
The Cytoplasmic Region of SARAF Reduces Triple-Negative Breast Cancer Metastasis through the Regulation of Store-Operated Calcium Entry

 , , ,
, , ,
Abstract
1. Introduction
2. Results
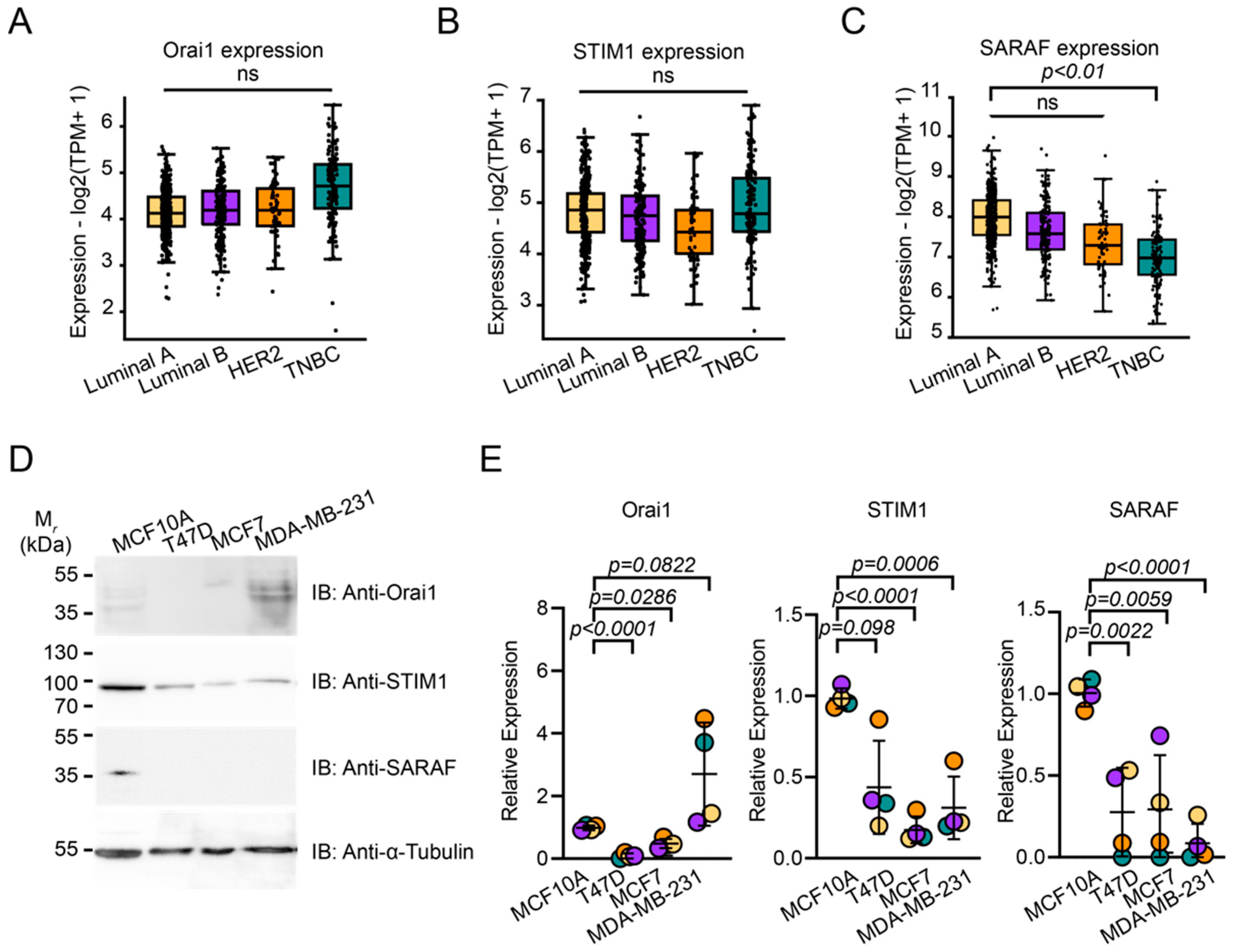
2.1. SARAF Expression Is Reduced in TNBC
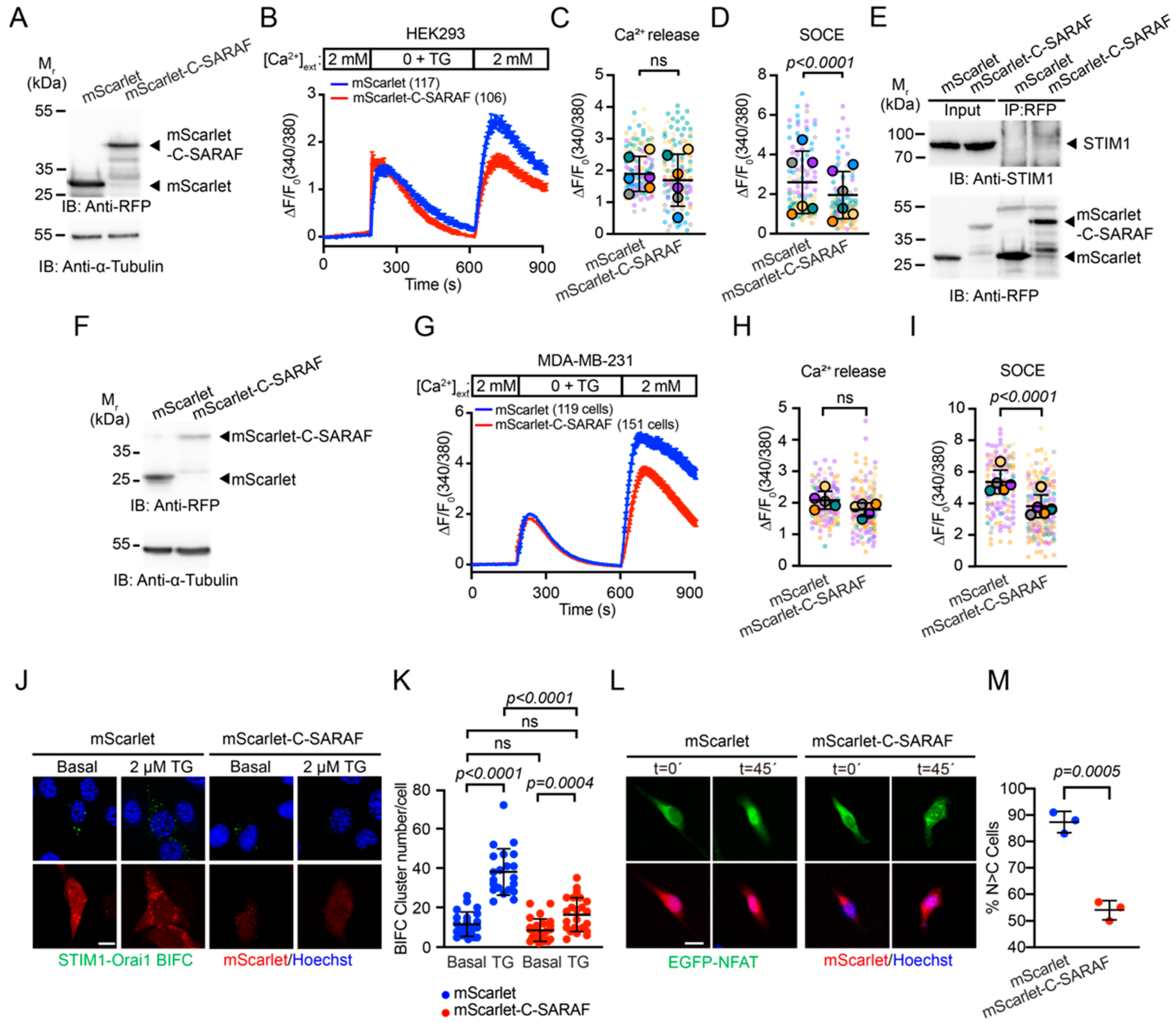
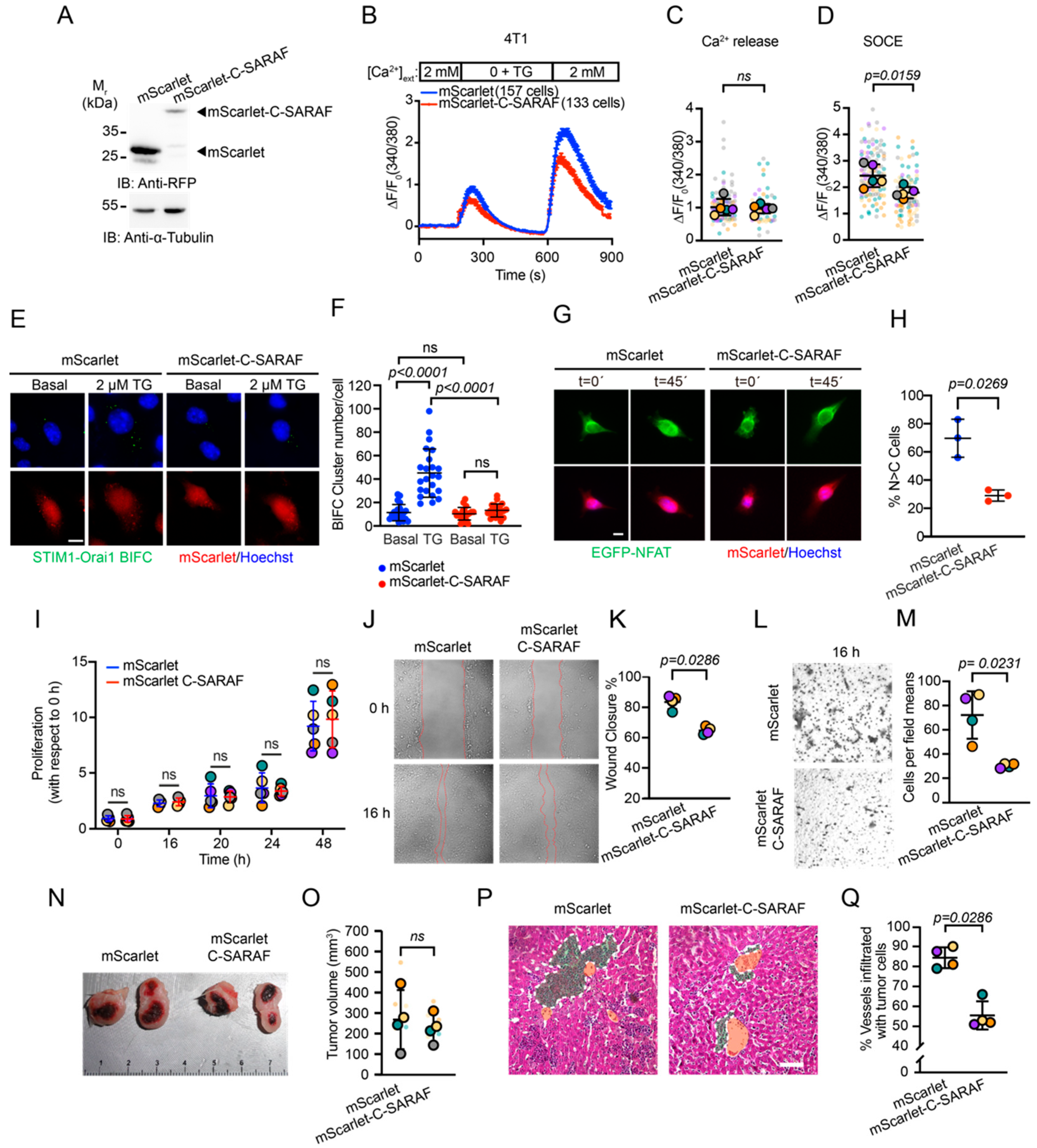
2.2. The C-Terminal SARAF Fragments Reduce Store-Operated Calcium Entry
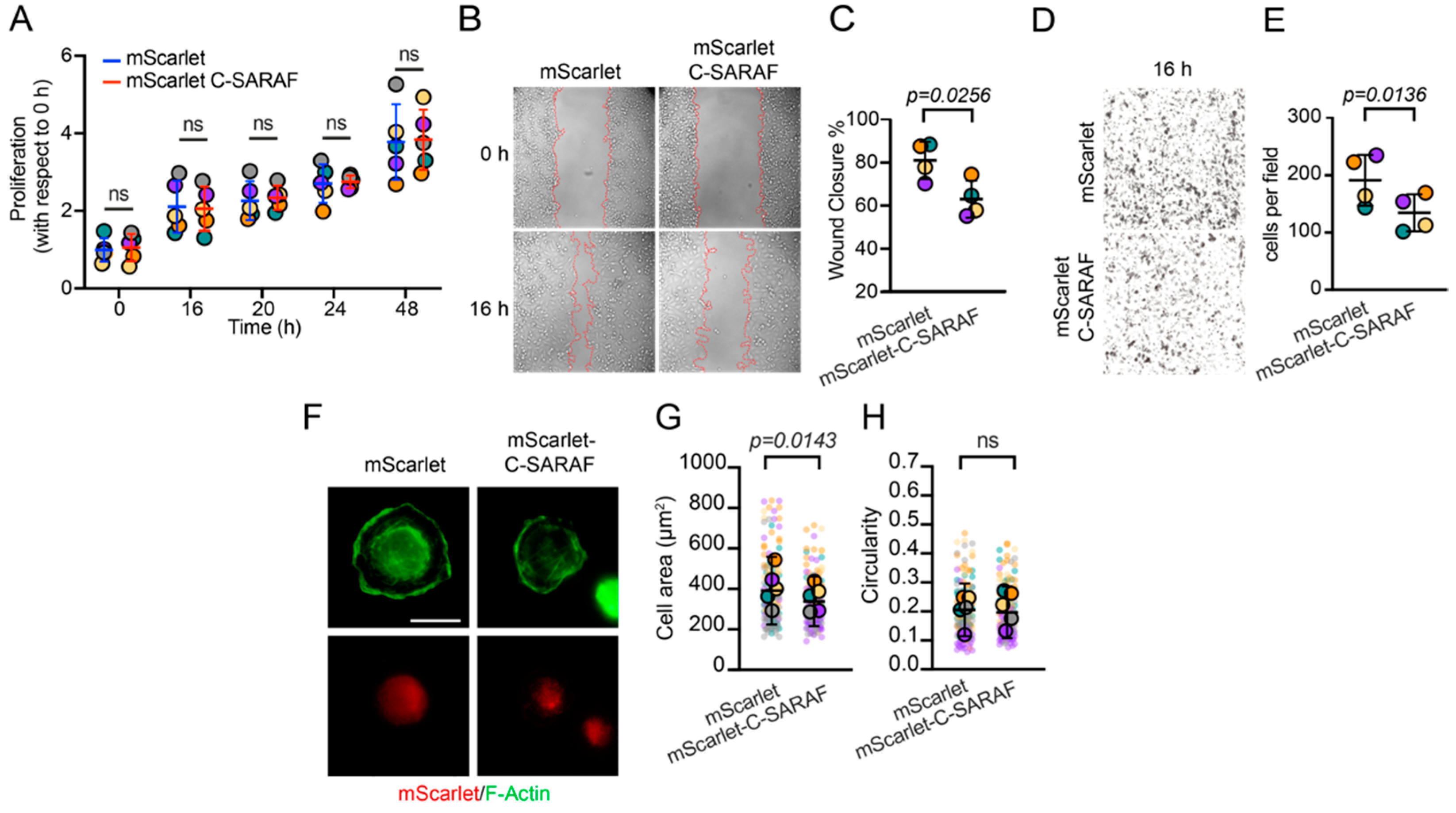
2.3. The C-Terminal SARAF Fragment Reduces Malignancy Features in Human TNBC
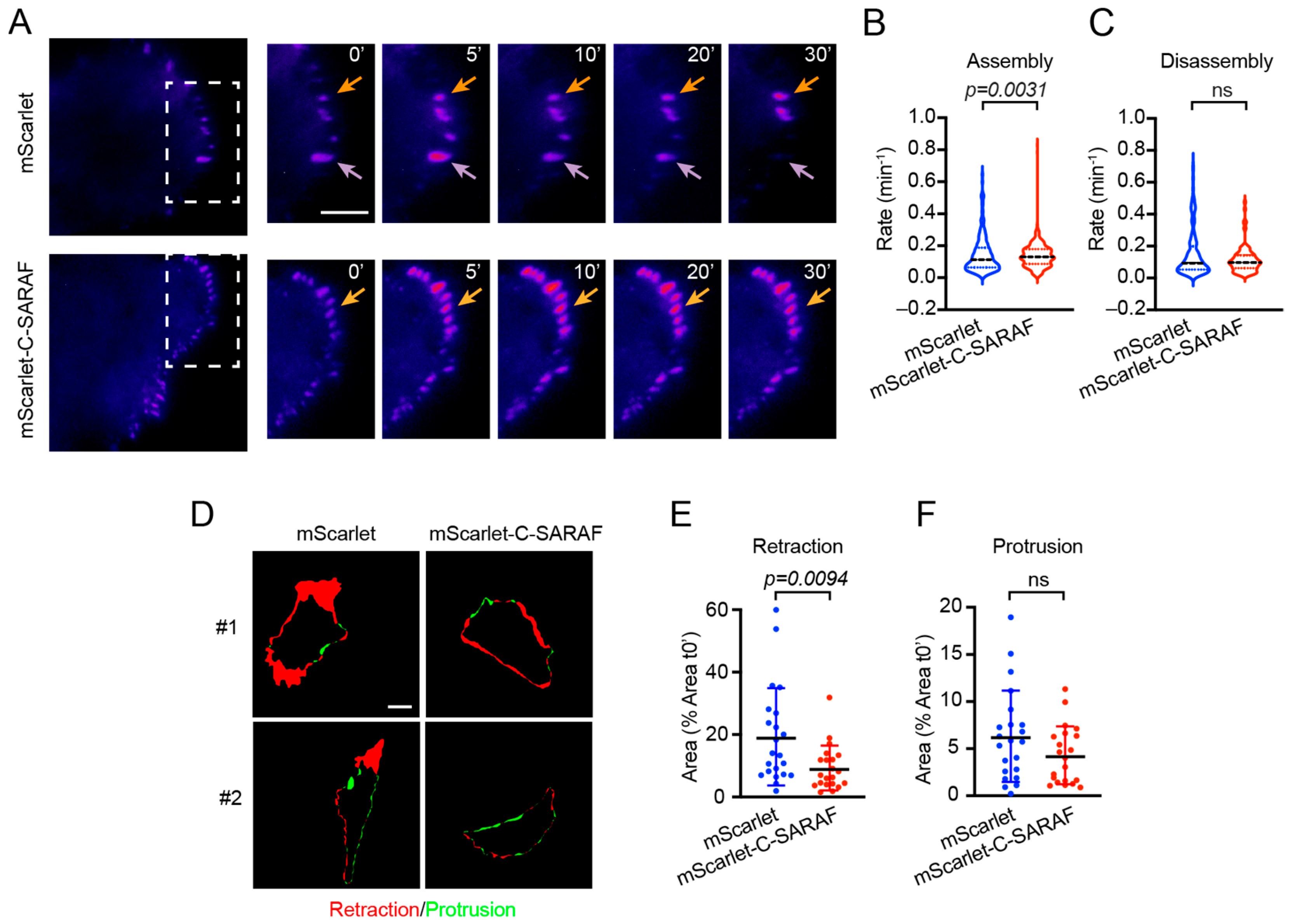
2.4. The C-Terminal SARAF Fragment Affects Dynamic Focal Adhesion and Cell Retraction
2.5. The C-Terminal SARAF Fragment Reduces SOCE Response, Malignancy Features and Metastasis, and Promotes Lymphocytic Infiltrate in the Primary Tumor TNBC in a Murine Model
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Transfections
4.2. Plasmids, Antibodies and Reagents
4.3. Ca2+ Imaging
4.4. Immunoblot Analysis
4.5. Immunoprecipitation
4.6. Biomolecular Fluorescence Complementation (BiFC)
4.7. NFAT Translocation
4.8. Focal Adhesion Dynamics
4.9. Spreading Assay
4.10. Proliferation MTT Assay
4.11. Transwell Chamber Invasion Assays
4.12. Scratch Assay
4.13. In Vivo Metastasis Model
4.14. Histology
4.15. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the Global Cancer Incidence and Mortality in 2018: GLOBOCAN Sources and Methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed]
- Harbeck, N.; Penault-Llorca, F.; Cortes, J.; Gnant, M.; Houssami, N.; Poortmans, P.; Ruddy, K.; Tsang, J.; Cardoso, F. Breast Cancer. Nat. Rev. Dis. Prim. 2019, 5, 66. [Google Scholar] [CrossRef] [PubMed]
- Badve, S.; Dabbs, D.J.; Schnitt, S.J.; Baehner, F.L.; Decker, T.; Eusebi, V.; Fox, S.B.; Ichihara, S.; Jacquemier, J.; Lakhani, S.R.; et al. Basal-like and Triple-Negative Breast Cancers: A Critical Review with an Emphasis on the Implications for Pathologists and Oncologists. Mod. Pathol. 2011, 24, 157–167. [Google Scholar] [CrossRef]
- Marchi, S.; Giorgi, C.; Galluzzi, L.; Pinton, P. Ca2+ Fluxes and Cancer. Mol. Cell 2020, 78, 1055–1069. [Google Scholar] [CrossRef]
- Monteith, G.R.; Prevarskaya, N.; Roberts-Thomson, S.J. The Calcium–Cancer Signalling Nexus. Nat. Rev. Cancer 2017, 17, 373–380. [Google Scholar] [CrossRef]
- Lee, D.; Hong, J.H. Ca2+ Signaling as the Untact Mode during Signaling in Metastatic Breast Cancer. Cancers 2021, 13, 1473. [Google Scholar] [CrossRef]
- Saldías, M.P.; Maureira, D.; Orellana-Serradell, O.; Silva, I.; Lavanderos, B.; Cruz, P.; Torres, C.; Cáceres, M.; Cerda, O. TRP Channels Interactome as a Novel Therapeutic Target in Breast Cancer. Front. Oncol. 2021, 11, 785635. [Google Scholar] [CrossRef]
- Hooper, R.; Zhang, X.; Webster, M.; Go, C.; Kedra, J.; Marchbank, K.; Gill, D.L.; Weeraratna, A.T.; Trebak, M.; Soboloff, J. Novel Protein Kinase C-Mediated Control of Orai1 Function in Invasive Melanoma. Mol. Cell Biol. 2015, 35, 2790–2798. [Google Scholar] [CrossRef]
- Stanisz, H.; Saul, S.; Müller, C.S.; Kappl, R.; Niemeyer, B.A.; Vogt, T.; Hoth, M.; Roesch, A.; Bogeski, I. Inverse Regulation of Melanoma Growth and Migration by Orai1/STIM2-Dependent Calcium Entry. Pigment. Cell Melanoma Res. 2014, 27, 442–453. [Google Scholar] [CrossRef]
- Sun, J.; Lu, F.; He, H.; Shen, J.; Messina, J.; Mathew, R.; Wang, D.; Sarnaik, A.A.; Chang, W.C.; Kim, M.; et al. STIM1- and Orai1-Mediated Ca2+ Oscillation Orchestrates Invadopodium Formation and Melanoma Invasion. J. Cell Biol. 2014, 207, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Motiani, R.K.; Hyzinski-García, M.C.; Zhang, X.; Henkel, M.M.; Abdullaev, I.F.; Kuo, Y.H.; Matrougui, K.; Mongin, A.A.; Trebak, M. STIM1 and Orai1 Mediate CRAC Channel Activity and Are Essential for Human Glioblastoma Invasion. Pflug. Arch. 2013, 465, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lkhagvadorj, S.; Lee, M.R.; Hwang, K.H.; Chung, H.C.; Jung, J.H.; Cha, S.K.; Eom, M. Orai1 and STIM1 Are Critical for Cell Migration and Proliferation of Clear Cell Renal Cell Carcinoma. Biochem. Biophys. Res. Commun. 2014, 448, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Karacicek, B.; Erac, Y.; Tosun, M. Functional Consequences of Enhanced Expression of STIM1 and Orai1 in Huh-7 Hepatocellular Carcinoma Tumor-Initiating Cells. BMC Cancer 2019, 19, 1–10. [Google Scholar] [CrossRef]
- Boutin, B.; Tajeddine, N.; Monaco, G.; Molgo, J.; Vertommen, D.; Rider, M.; Parys, J.B.; Bultynck, G.; Gailly, P. Endoplasmic Reticulum Ca2+ Content Decrease by PKA-Dependent Hyperphosphorylation of Type 1 IP3 Receptor Contributes to Prostate Cancer Cell Resistance to Androgen Deprivation. Cell Calcium. 2015, 57, 312–320. [Google Scholar] [CrossRef]
- Chen, Y.F.; Chiu, W.T.; Chen, Y.T.; Lin, P.Y.; Huang, H.J.; Chou, C.Y.; Chang, H.; Tang, M.J.; Shen, M.R. Calcium Store Sensor Stromal-Interaction Molecule 1-Dependent Signaling Plays an Important Role in Cervical Cancer Growth, Migration, and Angiogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 15225–15230. [Google Scholar] [CrossRef]
- Jardin, I.; Lopez, J.J.; Salido, G.M.; Rosado, J.A. Store-Operated Ca2+ Entry in Breast Cancer Cells: Remodeling and Functional Role. Int. J. Mol. Sci. 2018, 19, 4053. [Google Scholar] [CrossRef]
- Motiani, R.K.; Abdullaev, I.F.; Trebak, M. A Novel Native Store-Operated Calcium Channel Encoded by Orai3: Selective Requirement of Orai3 versus Orai1 in Estrogen Receptor-Positive versus Estrogen Receptor-Negative Breast Cancer Cells. J. Biol. Chem. 2010, 285, 19173–19183. [Google Scholar] [CrossRef]
- McAndrew, D.; Grice, D.M.; Peters, A.A.; Davis, F.M.; Stewart, T.; Rice, M.; Smart, C.E.; Brown, M.A.; Kenny, P.A.; Roberts-Thomson, S.J.; et al. ORAI1-Mediated Calcium Influx in Lactation and in Breast Cancer. Mol. Cancer Ther. 2011, 10, 448–460. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, J.J.; Huang, X.Y. Orai1 and STIM1 Are Critical for Breast Tumor Cell Migration and Metastasis. Cancer Cell 2009, 15, 124–134. [Google Scholar] [CrossRef]
- Chen, Y.F.; Lin, P.C.; Yeh, Y.M.; Chen, L.H.; Shen, M.R. Store-Operated Ca2+ Entry in Tumor Progression: From Molecular Mechanisms to Clinical Implications. Cancers 2019, 11, 899. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, T.A.; Lively, S.; Vincent, C.; Schlichter, L.C. Regulation of Podosome Formation, Microglial Migration and Invasion by Ca2+-Signaling Molecules Expressed in Podosomes. J. Neuroinflamm. 2012, 9, 250. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.W.; Chen, Y.F.; Chiu, W.T.; Chen, H.C.; Shen, M.R. STIM1-Dependent Ca2+ Signaling Regulates Podosome Formation to Facilitate Cancer Cell Invasion. Sci. Rep. 2017, 7, 11523. [Google Scholar] [CrossRef]
- Fujii, Y.; Shiota, M.; Ohkawa, Y.; Baba, A.; Wanibuchi, H.; Kinashi, T.; Kurosaki, T.; Baba, Y. Surf4 Modulates STIM1-Dependent Calcium Entry. Biochem. Biophys. Res. Commun. 2012, 422, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Quintana, A.; Rajanikanth, V.; Farber-Katz, S.; Gudlur, A.; Zhang, C.; Jing, J.; Zhou, Y.; Rao, A.; Hogan, P.G. TMEM110 Regulates the Maintenance and Remodeling of Mammalian ER-Plasma Membrane Junctions Competent for STIM-ORAI Signaling. Proc. Natl. Acad. Sci. USA 2015, 112, E7083–E7092. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, S.; Jung, H.J.; Kim, K.D.; Souda, P.; Whitelegge, J.; Gwack, Y. A Novel EF-Hand Protein, CRACR2A, Is a Cytosolic Ca2+ Sensor That Stabilizes CRAC Channels in T Cells. Nat. Cell Biol. 2010, 12, 436–446. [Google Scholar] [CrossRef]
- Walsh, C.M.; Doherty, M.K.; Tepikin, A.V.; Burgoyne, R.D. Evidence for an Interaction between Golli and STIM1 in Store-Operated Calcium Entry. Biochem. J. 2010, 430, 453–460. [Google Scholar] [CrossRef]
- Zeiger, W.; Ito, D.; Swetlik, C.; Oh-hora, M.; Villereal, M.L.; Thinakaran, G. Stanniocalcin 2 Is a Negative Modulator of Store-Operated Calcium Entry. Mol. Cell Biol. 2011, 31, 3710–3722. [Google Scholar] [CrossRef]
- Carreras-Sureda, A.; Cantero-Recasens, G.; Rubio-Moscardo, F.; Kiefer, K.; Peinelt, C.; Niemeyer, B.A.; Valverde, M.A.; Vicente, R. ORMDL3 Modulates Store-Operated Calcium Entry and Lymphocyte Activation. Hum. Mol. Genet. 2013, 22, 519–530. [Google Scholar] [CrossRef]
- Palty, R.; Raveh, A.; Kaminsky, I.; Meller, R.; Reuveny, E. SARAF Inactivates the Store Operated Calcium Entry Machinery to Prevent Excess Calcium Refilling. Cell 2012, 149, 425–438. [Google Scholar] [CrossRef]
- Zomot, E.; Achildiev Cohen, H.; Dagan, I.; Militsin, R.; Palty, R. Bidirectional Regulation of Calcium Release–Activated Calcium (CRAC) Channel by SARAF. J. Cell Biol. 2021, 220, e202104007. [Google Scholar] [CrossRef] [PubMed]
- Jha, A.; Ahuja, M.; Maléth, J.; Moreno, C.M.; Yuan, J.P.; Kim, M.S.; Muallem, S. The STIM1 CTID Domain Determines Access of SARAF to SOAR to Regulate Orai1 Channel Function. J. Cell Biol. 2013, 202, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Jardin, I.; Nieto-Felipe, J.; Alvarado, S.; Diez-Bello, R.; Lopez, J.J.; Salido, G.M.; Smani, T.; Rosado, J.A. SARAF and EFHB Modulate Store-Operated Ca2+ Entry and Are Required for Cell Proliferation, Migration and Viability in Breast Cancer Cells. Cancers 2021, 13, 4160. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Du, L.; Zhou, Y.; Li, M. Store-Operated CRAC Channel Inhibitors: Opportunities and Challenges. Future Med. Chem. 2016, 8, 817–832. [Google Scholar] [CrossRef] [PubMed]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular Principles of Metastasis: A Hallmark of Cancer Revisited. Signal Transduct. Target. Ther. 2020, 5, 28. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A Web Server for Cancer and Normal Gene Expression Profiling and Interactive Analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef]
- Hu, C.-D.; Chinenov, Y.; Kerppola, T.K. Visualization of Interactions among BZIP and Rel Family Proteins in Living Cells Using Bimolecular Fluorescence Complementation. Mol. Cell 2002, 9, 789–798. [Google Scholar] [CrossRef]
- Kerppola, T.K. Bimolecular Fluorescence Complementation (BiFC) Analysis as a Probe of Protein Interactions in Living Cells. Annu. Rev. Biophys. 2008, 37, 465–487. [Google Scholar] [CrossRef]
- Hertel, F.; Mo, G.C.H.; Duwé, S.; Dedecker, P.; Zhang, J. RefSOFI for Mapping Nanoscale Organization of Protein-Protein Interactions in Living Cells. Cell Rep. 2016, 14, 390–400. [Google Scholar] [CrossRef]
- Parsons, J.T.; Horwitz, A.R.; Schwartz, M.A. Cell Adhesion: Integrating Cytoskeletal Dynamics and Cellular Tension. Nat. Rev. Mol. Cell Biol. 2010, 11, 633–643. [Google Scholar] [CrossRef]
- Prevarskaya, N.; Skryma, R.; Shuba, Y. Ion Channels and the Hallmarks of Cancer. Trends Mol. Med. 2010, 16, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Collado, J.; Lopez, J.J.; Jardin, I.; Camello, P.J.; Falcon, D.; Regodon, S.; Salido, G.M.; Smani, T.; Rosado, J.A. Adenylyl Cyclase Type 8 Overexpression Impairs Phosphorylation-Dependent Orai1 Inactivation and Promotes Migration in MDA-MB-231 Breast Cancer Cells. Cancers 2019, 11, 1624. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-Y.; Zhang, Y.-Q.; Zeng, Y.-Z.; Zhu, J.-L.; Chen, H.; Wei, X.-L.; Liu, L.-J. TRPC1 Inhibits the Proliferation and Migration of Estrogen Receptor-Positive Breast Cancer and Gives a Better Prognosis by Inhibiting the PI3K/AKT Pathway. Breast Cancer Res. Treat. 2020, 182, 21–33. [Google Scholar] [CrossRef] [PubMed]
- el Hiani, Y.; Lehen’kyi, V.; Ouadid-Ahidouch, H.; Ahidouch, A. Activation of the Calcium-Sensing Receptor by High Calcium Induced Breast Cancer Cell Proliferation and TRPC1 Cation Channel over-Expression Potentially through EGFR Pathways. Arch. Biochem. Biophys. 2009, 486, 58–63. [Google Scholar] [CrossRef]
- Jardin, I.; Diez-Bello, R.; Falcon, D.; Alvarado, S.; Regodon, S.; Salido, G.M.; Smani, T.; Rosado, J.A. Melatonin Downregulates TRPC6, Impairing Store-Operated Calcium Entry in Triple-Negative Breast Cancer Cells. J. Biol. Chem. 2021, 296, 100254. [Google Scholar] [CrossRef]
- Jardin, I.; Diez-Bello, R.; Lopez, J.J.; Redondo, P.C.; Salido, G.M.; Smani, T.; Rosado, J.A. Trpc6 Channels Are Required for Proliferation, Migration and Invasion of Breast Cancer Cell Lines by Modulation of Orai1 and Orai3 Surface Exposure. Cancers 2018, 10, 331. [Google Scholar] [CrossRef]
- Yang, Y.; Jiang, Z.; Wang, B.; Chang, L.; Liu, J.; Zhang, L.; Gu, L. Expression of STIM1 Is Associated with Tumor Aggressiveness and Poor Prognosis in Breast Cancer. Pathol. Res. Pract. 2017, 213, 1043–1047. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, S.; Feng, R. STIM1 Plays an Important Role in TGF-β-Induced Suppression of Breast Cancer Cell Proliferation. Oncotarget 2016, 7, 16866–16878. [Google Scholar] [CrossRef]
- Zhang, S.; Miao, Y.; Zheng, X.; Gong, Y.; Zhang, J.; Zou, F.; Cai, C. STIM1 and STIM2 Differently Regulate Endogenous Ca2+ Entry and Promote TGF-β-Induced EMT in Breast Cancer Cells. Biochem. Biophys. Res. Commun. 2017, 488, 74–80. [Google Scholar] [CrossRef]
- Xie, J.; Pan, H.; Yao, J.; Zhou, Y.; Han, W. SOCE and Cancer: Recent Progress and New Perspectives. Int. J. Cancer 2016, 138, 2067–2077. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, N.; Pan, H.; Xie, J.; Han, W. Development of Store-Operated Calcium Entry-Targeted Compounds in Cancer. Front. Pharmacol. 2021, 12, 688244. [Google Scholar] [CrossRef] [PubMed]
- Mo, P.; Yang, S. The Store-Operated Calcium Channels in Cancer Metastasis: From Cell Migration, Invasion to Metastatic Colonization. Front. Biosci. Landmark 2018, 23, 1241–1256. [Google Scholar] [CrossRef]
- Benzerdjeb, N.; Sevestre, H.; Ahidouch, A.; Ouadid-Ahidouch, H. Orai3 Is a Predictive Marker of Metastasis and Survival in Resectable Lung Adenocarcinoma. Oncotarget 2016, 7, 81588–81597. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, R.S.; Lim, J.Y.; Turgut, A.; Servage, K.; Zhang, J.; Orth, K.; Sosale, N.G.; Lazzara, M.J.; Allegood, J.; Casanova, J.E. Calcium-Stimulated Disassembly of Focal Adhesions Mediated by an ORP3/IQSec1 Complex. eLife 2020, 9, e54113. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-S.; Lin, Y.-H.; Nguyen Thi, M.; Hsiao, S.-C.; Chiu, W.-T. STIM1 Controls the Focal Adhesion Dynamics and Cell Migration by Regulating SOCE in Osteosarcoma. Int. J. Mol. Sci. 2021, 23, 162. [Google Scholar] [CrossRef]
- Lee, S.K.; Kweon, Y.C.; Lee, A.R.; Lee, Y.Y.; Park, C.Y. Metastasis Enhancer PGRMC1 Boosts Store-Operated Ca2+ Entry by Uncoiling Ca2+ Sensor STIM1 for Focal Adhesion Turnover and Actomyosin Formation. Cell Rep. 2022, 38, 110281. [Google Scholar] [CrossRef]
- Jeon, I.S.; Kim, H.R.; Shin, E.Y.; Kim, E.G.; Han, H.S.; Hong, J.T.; Lee, H.K.; Song, K.D.; Choi, J.K. Modulation of Store-Operated Calcium Entry and Nascent Adhesion by P21-Activated Kinase 1. Exp. Mol. Med. 2018, 50, 1–10. [Google Scholar] [CrossRef]
- Rizaner, N.; Onkal, R.; Fraser, S.P.; Pristerá, A.; Okuse, K.; Djamgoz, M.B.A. Intracellular Calcium Oscillations in Strongly Metastatic Human Breast and Prostate Cancer Cells: Control by Voltage-Gated Sodium Channel Activity. Eur. Biophys. J. 2016, 45, 735–748. [Google Scholar] [CrossRef]
- Sanchez-Collado, J.; Lopez, J.J.; Cantonero, C.; Jardin, I.; Regodón, S.; Redondo, P.C.; Gordillo, J.; Smani, T.; Salido, G.M.; Rosado, J.A. Orai2 Modulates Store-Operated Ca2+ Entry and Cell Cycle Progression in Breast Cancer Cells. Cancers 2022, 14, 114. [Google Scholar] [CrossRef]
- Chakraborty, S.; Ghosh, S.; Banerjee, B.; Santra, A.; Adhikary, A.; Misra, A.K.; Sen, P.C. Phemindole, a Synthetic Di-Indole Derivative Maneuvers the Store Operated Calcium Entry (SOCE) to Induce Potent Anti-Carcinogenic Activity in Human Triple Negative Breast Cancer Cells. Front. Pharmacol. 2016, 7, 114. [Google Scholar] [CrossRef]
- Blanco, C.; Morales, D.; Mogollones, I.; Vergara-Jaque, A.; Vargas, C.; Álvarez, A.; Riquelme, D.; Leiva-Salcedo, E.; González, W.; Morales, D.; et al. EB1- and EB2-dependent anterograde trafficking of TRPM4 regulates focal adhesion turnover and cell invasion. FASEB J. 2019, 33, 9434–9452. [Google Scholar] [CrossRef] [PubMed]
- Rivas, J.; Díaz, N.; Silva, I.; Morales, D.; Lavanderos, B.; Álvarez, A.; Saldías, M.P.; Pulgar, E.; Cruz, P.; Maureira, D.; et al. KCTD5, a Novel TRPM4-Regulatory Protein Required for Cell Migration as a New Predictor for Breast Cancer Prognosis. FASEB J. 2020, 34, 7847–7865. [Google Scholar] [CrossRef] [PubMed]
- Beals, C.R.; Clipstone, N.A.; Ho, S.N.; Crabtree, G.R. Nuclear localization of NF-ATc by a calcineurin-dependent, cyclosporin-sensitive intramolecular interaction. Genes Dev. 1997, 11, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Mattheyses, A.L.; Simon, S.M.; Rappoport, J.Z. Imaging with Total Internal Reflection Fluorescence Microscopy for the Cell Biologist. J. Cell Sci. 2010, 123, 3621–3628. [Google Scholar] [CrossRef]
- Cáceres, M.; Ortiz, L.; Recabarren, T.; Romero, A.; Colombo, A.; Leiva-Salcedo, E.; Varela, D.; Rivas, J.; Silva, I.; Morales, D.; et al. TRPM4 Is a Novel Component of the Adhesome Required for Focal Adhesion Disassembly, Migration and Contractility. PLoS ONE 2015, 10, e0130540. [Google Scholar] [CrossRef]
- Goetz, J.G.; Joshi, B.; Lajoie, P.; Strugnell, S.S.; Scudamore, T.; Kojic, L.D.; Nabi, I.R. Concerted Regulation of Focal Adhesion Dynamics by Galectin-3 and Tyrosine-Phosphorylated Caveolin-1. J. Cell Biol. 2008, 180, 1261–1275. [Google Scholar] [CrossRef]
- Ghosh, A.; Sarkar, S.; Banerjee, S.; Behbod, F.; Tawfik, O.; McGregor, D.; Graff, S.; Banerjee, S.K. MIND Model for Triple-Negative Breast Cancer in Syngeneic Mice for Quick and Sequential Progression Analysis of Lung Metastasis. PLoS ONE 2018, 13, e0198143. [Google Scholar] [CrossRef]
- Sápi, J.; Kovács, L.; Drexler, D.A.; Kocsis, P.; Gajári, D.; Sápi, Z. Tumor Volume Estimation and Quasi-Continuous Administration for Most Effective Bevacizumab Therapy. PLoS ONE 2015, 10, e0142190. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid | Source | Catalog # | Amino Acid Sequence Information |
|---|---|---|---|
| SARAF-myc-DDK | Origene | RC201864 | - |
| mScarlet-H | Addgene | 85043 | - |
| mScarlet-C-SARAF | Cerda Lab | - | cytosolic domain SARAF (195–339 aa) fused to mScarlet protein |
| EGFP-Paxillin | Dr. Christopher Turner | ||
| EGFPC1-huNFATc1EE-WT | Addgene | 24219 | - |
| pcDNA3-Venus-173-N-Orai1 | Addgene | 87618 | - |
| pcDNA3.1-STIM1-Venus-173-C | Addgene | 87619 | - |
| Antibody | Isotype | Dilution | Final [], µg/mL | Source | Type | Catalog | RRID | Purification |
|---|---|---|---|---|---|---|---|---|
| Mouse anti-Orai1 | IgG1 | 1:1000 (IB) | 1 | Santa Cruz Biotechnology | mAb | sc-377281 | AB_2797381 | - |
| Mouse Anti-STIM1 | IgG2a | 1:1000 (IB) | Origene | mAb | AM20946PU-N | AB_10849514 | - | |
| Mouse anti-α-tubulin | IgG1 | 1:5000 (IB) | 1 | Sigma Aldrich | mAb | T5168 | AB_477579 | Asc |
| Mouse Anti-RFP [6G6] | IgG2c | 1:2000 (IB) 1:100 (IP) | ChromoTek | mAb | 6g6 | AB_2631395 | AP | |
| Rat Anti-RFP [5F8] | IgG2a | 1:100 (IP) | ChromoTek | mAb | 5f8 | AB_2336064 | AP | |
| Rabbit Anti-STIM | IgG | 1:2000 (IB) | Cell Signaling | mAb | D88E10 | AB_10828699 | ||
| Rabbit Anti-SARAF (TMEM66) | IgG | 1:250 (IB) | – | Origene | pAb | TA341997 | – | AP |
| Goat anti-Rabbit HRP-conjugated Antibody | IgG | 1:5000 (IB) | – | R&D systems | pAb | HAF008 | AB_357235 | AP |
| Goat anti-mouse HRP-conjugated Antibody | IgG | 1:5000 (IB) | – | R&D systems | pAb | HAF007 | AB_357234 | AP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saldías, M.P.; Cruz, P.; Silva, I.; Orellana-Serradell, O.; Lavanderos, B.; Maureira, D.; Pinto, R.; Cerda, O. The Cytoplasmic Region of SARAF Reduces Triple-Negative Breast Cancer Metastasis through the Regulation of Store-Operated Calcium Entry. Int. J. Mol. Sci. 2023, 24, 5306. https://doi.org/10.3390/ijms24065306
Saldías MP, Cruz P, Silva I, Orellana-Serradell O, Lavanderos B, Maureira D, Pinto R, Cerda O. The Cytoplasmic Region of SARAF Reduces Triple-Negative Breast Cancer Metastasis through the Regulation of Store-Operated Calcium Entry. International Journal of Molecular Sciences. 2023; 24(6):5306. https://doi.org/10.3390/ijms24065306
Chicago/Turabian StyleSaldías, María Paz, Pablo Cruz, Ian Silva, Octavio Orellana-Serradell, Boris Lavanderos, Diego Maureira, Raquel Pinto, and Oscar Cerda. 2023. "The Cytoplasmic Region of SARAF Reduces Triple-Negative Breast Cancer Metastasis through the Regulation of Store-Operated Calcium Entry" International Journal of Molecular Sciences 24, no. 6: 5306. https://doi.org/10.3390/ijms24065306
APA StyleSaldías, M. P., Cruz, P., Silva, I., Orellana-Serradell, O., Lavanderos, B., Maureira, D., Pinto, R., & Cerda, O. (2023). The Cytoplasmic Region of SARAF Reduces Triple-Negative Breast Cancer Metastasis through the Regulation of Store-Operated Calcium Entry. International Journal of Molecular Sciences, 24(6), 5306. https://doi.org/10.3390/ijms24065306