vivo. Antimicrob. Agents Chemother. 2010, 54, 3853–3860. [Google Scholar] [CrossRef] [PubMed]
- Avitabile, C.; Netti, F.; Orefice, G.; Palmieri, M.; Nocerino, N.; Malgieri, G.; D’Andrea, L.D.; Capparelli, R.; Fattorusso, R.; Romanelli, A. Design, structural and functional characterization of a Temporin-1b analog active against Gram-negative bacteria. Biochim. Biophys. Acta 2013, 1830, 3767–3775. [Google Scholar] [CrossRef]
- Marcocci, M.E.; Amatore, D.; Villa, S.; Casciaro, B.; Aimola, P.; Franci, G.; Grieco, P.; Galdiero, M.; Palamara, A.T.; Mangoni, M.L.; et al. The Amphibian Antimicrobial Peptide Temporin B Inhibits In Vitro Herpes Simplex Virus 1 Infection. Antimicrob. Agents Chemother. 2018, 62, e02367-17. [Google Scholar] [CrossRef] [PubMed]
- Sood, R.; Domanov, Y.; Kinnunen, P.K. Fluorescent temporin B derivative and its binding to liposomes. J. Fluoresc. 2007, 17, 223–234. [Google Scholar] [CrossRef]
- Malgieri, G.; Avitabile, C.; Palmieri, M.; D’Andrea, L.D.; Isernia, C.; Romanelli, A.; Fattorusso, R. Structural basis of a temporin 1b analogue antimicrobial activity against Gram negative bacteria determined by CD and NMR techniques in cellular environment. ACS Chem. Biol. 2015, 10, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Bhunia, A.; Saravanan, R.; Mohanram, H.; Mangoni, M.L.; Bhattacharjya, S. NMR structures and interactions of temporin-1Tl and temporin-1Tb with lipopolysaccharide micelles: Mechanistic insights into outer membrane permeabilization and synergistic activity. J. Biol. Chem. 2011, 286, 24394–24406. [Google Scholar] [CrossRef]
- Zhao, H.; Rinaldi, A.C.; Di Giulio, A.; Simmaco, M.; Kinnunen, P.K. Interactions of the antimicrobial peptides temporins with model biomembranes. Comparison of temporins B and L. Biochemistry 2002, 41, 4425–4436. [Google Scholar] [CrossRef]
- Mahalka, A.K.; Kinnunen, P.K. Binding of amphipathic alpha-helical antimicrobial peptides to lipid membranes: Lessons from temporins B and L. Biochim. Biophys. Acta 2009, 1788, 1600–1609. [Google Scholar] [CrossRef]
- Domanov, Y.A.; Kinnunen, P.K. Antimicrobial peptides temporins B and L induce formation of tubular lipid protrusions from supported phospholipid bilayers. Biophys. J. 2006, 91, 4427–4439. [Google Scholar] [CrossRef]
- Zhang, S.; Ma, M.; Shao, Z.; Zhang, J.; Fu, L.; Li, X.; Fang, W.; Gao, L. Structure and Formation Mechanism of Antimicrobial Peptides Temporin B- and L-Induced Tubular Membrane Protrusion. Int. J. Mol. Sci. 2021, 22, 11015. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Epand, R.F.; Rosenfeld, Y.; Peleg, A.; Barra, D.; Epand, R.M.; Shai, Y. Lipopolysaccharide, a key molecule involved in the synergism between temporins in inhibiting bacterial growth and in endotoxin neutralization. J. Biol. Chem. 2008, 283, 22907–22917. [Google Scholar] [CrossRef]
- Avitabile, C.; D’Andrea, L.D.; Saviano, M.; Olivieri, M.; Cimmino, A.; Romanelli, A. Binding studies of antimicrobial peptides to Escherichia coli cells. Biochem. Biophys. Res. Commun. 2016, 478, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Bezzerri, V.; Avitabile, C.; Dechecchi, M.C.; Lampronti, I.; Borgatti, M.; Montagner, G.; Cabrini, G.; Gambari, R.; Romanelli, A. Antibacterial and anti-inflammatory activity of a temporin B peptide analogue on an in vitro model of cystic fibrosis. J. Pept. Sci. 2014, 20, 822–830. [Google Scholar] [CrossRef]
- Hurley, K.A.; Santos, T.M.A.; Nepomuceno, G.M.; Huynh, V.; Shaw, J.T.; Weibel, D.B. Targeting the Bacterial Division Protein FtsZ. J. Med. Chem. 2016, 59, 6975–6998. [Google Scholar] [CrossRef] [PubMed]
- Kakar, A.; Holzknecht, J.; Dubrac, S.; Gelmi, M.L.; Romanelli, A.; Marx, F. New Perspectives in the Antimicrobial Activity of the Amphibian Temporin B: Peptide Analogs Are Effective Inhibitors of Candida albicans Growth. J. Fungi. 2021, 7, 457. [Google Scholar] [CrossRef] [PubMed]
- Kakar, A.; Sastre-Velasquez, L.E.; Hess, M.; Galgoczy, L.; Papp, C.; Holzknecht, J.; Romanelli, A.; Varadi, G.; Malanovic, N.; Marx, F. The Membrane Activity of the Amphibian Temporin B Peptide Analog TB_KKG6K Sheds Light on the Mechanism That Kills Candida albicans. mSphere 2022, 7, e0029022. [Google Scholar] [CrossRef]
- Grassi, L.; Maisetta, G.; Maccari, G.; Esin, S.; Batoni, G. Analogs of the Frog-skin Antimicrobial Peptide Temporin 1Tb Exhibit a Wider Spectrum of Activity and a Stronger Antibiofilm Potential as Compared to the Parental Peptide. Front. Chem. 2017, 5, 24. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Maisetta, G.; Di Luca, M.; Gaddi, L.M.; Esin, S.; Florio, W.; Brancatisano, F.L.; Barra, D.; Campa, M.; Batoni, G. Comparative analysis of the bactericidal activities of amphibian peptide analogues against multidrug-resistant nosocomial bacterial strains. Antimicrob. Agents Chemother. 2008, 52, 85–91. [Google Scholar] [CrossRef]
- D’Auria, F.D.; Casciaro, B.; De Angelis, M.; Marcocci, M.E.; Palamara, A.T.; Nencioni, L.; Mangoni, M.L. Antifungal Activity of the Frog Skin Peptide Temporin G and Its Effect on Candida albicans Virulence Factors. Int. J. Mol. Sci. 2022, 23, 6345. [Google Scholar] [CrossRef]
- Casciaro, B.; Loffredo, M.R.; Cappiello, F.; Fabiano, G.; Torrini, L.; Mangoni, M.L. The Antimicrobial Peptide Temporin G: Anti-Biofilm, Anti-Persister Activities, and Potentiator Effect of Tobramycin Efficacy Against Staphylococcus aureus. Int. J. Mol. Sci. 2020, 21, 9410. [Google Scholar] [CrossRef]
- De Angelis, M.; Casciaro, B.; Genovese, A.; Brancaccio, D.; Marcocci, M.E.; Novellino, E.; Carotenuto, A.; Palamara, A.T.; Mangoni, M.L.; Nencioni, L. Temporin G, an amphibian antimicrobial peptide against influenza and parainfluenza respiratory viruses: Insights into biological activity and mechanism of action. FASEB J. 2021, 35, e21358. [Google Scholar] [CrossRef]
- Marcocci, M.E.; Jackowska, B.G.; Prezioso, C.; Protto, V.; De Angelis, M.; Di Leva, F.S.; Casciaro, B.; Carotenuto, A.; Mangoni, M.L.; Palamara, A.T.; et al. The Inhibition of DNA Viruses by the Amphibian Antimicrobial Peptide Temporin G: A Virological Study Addressing HSV-1 and JPCyV. Int. J. Mol. Sci. 2022, 23, 7194. [Google Scholar] [CrossRef]
- Di Somma, A.; Recupido, F.; Cirillo, A.; Romano, A.; Romanelli, A.; Caserta, S.; Guido, S.; Duilio, A. Antibiofilm Properties of Temporin-L on Pseudomonas fluorescens in Static and In-Flow Conditions. Int. J. Mol. Sci. 2020, 21, 8526. [Google Scholar] [CrossRef] [PubMed]
- Zannella, C.; Chianese, A.; Palomba, L.; Marcocci, M.E.; Bellavita, R.; Merlino, F.; Grieco, P.; Folliero, V.; De Filippis, A.; Mangoni, M.; et al. Broad-Spectrum Antiviral Activity of the Amphibian Antimicrobial Peptide Temporin L and Its Analogs. Int. J. Mol. Sci. 2022, 23, 2060. [Google Scholar] [CrossRef] [PubMed]
- Giacometti, A.; Cirioni, O.; Ghiselli, R.; Mocchegiani, F.; Orlando, F.; Silvestri, C.; Bozzi, A.; Di Giulio, A.; Luzi, C.; Mangoni, M.L.; et al. Interaction of antimicrobial peptide temporin L with lipopolysaccharide in vitro and in experimental rat models of septic shock caused by gram-negative bacteria. Antimicrob. Agents Chemother. 2006, 50, 2478–2486. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, A.C.; Mangoni, M.L.; Rufo, A.; Luzi, C.; Barra, D.; Zhao, H.; Kinnunen, P.K.; Bozzi, A.; Di Giulio, A.; Simmaco, M. Temporin L: Antimicrobial, haemolytic and cytotoxic activities, and effects on membrane permeabilization in lipid vesicles. Biochem. J. 2002, 368 Pt 1, 91–100. [Google Scholar] [CrossRef]
- Zhao, H.; Kinnunen, P.K. Binding of the antimicrobial peptide temporin L to liposomes assessed by Trp fluorescence. J. Biol. Chem. 2002, 277, 25170–25177. [Google Scholar] [CrossRef] [PubMed]
- Avitabile, C.; D’Andrea, L.D.; Saviano, M.; Romanelli, A. Determination of the secondary structure of peptides in the presence of Gram positive bacterium S. epidermidis cells. RSC Adv. 2016, 6, 51407–51410. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Papo, N.; Barra, D.; Simmaco, M.; Bozzi, A.; Di Giulio, A.; Rinaldi, A.C. Effects of the antimicrobial peptide temporin L on cell morphology, membrane permeability and viability of Escherichia coli. Biochem. J. 2004, 380 Pt 3, 859–865. [Google Scholar] [CrossRef]
- Manzo, G.; Ferguson, P.M.; Hind, C.K.; Clifford, M.; Gustilo, V.B.; Ali, H.; Bansal, S.S.; Bui, T.T.; Drake, A.F.; Atkinson, R.A.; et al. Temporin L and aurein 2.5 have identical conformations but subtly distinct membrane and antibacterial activities. Sci. Rep. 2019, 9, 10934. [Google Scholar] [CrossRef]
- Di Somma, A.; Avitabile, C.; Cirillo, A.; Moretta, A.; Merlino, A.; Paduano, L.; Duilio, A.; Romanelli, A. The antimicrobial peptide Temporin L impairs E. coli cell division by interacting with FtsZ and the divisome complex. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129606. [Google Scholar] [CrossRef]
- Brancaccio, D.; Pizzo, E.; Cafaro, V.; Notomista, E.; De Lise, F.; Bosso, A.; Gaglione, R.; Merlino, F.; Novellino, E.; Ungaro, F.; et al. Antimicrobial peptide Temporin-L complexed with anionic cyclodextrins results in a potent and safe agent against sessile bacteria. Int. J. Pharm. 2020, 584, 119437. [Google Scholar] [CrossRef] [PubMed]
- Mizukami, S.; Kashibe, M.; Matsumoto, K.; Hori, Y.; Kikuchi, K. Enzyme-triggered compound release using functionalized antimicrobial peptide derivatives. Chem. Sci. 2017, 8, 3047–3053. [Google Scholar] [CrossRef]
- Grieco, P.; Carotenuto, A.; Auriemma, L.; Saviello, M.R.; Campiglia, P.; Gomez-Monterrey, I.M.; Marcellini, L.; Luca, V.; Barra, D.; Novellino, E.; et al. The effect of D-amino acid substitution on the selectivity of temporin L towards target cells: Identification of a potent anti-Candida peptide. BBA-Biomembr. 2013, 1828, 652–660. [Google Scholar] [CrossRef]
- Bellavita, R.; Raucci, F.; Merlino, F.; Piccolo, M.; Ferraro, M.G.; Irace, C.; Santamaria, R.; Iqbal, A.J.; Novellino, E.; Grieco, P.; et al. Temporin L-derived peptide as a regulator of the acute inflammatory response in zymosan-induced peritonitis. Biomed. Pharm. 2020, 123, 109788. [Google Scholar] [CrossRef] [PubMed]
- Buommino, E.; Carotenuto, A.; Antignano, I.; Bellavita, R.; Casciaro, B.; Loffredo, M.R.; Merlino, F.; Novellino, E.; Mangoni, M.L.; Nocera, F.P.; et al. The Outcomes of Decorated Prolines in the Discovery of Antimicrobial Peptides from Temporin-L. ChemMedChem 2019, 14, 1283–1290. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Carotenuto, A.; Auriemma, L.; Saviello, M.R.; Campiglia, P.; Gomez-Monterrey, I.; Malfi, S.; Marcellini, L.; Barra, D.; Novellino, E.; et al. Structure-activity relationship, conformational and biological studies of temporin L analogues. J. Med. Chem. 2011, 54, 1298–1307. [Google Scholar] [CrossRef]
- Bellavita, R.; Buommino, E.; Casciaro, B.; Merlino, F.; Cappiello, F.; Marigliano, N.; Saviano, A.; Maione, F.; Santangelo, R.; Mangoni, M.L.; et al. Synthetic Amphipathic beta-Sheet Temporin-Derived Peptide with Dual Antibacterial and Anti-Inflammatory Activities. Antibiotics 2022, 11, 1285. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Kumar, A.; Tripathi, A.K.; Tandon, A.; Ghosh, J.K. Modulation of anti-endotoxin property of Temporin L by minor amino acid substitution in identified phenylalanine zipper sequence. Biochem. J. 2016, 473, 4045–4062. [Google Scholar] [CrossRef]
- Kumari, T.; Verma, D.P.; Afshan, T.; Verma, N.K.; Pant, G.; Ali, M.; Shukla, P.K.; Mitra, K.; Ghosh, J.K. A Noncytotoxic Temporin L Analogue with In Vivo Antibacterial and Antiendotoxin Activities and a Nonmembrane-Lytic Mode of Action. ACS Infect. Dis. 2020, 6, 2369–2385. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Luo, W.; Zhong, H.; Wang, M.; Song, Y.; Deng, S.; Zhang, Y. Molecular cloning and characterization of antimicrobial peptides from skin of Hylarana guentheri. Acta Biochim. Biophys. Sin. (Shanghai) 2017, 49, 450–457. [Google Scholar] [CrossRef]
- Mishra, B.; Wang, G. Ab initio design of potent anti-MRSA peptides based on database filtering technology. J. Am. Chem. Soc. 2012, 134, 12426–12429. [Google Scholar] [CrossRef]
- Wei, H.; Xie, Z.; Tan, X.; Guo, R.; Song, Y.; Xie, X.; Wang, R.; Li, L.; Wang, M.; Zhang, Y. Temporin-Like Peptides Show Antimicrobial and Anti-Biofilm Activities against Streptococcus mutans with Reduced Hemolysis. Molecules 2020, 25, 5724. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Xie, S.; Jin, X.; Zhu, R.; Wei, S.; Song, Y.; Xie, X.; Wang, R.; Li, L.; Wang, M.; et al. Four temporin-derived peptides exhibit antimicrobial and antibiofilm activities against methicillin-resistant. Acta Biochim. Biophys. Sin. (Shanghai) 2022, 54, 350–360. [Google Scholar]
- Halverson, T.; Basir, Y.J.; Knoop, F.C.; Conlon, J.M. Purification and characterization of antimicrobial peptides from the skin of the North American green frog Rana clamitans. Peptides 2000, 21, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.F.; Lips, K.R.; Knoop, F.C.; Fritzsch, B.; Miller, C.; Conlon, J.M. Antimicrobial peptides and protease inhibitors in the skin secretions of the crawfish frog, Rana areolata. Biochim. Biophys. Acta 2002, 1601, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Wang, X.; Lushnikova, T.; Zhang, Y.; Golla, R.M.; Narayana, J.L.; Wang, C.; McGuire, T.R.; Wang, G. Antibacterial, antifungal, anticancer activities and structural bioinformatics analysis of six naturally occurring temporins. Peptides 2018, 106, 9–20. [Google Scholar] [CrossRef]
- Conlon, J.M.; Meetani, M.A.; Coquet, L.; Jouenne, T.; Leprince, J.; Vaudry, H.; Kolodziejek, J.; Nowotny, N.; King, J.D. Antimicrobial peptides from the skin secretions of the New World frogs Lithobates capito and Lithobates warszewitschii (Ranidae). Peptides 2009, 30, 1775–1781. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yu, Z.; Hu, Y.; Li, F.; Liu, L.; Zheng, H.; Meng, H.; Yang, S.; Yang, X.; Liu, J. Novel antimicrobial peptides isolated from the skin secretions of Hainan odorous frog, Odorrana hainanensis. Peptides 2012, 35, 285–290. [Google Scholar] [CrossRef]
- Chen, T.; Zhou, M.; Rao, P.; Walker, B.; Shaw, C. The Chinese bamboo leaf odorous frog (Rana (Odorrana) versabilis) and North American Rana frogs share the same families of skin antimicrobial peptides. Peptides 2006, 27, 1738–1744. [Google Scholar] [CrossRef]
- Abbassi, F.; Oury, B.; Blasco, T.; Sereno, D.; Bolbach, G.; Nicolas, P.; Hani, K.; Amiche, M.; Ladram, A. Isolation, characterization and molecular cloning of new temporins from the skin of the North African ranid Pelophylax saharica. Peptides 2008, 29, 1526–1533. [Google Scholar] [CrossRef] [PubMed]
- Abbassi, F.; Galanth, C.; Amiche, M.; Saito, K.; Piesse, C.; Zargarian, L.; Hani, K.; Nicolas, P.; Lequin, O.; Ladram, A. Solution structure and model membrane interactions of temporins-SH, antimicrobial peptides from amphibian skin. A NMR spectroscopy and differential scanning calorimetry study. Biochemistry 2008, 47, 10513–10525. [Google Scholar] [CrossRef] [PubMed]
- Abbassi, F.; Raja, Z.; Oury, B.; Gazanion, E.; Piesse, C.; Sereno, D.; Nicolas, P.; Foulon, T.; Ladram, A. Antibacterial and leishmanicidal activities of temporin-SHd, a 17-residue long membrane-damaging peptide. Biochimie 2013, 95, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Andre, S.; Raja, Z.; Humblot, V.; Piesse, C.; Foulon, T.; Sereno, D.; Oury, B.; Ladram, A. Functional Characterization of Temporin-SHe, a New Broad-Spectrum Antibacterial and Leishmanicidal Temporin-SH Paralog from the Sahara Frog (Pelophylax saharicus). Int. J. Mol. Sci. 2020, 21, 6713. [Google Scholar] [CrossRef]
- Abbassi, F.; Lequin, O.; Piesse, C.; Goasdoue, N.; Foulon, T.; Nicolas, P.; Ladram, A. Temporin-SHf, a new type of phe-rich and hydrophobic ultrashort antimicrobial peptide. J. Biol. Chem. 2010, 285, 16880–16892. [Google Scholar] [CrossRef] [PubMed]
- Olleik, H.; Baydoun, E.; Perrier, J.; Hijazi, A.; Raymond, J.; Manzoni, M.; Dupuis, L.; Pauleau, G.; Goudard, Y.; Villeon, B.; et al. Temporin-SHa and Its Analogs as Potential Candidates for the Treatment of Helicobacter pylori. Biomolecules 2019, 9, 598. [Google Scholar] [CrossRef] [PubMed]
- Crepin, A.; Jegou, J.F.; Andre, S.; Ecale, F.; Croitoru, A.; Cantereau, A.; Berjeaud, J.M.; Ladram, A.; Verdon, J. In vitro and intracellular activities of frog skin temporins against Legionella pneumophila and its eukaryotic hosts. Sci. Rep. 2020, 10, 3978. [Google Scholar] [CrossRef]
- Eggimann, G.A.; Sweeney, K.; Bolt, H.L.; Rozatian, N.; Cobb, S.L.; Denny, P.W. The role of phosphoglycans in the susceptibility of Leishmania mexicana to the temporin family of anti-microbial peptides. Molecules 2015, 20, 2775–2785. [Google Scholar] [CrossRef]
- Roy, M.; Lebeau, L.; Chessa, C.; Damour, A.; Ladram, A.; Oury, B.; Boutolleau, D.; Bodet, C.; Leveque, N. Comparison of Anti-Viral Activity of Frog Skin Anti-Microbial Peptides Temporin-Sha and [K3]SHa to LL-37 and Temporin-Tb against Herpes Simplex Virus Type 1. Viruses 2019, 11, 77. [Google Scholar] [CrossRef]
- Maharjan, R.; Khan, A.I.; Nadeem-Ul-Haque, M.; Maresca, M.; Choudhary, M.I.; Shaheen, F.; Simjee, S.U. Serum Stable and Low Hemolytic Temporin-SHa Peptide Analogs Disrupt Cell Membrane of Methicillin-Resistant Staphylococcus aureus (MRSA). Probiotics Antimicrob. Proteins 2022, 14, 391–405. [Google Scholar] [CrossRef]
- Shaheen, F.; Nadeem-Ul-Haque, M.; Ahmed, A.; Simjee, S.U.; Ganesan, A.; Jabeen, A.; Shah, Z.A.; Choudhary, M.I. Synthesis of breast cancer targeting conjugate of temporin-SHa analog and its effect on pro- and anti-apoptotic protein expression in MCF-7 cells. Peptides 2018, 106, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Raja, Z.; Andre, S.; Abbassi, F.; Humblot, V.; Lequin, O.; Bouceba, T.; Correia, I.; Casale, S.; Foulon, T.; Sereno, D.; et al. Insight into the mechanism of action of temporin-SHa, a new broad-spectrum antiparasitic and antibacterial agent. PLoS ONE 2017, 12, e0174024. [Google Scholar] [CrossRef] [PubMed]
- Brunet, K.; Verdon, J.; Ladram, A.; Arnault, S.; Rodier, M.H.; Cateau, E. Antifungal activity of [K(3)]temporin-SHa against medically relevant yeasts and moulds. Can. J. Microbiol. 2022, 68, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Andre, S.; Washington, S.K.; Darby, E.; Vega, M.M.; Filip, A.D.; Ash, N.S.; Muzikar, K.A.; Piesse, C.; Foulon, T.; O’Leary, D.J.; et al. Structure-Activity Relationship-based Optimization of Small Temporin-SHf Analogs with Potent Antibacterial Activity. ACS Chem. Biol. 2015, 10, 2257–2266. [Google Scholar] [CrossRef] [PubMed]
- Piras, A.M.; Maisetta, G.; Sandreschi, S.; Gazzarri, M.; Bartoli, C.; Grassi, L.; Esin, S.; Chiellini, F.; Batoni, G. Chitosan nanoparticles loaded with the antimicrobial peptide temporin B exert a long-term antibacterial activity in vitro against clinical isolates of Staphylococcus epidermidis. Front. Microbiol. 2015, 6, 372. [Google Scholar] [CrossRef]
- Parilti, R.; Caprasse, J.; Riva, R.; Alexandre, M.; Vandegaart, H.; Bebrone, C.; Dupont-Gillain, C.; Howdle, S.M.; Jerome, C. Antimicrobial peptide encapsulation and sustained release from polymer network particles prepared in supercritical carbon dioxide. J. Colloid. Interf. Sci. 2018, 532, 112–117. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, H.; Wang, B.; Wang, R.X.; Wang, C.; Shang, D.J. Characterization and Anticancer Activity of a Folic Acid Conjugated and Cationic Peptide L-K6 Encapsulated Cancer-Targeting Liposomal Drug Delivery System. Int. J. Pept. Res. Ther. 2022, 28, 82. [Google Scholar] [CrossRef]
- Rai, A.; Ferrao, R.; Palma, P.; Patricio, T.; Parreira, P.; Anes, E.; Tonda-Turo, C.; Martins, M.C.L.; Alves, N.; Ferreira, L. Antimicrobial peptide-based materials: Opportunities and challenges. J. Mater. Chem. B 2022, 10, 2384–2429. [Google Scholar] [CrossRef]
- Lombana, A.; Raja, Z.; Casale, S.; Pradier, C.M.; Foulon, T.; Ladram, A.; Humblot, V. Temporin-SHa peptides grafted on gold surfaces display antibacterial activity. J. Pept. Sci. 2014, 20, 563–569. [Google Scholar] [CrossRef]
- Oger, P.C.; Piesse, C.; Ladram, A.; Humblot, V. Engineering of Antimicrobial Surfaces by Using Temporin Analogs to Tune the Biocidal/antiadhesive Effect. Molecules 2019, 24, 814. [Google Scholar] [CrossRef]
- Masurier, N.; Tissot, J.B.; Boukhriss, D.; Jebors, S.; Pinese, C.; Verdie, P.; Amblard, M.; Mehdi, A.; Martinez, J.; Humblot, V.; et al. Site-specific grafting on titanium surfaces with hybrid temporin antibacterial peptides. J. Mater. Chem. B 2018, 6, 1782–1790. [Google Scholar] [CrossRef] [PubMed]
- Sperandeo, P.; Bosco, F.; Clerici, F.; Polissi, A.; Gelmi, M.L.; Romanelli, A. Covalent Grafting of Antimicrobial Peptides onto Microcrystalline Cellulose. ACS Appl. Bio Mater. 2020, 3, 4895–4901. [Google Scholar] [CrossRef] [PubMed]
- Aslanli, A.; Lyagin, I.; Efremenko, E. Charges’ interaction in polyelectrolyte (nano)complexing of His(6)-OPH with peptides: Unpredictable results due to imperfect or useless concept? Int. J. Biol. Macromol. 2019, 140, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Aslanli, A.; Stepanov, N.; Razheva, T.; Podorozhko, E.A.; Lyagin, I.; Lozinsky, V.I.; Efremenko, E. Enzymatically Functionalized Composite Materials Based on Nanocellulose and Poly(Vinyl Alcohol) Cryogel and Possessing Antimicrobial Activity. Materials 2019, 12, 3619. [Google Scholar] [CrossRef]
- Aslanli, A.; Lyagin, I.; Stepanov, N.; Presnov, D.; Efremenko, E. Bacterial Cellulose Containing Combinations of Antimicrobial Peptides with Various QQ Enzymes as a Prototype of an “Enhanced Antibacterial” Dressing: In Silico and In Vitro Data. Pharmaceutics 2020, 12, 1155. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Cheng, Z.; Wen, R.; Zhao, X.; Yu, X.; Sun, L.; Zhang, Y.; Han, Z.; Yuan, Y.; Kang, L. Novel SA@Ca2+/RCSPs core-shell structure nanofibers by electrospinning for wound dressings. RSC Adv. 2018, 8, 15558–15566. [Google Scholar] [CrossRef]
- Ji, X.; Li, R.; Liu, G.M.; Jia, W.Y.; Sun, M.L.; Liu, Y.; Luo, Y.G.; Cheng, Z.Q. Phase separation-based electrospun Janus nanofibers loaded with Rana chensinensis skin peptides/silver nanoparticles for wound healing. Mater. Des. 2021, 207, 109864. [Google Scholar] [CrossRef]
- Ji, X.; Liu, G.M.; Cui, Y.T.; Jia, W.Y.; Luo, Y.G.; Cheng, Z.Q. A hybrid system of hydrogel/frog egg-like microspheres accelerates wound healing via sustained delivery ofRCSPs. J. Appl. Polym. Sci. 2020, 137, 718377. [Google Scholar] [CrossRef]
- da Silva-Junio, A.G.; Frias, I.A.M.; Lima-Neto, R.G.; Migliolo, L.; PS, E.S.; Oliveira, M.D.L.; Andrade, C.A.S. Electrochemical biosensor based on Temporin-PTA peptide for detection of microorganisms. J. Pharm. Biomed. Anal. 2022, 216, 114788. [Google Scholar] [CrossRef]
- Zhao, N.; Bardine, C.; Lourenco, A.L.; Wang, Y.H.; Huang, Y.; Cleary, S.J.; Wilson, D.M.; Oh, D.Y.; Fong, L.; Looney, M.R.; et al. In Vivo Measurement of Granzyme Proteolysis from Activated Immune Cells with PET. ACS Cent. Sci. 2021, 7, 1638–1649. [Google Scholar] [CrossRef] [PubMed]



| Genus | Specie | Peptide Name | Sequence a |
|---|---|---|---|
| Amolops | |||
| A. chunganensis | Temporin CG1 | FLPFVGNLLKGLL | |
| Temporin CG2 | FFPIVGKLLSGLF | ||
| Temporin CG3 | FLPIVGKLLSGLF | ||
| Temporin CG4 | FLPILGNLLNGLL | ||
| Temporin CG5 | FLPFVGNLLNGLL | ||
| A. daiyunensis | Temporin DY1 | FFPMLADLVSKIF | |
| A. jingdongensis | Temporin AJ1 | FLPIVTGLLSSLL | |
| Temporin AJ2 | FFPIGGKLLFGLL | ||
| Temporin AJ3 | FFPIVGKLLSGLL | ||
| Temporin AJ4 | FFPIVGKLLFGLL | ||
| Temporin AJ5 | FPPIVGKLLFGLL | ||
| Temporin AJ6 | FLPIVGKLLSGLL | ||
| Temporin AJ7 | FLPIVGKLPSGLL | ||
| Temporin AJ8 | FFPIVGKRLYGLL | ||
| Temporin AJ9 | FLPIVGKLLSGLTGLL | ||
| Temporin AJ10 | FLPIVGKLLSGLSGLL | ||
| Temporin AJ11 | FFPIGGKLLSGLTGLL | ||
| A. lifanensis | Temporin LF1 | FLPFVGKLLSGLL | |
| A. loloensis | Temporin ALa | FLPIVGKLLSGLSGLL | |
| Temporin ALb | FLPIVGKLLSGLL | ||
| Temporin ALc | LLPIVGKGLSGKL | ||
| Temporin ALd | FLPIAGKLLSGLSGLL | ||
| Temporin ALe | FFPIVGKLLFGLSGLL | ||
| Temporin ALf | FFPIVGKLLSGLSGLL | ||
| Temporin ALg | FFPIVGKLLFGLFGLL | ||
| Temporin ALh | FLPIVGKLLSGLSGLS | ||
| Temporin ALi | FFPIVGKLLSGLL | ||
| Temporin ALj | FFPIVGKLLFGLL | ||
| Temporin ALk | FFPIVGKLLS | ||
| Euphlyctis | |||
| E. cyanophlyctis | Temporin ECa | FLPGLLAGLL | |
| Hylarana | |||
| H. guentheri | Temporin GH | FLPLLFGAISHLL | |
| Temporin GHa | FLQHIIGALGHLF | ||
| Temporin GHb | FIHHIIGALGHLF | ||
| Temporin GHc | FLQHIIGALTHIF | ||
| Temporin GHd | FLQHIIGALSHFF | ||
| Temporin GUa | FLQHIIGALSHIF | ||
| Temporin GUb | FLPLLFGAISHIL | ||
| Temporin GUc | FFPLIFGALSSILPKIL | ||
| H. latouchii | Temporin LT1 | FLPGLIAGIAKML | |
| Temporin LT2 | FLPIALKALGSIFPKIL | ||
| Temporin LTa | FFPLVLGALGSILPKIF | ||
| Temporin LTb | FIITGLVRGLTKLF | ||
| Temporin LTc | SLSRFLSFLKIVYPPAF | ||
| Temporin LTe | FLAGLIGGLAKML | ||
| Temporin HLa | FFPLIFGALSSILPKIL | ||
| Temporin HLb | FLQHIIGALSHIF | ||
| H. picturata | Temporin PTA | FFGSVLKLIPKIL | |
| H. taipehensis | Temporin TP1 | FLPVLGKVIKLVGGLL | |
| Temporin TP2 | FLPLLVGAISSILPKIF | ||
| Temporin TP3 | FLPLLFGALSTLLPKIF | ||
| H. spinulosa | Temporin SN1 | FFPFLLGALGSLLPKIF | |
| Temporin SN2 | FITGLIGGLMKAL | ||
| Temporin SN3 | FISGLIGGLMKAL | ||
| Temporin SN4 | FITGLISGLMKAL | ||
| Temporin SN5 | FFPLVLGALGSILPKIF | ||
| H. maosonensis | Temporin MS1 | FLTGLIGGLMKALGK | |
| Temporin MS4 | FLSGLIGGLAKMLGK | ||
| Limnonectes | |||
| L. kuhlii | Temporin LK1 | FFPLLFGALSSMMPKLF | |
| Lithobates | |||
| L. areolatus | Temporin 1AR | FLPIVGRLISGLL | |
| L. capito | Temporin CPa | IPPFIKKVLTTVF | |
| Temporin CPb | FLPIVGRLISGIL | ||
| L. catesbeianus | Temporin La | LLRHVVKILEKYL | |
| L. clamitans | Temporin 1Ca | FLPFLAKILTGVL | |
| Temporin 1Cb | FLPLFASLIGKLL | ||
| Temporin 1Cc | FLPFLASLLTKVL | ||
| Temporin 1Cd | FLPFLASLLSKVL | ||
| Temporin 1Ce | FLPFLATLLSKVL | ||
| L. grylio | Temporin 1Ga | SILPTIVSFLSKVF | |
| Temporin 1Gb | SILPTIVSFLSKFL | ||
| Temporin 1Gc | SILPTIVSFLTKFL | ||
| Temporin 1Gd | FILPLIASFLSKFL | ||
| L. heckscheri | Temporin 1HKa | SIFPAIVSFLSKFL | |
| L. okaloosae | Temporin 1OLa | FLPFLKSILGKIL | |
| Temporin 1OLb | FLPFFASLLGKLL | ||
| L. onca | Temporin ONa | FLPTFGKILSGLF | |
| L. palustris | Temporin 1PLa | FLPLVGKILSGLI | |
| L. pipiens | Temporin 1P | FLPIVGKLLSGLL | |
| L. septentrionalis | Temporin 1SPa | FLPLVGKILSGLI | |
| Temporin 1SPb | FLSAITSLLGKLL | ||
| Temporin 1SPc | FLSAITSILGKLF | ||
| L. sphenacephalus | Temporin 1S | LLFGKIISRLLGN | |
| L. virgatipes | Temporin 1Va | FLSSIGKILGNLL | |
| Temporin 1Vb | FLSIIAKVLGSLF | ||
| Temporin 1Vc | FLPLVTMLLGKLF | ||
| Odorrana | |||
| O. hainanensis | Temporin HN1 | AILTTLANWARKFL | |
| Temporia HN2 | NILNTIINLAKKIL | ||
| O. versabilis | Temporin 1VE | FLPLVGKILSGLI | |
| Pelophylax | |||
| P. esculentus | Temporin PE | FLPIVAKLLSGLL | |
| Temporin 1Re | FLPGLLAGLL | ||
| Temporin 1Ec | FLPVIAGLLSKLF | ||
| Temporin 1Ee | FLPVIAGVLSKLF | ||
| P. fukienensis | Temporin PF | FLPLIAGLFGKIF | |
| P. hubeiensis | Temporin HB1 | FLPLLAGLAAKWF | |
| Temporin HB2 | FLPFLAGLFGKIF | ||
| P. nigromaculatus | Temporin 1KM | FIPLVSGLFSRLL | |
| Temporin 1GY | VIPIVSGLLSSLL | ||
| P. ridibundus | Temporin Ra | FLKPLFNAALKLLP | |
| Temporin Rb | FLPVLAGVLSRA | ||
| P. saharicus | Temporin SHa | FLSGIVGMLGKLF | |
| Temporin SHb | FLPIVTNLLSGLL | ||
| Temporin SHc | FLSHIAGFLSNLF | ||
| Temporin SHd | FLPAALAGIGGILGKLF | ||
| Temporin SHe | FLPALAGIAGLLGKIF | ||
| Temporin SHf | FFFLSRIF | ||
| Rana | |||
| R. aurora | Temporin 1AUa | FLPIIGQLLSGLL | |
| R. boylii | Temporin 1BYa | FLPIIAKVLSGLL | |
| R. cascadae | Temporin 1CSa | FLPIVGKLLSGLL | |
| Temporin 1CSb | FLPIIGKLLSGLL | ||
| Temporin 1CSc | FLPLVTGLLSGLL | ||
| Temporin 1CSd | NFLGTLVNLAKKIL | ||
| R. chensinensis | Temporin 1CEa | FVDLKKIANIINSIF | |
| Temporin 1CEb | ILPILSLIGGLL | ||
| Temporin 1CEc | IIPLPLGYFAKKT | ||
| Temporin 1CEd | ILPLIASLIGGLL | ||
| Temporin 1CEe | ILPIIGKILSTIFGK | ||
| Temporin 1CEf | ILPILGKILSTIL | ||
| Temporin 1CEg | ILPIFSWIGHLF | ||
| Temporin 1CEh | FVDLKKIANILNSIF | ||
| R. draytonii | Temporin 1DRa | HFLGTLVNLAKKIL | |
| Temporin 1DRb | NFLGTLVNLAKKIL | ||
| Temporin 1DRc | FLPIIASVLSSLL | ||
| R. dybowskii | Temporin 1DYa | FIGPIISALASLFG | |
| Temporin CDYa | LPLVGNLLNDLL | ||
| Temporin CDYb | ILPILSLIGGLL | ||
| Temporin CDYc | VLPLVGNLLNDLL | ||
| Temporin CDYd | FIGPLISALASLFKG | ||
| Temporin CDYe | FIGPIISALASLFGG | ||
| R. italica | Temporin ITa | FLGAIAQALTSLLGKL | |
| R. japonica | Temporin 1Ja | ILPLVGNLLNDLL | |
| R. luteiventris | Temporin 1La | VLPLISMALGKLL | |
| Temporin 1Lb | NFLGTLINLAKKIM | ||
| Temporin 1Lc | FLPILINLIHKGLL | ||
| R. muscosa | Temporin 1M | FLPIVGKLLSGLL | |
| R. nigromaculata | Temporin 1RNa | ILPIRSLIKKLL | |
| Temporin 1RNb | FLPLKKLRFGLL | ||
| R. nigrovittata | Temporin RN1 | FLPLVLGALSGILPKIL | |
| Temporin RN2 | FFPLLFGALSSLLPKLF | ||
| Temporin RN3 | FFPLLFGALSSHLPKLF | ||
| R. ornativentris | Temporin 1Oa | FLPLLASLFSRLL | |
| Temporin 1Ob | FLPLIGKILGTIL | ||
| Temporin 1Oc | FLPLLASLFSRLF | ||
| Temporin 1Od | FLPLLASLFSGLF | ||
| Temporin 1Oe | ILPLLGNLLNGLL | ||
| Temporin 1Of | SLLLKGLASIAKLF | ||
| Temporin 1Og | FLSSLLSKVVSLFT | ||
| R. pirica | Temporin 1PRa | ILPILGNLLNGLL | |
| Temporin 1PRb | ILPILGNLLNSLL | ||
| R. pretiosa | Temporin PRa | FLPILGNLLSGLL | |
| Temporin PRb | FLPIITNLLGKLL | ||
| Temporin PRc | NFLDTLINLAKKFI | ||
| Temporin PRe | FLPLAMALGKLL | ||
| R. sakuraii | Temporin 1Ska | FLPVILPVIGKLLNGIL | |
| Temporin 1SKb | FLPVILPVIGKLLSGIL | ||
| R. tagoi | Temporin 1TGa | FLPILGKLLSGIL | |
| Temporin 1TGb | AVDLAKIANKVLSSLF | ||
| Temporin 1TGc | FLPVILPVIGKLLSGIL | ||
| R. tagoi okiensis | Temporin TOa | FLPILGKLLSGFL | |
| Temporin TOb | FLPILGKLLSGLL | ||
| R. temporaria | Temporin A | FLPLIGRVLSGIL | |
| Temporin B | LLPIVGNLLKSLL | ||
| Temporin C | ILPILGNLLNGLL | ||
| Temporin D | LLPIVGNLLNSLL | ||
| Temporin E | VLPIIGNLLNSLL | ||
| Temporin F | FLPLIGKVLSGIL | ||
| Temporin G | FFPVIGRILNGIL | ||
| Temporin H | LSPNLLKSLL | ||
| Temporin K | LPPNLLKSLL | ||
| Temporin L | FVQWFSKFLGRIL | ||
| R. tsushimensis | Temporin 1TSa | FLGALAKIISGIF | |
| Temporin 1TSb | FLPLLGNLLNGLL | ||
| Temporin 1TSc | FLPLLGNLLRGLL | ||
| Temporin 1TSd | FLPLLASLIGGML |
| Microorganism | Antimicrobial Activity (µM) | Ref |
|---|---|---|
| E. coli D21 | LC: 11.9 | [37] |
| S. aureus Cowan 1 | LC: 2.3 | [37] |
| C. albicans ATCC 10261 | LC: 3.4 | [37] |
| Microorganism | Antimicrobial Activity | Ref |
|---|---|---|
| E. coli a | MIC: >100 μg/mL | [63] |
| S. aureus A170 | MIC 100: 25 μg/mL | [63] |
| C. albicans ATCC 10261 | LC: 4.0 μM | [37] |
| Microorganism | Antimicrobial Activity (µM) | Ref |
|---|---|---|
| S. aureus a | BC: 24 | [79] |
| C. albicans ATCC10231 | MIC: 16 | [80] |
| Microorganism | Antimicrobial Activity as MIC (µM) | Ref |
|---|---|---|
| E. coli D21 | 12 | [58] |
| S. aureus ATCC 25923 | 3 | [58] |
| C. albicans ATCC 10231 | 12 | [58] |
| Peptide | Antimicrobial Activity as MIC (µM) | |||||
|---|---|---|---|---|---|---|
| S. aureus ATCC 25923 | Ref | E. coli ATCC 25922 | Ref | C. albicans ATCC 10231 | Ref | |
| Temporin GHa | 6.8 | [103] | 26.6 | [103] | 26.6 | [103] |
| Temporin GHb | 6.8 | [103] | 13.2 | [103] | 26.4 | [103] |
| Temporin GHc | 12.9 | [103] | 104 | [103] | 104 | [103] |
| Temporin GHd | 12.7 | [103] | 12.7 | [103] | 25.5 | [103] |
| Temporin PTa | 3.12 a | [104] | 25 b | [104] | n.d. | |
| Peptide | Antimicrobial Activity as MIC (µM) | |||||
|---|---|---|---|---|---|---|
| S. aureus | Ref | E. coli ATCC 25922 | Ref | C. albicans | Ref | |
| Temporin 1Cb | 140 a | [106] | N.A. | [106] | N.A. c | [106] |
| Temporin 1Cd | 80 a | [106] | N.A. | [106] | N.A. c | [106] |
| Temporin 1Ce | 100 a | [106] | N.A. | [106] | N.A. c | [106] |
| Temporin 1AR | 15 a | [107] | 125 | [107] | N.A. c,e | [107] |
| Temporin CPa | ≥25 b | [108] | 12.5 | [108] | 25 d | [108] |
| Temporin CPb | 12.5 b | [108] | >50 | [108] | 50 d | [108] |
| Peptide | Antimicrobial Activity as MIC (µM) | |||||
|---|---|---|---|---|---|---|
| S. aureus ATCC25923 | Ref | E. coli ATCC25922 | Ref | C. albicans ATCC2002 | Ref | |
| Temporin HN1 | 37.5 | [110] | N.A. | [110] | 75 | [110] |
| Temporin HN2 | 4.8 | [110] | N.A. | [110] | 9.8 | [110] |
| Peptide | Antimicrobial Activity as MIC (µM) | |||||
|---|---|---|---|---|---|---|
| S. aureus ATCC25923 | Ref | E. coli ATCC25922 | Ref | C. albicans ATCC90028 | Ref | |
| Temporin SHa | 3 | [113] | 10 | [113] | 16 | [113] |
| Temporin SHb | 58 | [113] | 231 | [113] | >116 | [113] |
| Temporin SHc | 10 | [113] | >161 | [113] | 20 | [113] |
| Temporin SHd | 6.25 | [114] | 5 | [114] | 100 | [114] |
| Temporin SHe | 3.12 | [115] | 25 | [115] | >100 | [115] |
| Temporin SHf | 12.5 | [116] | 25 | [116] | 50 | [116] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D'Andrea, L.D.; Romanelli, A. Temporins: Multifunctional Peptides from Frog Skin. Int. J. Mol. Sci. 2023, 24, 5426. https://doi.org/10.3390/ijms24065426
D'Andrea LD, Romanelli A. Temporins: Multifunctional Peptides from Frog Skin. International Journal of Molecular Sciences. 2023; 24(6):5426. https://doi.org/10.3390/ijms24065426
Chicago/Turabian StyleD'Andrea, Luca Domenico, and Alessandra Romanelli. 2023. "Temporins: Multifunctional Peptides from Frog Skin" International Journal of Molecular Sciences 24, no. 6: 5426. https://doi.org/10.3390/ijms24065426
APA StyleD'Andrea, L. D., & Romanelli, A. (2023). Temporins: Multifunctional Peptides from Frog Skin. International Journal of Molecular Sciences, 24(6), 5426. https://doi.org/10.3390/ijms24065426