
Uridine Alleviates Sepsis-Induced Acute Lung Injury by Inhibiting Ferroptosis of Macrophage


Abstract
1. Introduction
2. Results
2.1. Uridine Metabolism Homeostasis Was Disturbed in Sepsis-Induced ALI
2.2. Uridine Supplementation Alleviated Sepsis-Induced ALI and Inflammation of Lung Tissue
2.3. Uridine Inhibited Ferroptosis in Lung Tissue Dependent on Nrf2 Activation and ACSL4 Inhibition
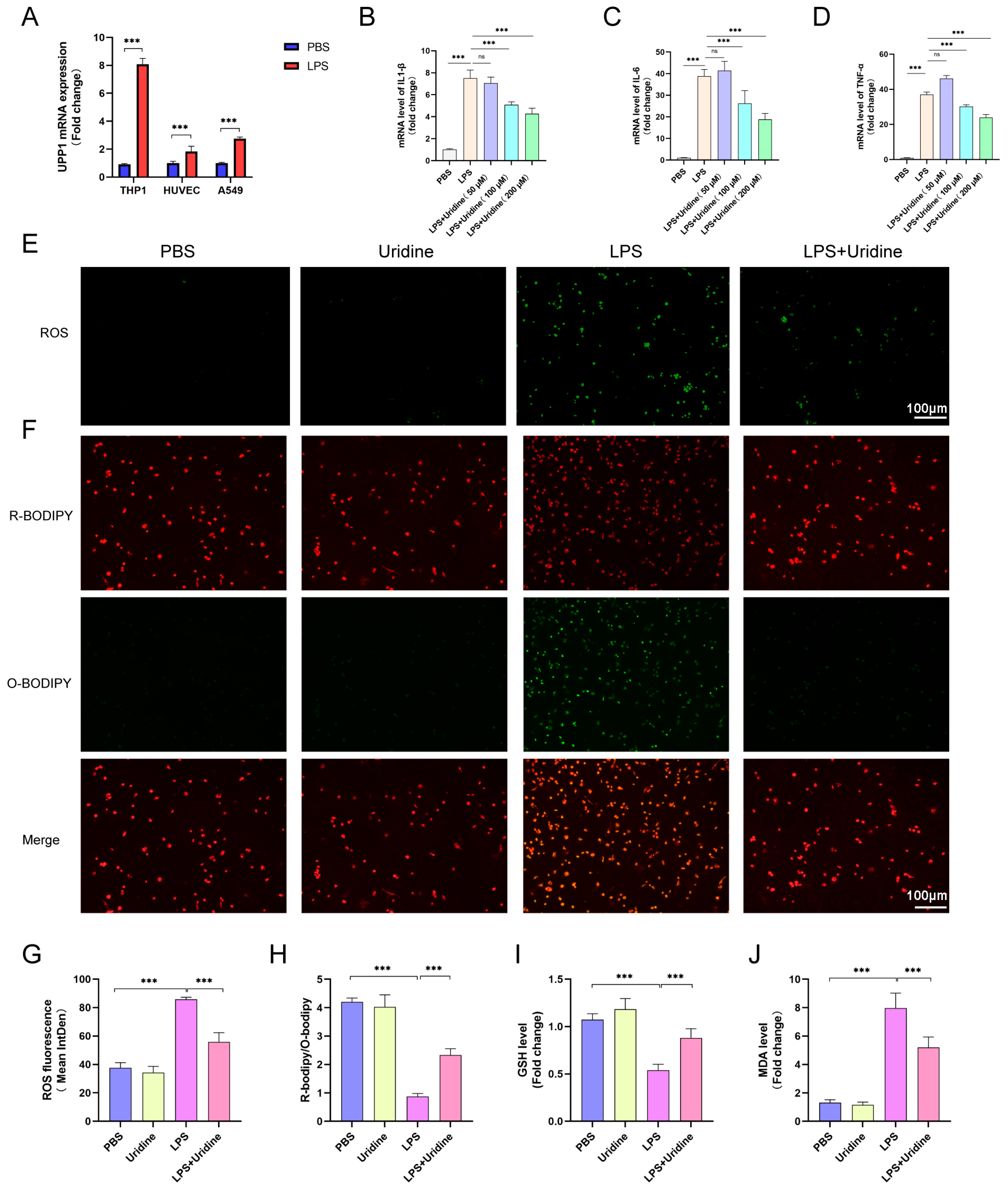
2.4. Uridine Supplementation Restrained Inflammation and Oxidative Stress in THP-1 Cells
2.5. Uridine Suppressed Ferroptosis in THP-1 Cells via Activating Nrf2 and Inhibiting ACSL4 Expression
2.6. Nrf2 Knockdown Abrogated the Ferroptosis Resistance Effect of Uridine
2.7. Dysregulated Uridine Metabolism Was Confirmed in Patients with Sepsis
3. Discussion
4. Methods and Materials
4.1. Chemical and Reagents
4.2. Establishment of Sepsis-Induced Acute Lung Injury Model
4.3. BALF Collection, Processing and Determination of Protein Concentration
4.4. Lung Wet-to-Dry Weight Ratio Measurement
4.5. H&E Staining and Lung Injury Score Evaluation
4.6. Assay of MPO Activity
4.7. Quantification of Cytokines, Detection of Tissue Iron and Lipid Peroxidation Level
4.8. Cell Culture and Transfection
4.9. Macrophage Differentiation and Treatment
4.10. GSH Measurement
4.11. Cell Viability Assay
4.12. RNA Extraction and Real-Time Polymerase Chain Reaction
4.13. Western Blotting
4.14. ROS, Lipid Peroxidation and Mitochondrial Potential Measurement
4.15. Dataset Collection and Preprocessing
4.16. Bioinformatic Analysis and Visualization
4.17. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butt, Y.; Kurdowska, A.; Allen, T.C. Acute Lung Injury: A Clinical and Molecular Review. Arch. Pathol. Lab. Med. 2016, 140, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.; Rubenfeld, G.D. Fifty Years of Research in ARDS. The Epidemiology of Acute Respiratory Distress Syndrome. A 50th Birthday Review. Am. J. Respir. Crit. Care Med. 2017, 195, 860–870. [Google Scholar] [CrossRef] [PubMed]
- De Freitas Caires, N.; Gaudet, A.; Portier, L.; Tsicopoulos, A.; Mathieu, D.; Lassalle, P. Endocan, sepsis, pneumonia, and acute respiratory distress syndrome. Crit. Care 2018, 22, 280. [Google Scholar] [CrossRef] [PubMed]
- Sadowitz, B.; Roy, S.; Gatto, L.A.; Habashi, N.; Nieman, G. Lung injury induced by sepsis: Lessons learned from large animal models and future directions for treatment. Expert Rev. Anti Infect. Ther. 2011, 9, 1169–1178. [Google Scholar] [CrossRef]
- Kumar, V. Pulmonary Innate Immune Response Determines the Outcome of Inflammation During Pneumonia and Sepsis-Associated Acute Lung Injury. Front. Immunol. 2020, 11, 1722. [Google Scholar] [CrossRef]
- Wick, K.D.; McAuley, D.F.; Levitt, J.E.; Beitler, J.R.; Annane, D.; Riviello, E.D.; Calfee, C.S.; Matthay, M.A. Promises and challenges of personalized medicine to guide ARDS therapy. Crit. Care 2021, 25, 404. [Google Scholar] [CrossRef]
- Patel, V.J.; Biswas Roy, S.; Mehta, H.J.; Joo, M.; Sadikot, R.T. Alternative and Natural Therapies for Acute Lung Injury and Acute Respiratory Distress Syndrome. Biomed. Res. Int. 2018, 2018, 2476824. [Google Scholar] [CrossRef]
- Connolly, G.P.; Duley, J.A. Uridine and its nucleotides: Biological actions, therapeutic potentials. Trends. Pharmacol. Sci. 1999, 20, 218–225. [Google Scholar] [CrossRef]
- Anderson, C.M.; Parkinson, F.E. Potential signalling roles for UTP and UDP: Sources, regulation and release of uracil nucleotides. Trends Pharmacol. Sci. 1997, 18, 387–392. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, S.; Xie, C.; Fang, J. Uridine Metabolism and Its Role in Glucose, Lipid, and Amino Acid Homeostasis. Biomed. Res. Int. 2020, 2020, 7091718. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Koyama, H.; Kurajoh, M.; Shoji, T.; Tsutsumi, Z.; Moriwaki, Y. Biochemistry of uridine in plasma. Clin. Chim. Acta 2011, 412, 1712–1724. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Marra, R.; Andolfo, I.; Manna, F.; De Rosa, G.; Rosato, B.E.; Radhakrishnan, K.; Fahey, M.; Iolascon, A. Uridine treatment normalizes the congenital dyserythropoietic anemia type II-like hematological phenotype in a patient with homozygous mutation in the CAD gene. Am. J. Hematol. 2020, 95, 1423–1426. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, H.; Gong, J.; Yu, H.; Wu, D.; Hou, J.; Li, M.; Sun, X. Aging affects the biological activity of fibroblast growth factor (FGF) in gastric epithelial cell, which is partially rescued by uridine. Bioengineered 2022, 13, 3724–3738. [Google Scholar] [CrossRef] [PubMed]
- Chenna Narendra, S.; Chalise, J.P.; Magnusson, M.; Uppugunduri, S. Local but Not Systemic Administration of Uridine Prevents Development of Antigen-Induced Arthritis. PLoS ONE 2015, 10, e0141863. [Google Scholar] [CrossRef]
- Jeengar, M.K.; Thummuri, D.; Magnusson, M.; Naidu, V.G.M.; Uppugunduri, S. Uridine Ameliorates Dextran Sulfate Sodium (DSS)-Induced Colitis in Mice. Sci. Rep. 2017, 7, 3924. [Google Scholar] [CrossRef]
- Cicko, S.; Grimm, M.; Ayata, K.; Beckert, J.; Meyer, A.; Hossfeld, M.; Zissel, G.; Idzko, M.; Müller, T. Uridine supplementation exerts anti-inflammatory and anti-fibrotic effects in an animal model of pulmonary fibrosis. Respir. Res. 2015, 16, 105. [Google Scholar] [CrossRef]
- Jiang, N.; Zhao, Z. Intestinal aging is alleviated by uridine via regulating inflammation and oxidative stress in vivo and in vitro. Cell Cycle 2022, 21, 1519–1531. [Google Scholar] [CrossRef]
- Adant, I.; Bird, M.; Decru, B.; Windmolders, P.; Wallays, M.; de Witte, P.; Rymen, D.; Witters, P.; Vermeersch, P.; Cassiman, D.; et al. Pyruvate and uridine rescue the metabolic profile of OXPHOS dysfunction. Mol. Metab. 2022, 63, 101537. [Google Scholar] [CrossRef]
- Le, T.T.; Ziemba, A.; Urasaki, Y.; Hayes, E.; Brotman, S.; Pizzorno, G. Disruption of uridine homeostasis links liver pyrimidine metabolism to lipid accumulation. J. Lipid Res. 2013, 54, 1044–1057. [Google Scholar] [CrossRef]
- Ye, J.; Jin, Z.; Chen, S.; Guo, W. Uridine relieves MSCs and chondrocyte senescence in vitvo and exhibits the potential to treat osteoarthritis in vivo. Cell Cycle 2022, 21, 33–48. [Google Scholar] [CrossRef]
- Krylova, I.B.; Selina, E.N.; Bulion, V.V.; Rodionova, O.M.; Evdokimova, N.R.; Belosludtseva, N.V.; Shigaeva, M.I.; Mironova, G.D. Uridine treatment prevents myocardial injury in rat models of acute ischemia and ischemia/reperfusion by activating the mitochondrial ATP-dependent potassium channel. Sci. Rep. 2021, 11, 16999. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, W.; Geng, L.; Sun, L.; Wang, Q.; Yu, Y.; Yan, P.; Liang, C.; Ren, J.; Song, M.; et al. Cross-species metabolomic analysis identifies uridine as a potent regeneration promoting factor. Cell Discov. 2022, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Mironova, G.D.; Khrenov, M.O.; Yu, T.E.; Glushkova, O.V.; Parfenyuk, S.B.; Novoselova, T.V.; Lunin, S.M.; Belosludtseva, N.V.; Novoselova, E.G.; Lemasters, J.J. The role of mitochondrial KATP channel in anti-inflammatory effects of uridine in endotoxemic mice. Arch. Biochem. Biophys. 2018, 654, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, J.; Xie, W. The role of ferroptosis in acute lung injury. Mol. Cell. Biochem. 2022, 477, 1453–1461. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, X.; Liu, X.; Shen, L.; Chen, Q.; Shu, Q. Targeting Ferroptosis as a Promising Therapeutic Strategy for Ischemia-Reperfusion Injury. Antioxidants 2022, 11, 2196. [Google Scholar] [CrossRef]
- Mou, Y.; Wang, J.; Wu, J.; He, D.; Zhang, C.; Duan, C.; Li, B. Ferroptosis, a new form of cell death: Opportunities and challenges in cancer. J. Hematol. Oncol. 2019, 12, 34. [Google Scholar] [CrossRef]
- Li, S.; Wang, R.; Wang, Y.; Liu, Y.; Qiao, Y.; Li, P.; Chen, J.; Pan, S.; Feng, Q.; Liu, Z.; et al. Ferroptosis: A new insight for treatment of acute kidney injury. Front. Pharmacol. 2022, 13, 1065867. [Google Scholar] [CrossRef]
- Dodson, M.; Castro-Portuguez, R.; Zhang, D.D. NRF2 plays a critical role in mitigating lipid peroxidation and ferroptosis. Redox Biol. 2019, 23, 101107. [Google Scholar] [CrossRef]
- Mao, C.; Liu, X.; Zhang, Y.; Lei, G.; Yan, Y.; Lee, H.; Koppula, P.; Wu, S.; Zhuang, L.; Fang, B.; et al. DHODH-mediated ferroptosis defence is a targetable vulnerability in cancer. Nature 2021, 593, 586–590. [Google Scholar] [CrossRef]
- Stockwell, B.R. Ferroptosis turns 10: Emerging mechanisms, physiological functions, and therapeutic applications. Cell 2022, 185, 2401–2421. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.D.; Badovinac, V.P.; Griffith, T.S. CD4 T Cell Responses and the Sepsis-Induced Immunoparalysis State. Front. Immunol. 2020, 11, 1364. [Google Scholar] [CrossRef] [PubMed]
- Jensen, I.J.; Sjaastad, F.V.; Griffith, T.S.; Badovinac, V.P. Sepsis-Induced T Cell Immunoparalysis: The Ins and Outs of Impaired T Cell Immunity. J. Immunol. 2018, 200, 1543–1553. [Google Scholar] [CrossRef]
- Li, R.; Li, X.; Zhao, J.; Meng, F.; Yao, C.; Bao, E.; Sun, N.; Chen, X.; Cheng, W.; Hua, H.; et al. Mitochondrial STAT3 exacerbates LPS-induced sepsis by driving CPT1a-mediated fatty acid oxidation. Theranostics 2022, 12, 976–998. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Xie, M.; Yang, M.; Yu, Y.; Zhu, S.; Hou, W.; Kang, R.; Lotze, M.T.; Billiar, T.R.; Wang, H.; et al. PKM2 regulates the Warburg effect and promotes HMGB1 release in sepsis. Nat. Commun. 2014, 5, 4436. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, B.; Zhang, D.; Wang, Z.; Zhao, Y.; Yu, Q. Uridine alleviates LPS-induced ARDS and improves insulin sensitivity by decreasing oxidative stress and inflammatory processes. Physiol. Int. 2022, 109, 215–229. [Google Scholar] [CrossRef]
- Liu, Q.; Zhai, L.; Han, M.; Shi, D.; Sun, Z.; Peng, S.; Wang, M.; Zhang, C.; Gao, J.; Yan, W.; et al. SH2 Domain-Containing Phosphatase 2 Inhibition Attenuates Osteoarthritis by Maintaining Homeostasis of Cartilage Metabolism via the Docking Protein 1/Uridine Phosphorylase 1/Uridine Cascade. Arthritis Rheumatol. 2022, 74, 462–474. [Google Scholar] [CrossRef]
- Zheng, W.V.; Li, Y.; Cheng, X.; Xu, Y.; Zhou, T.; Li, D.; Xiong, Y.; Wang, S.; Chen, Z. Uridine alleviates carbon tetrachloride-induced liver fibrosis by regulating the activity of liver-related cells. J. Cell. Mol. Med. 2022, 26, 840–854. [Google Scholar] [CrossRef]
- Gonçalves da Silva, E.F.; Costa, B.P.; Nassr, M.T.; de Souza Basso, B.; Bastos, M.S.; Antunes, G.L.; Reghelin, C.K.; Rosa Garcia, M.C.; Schneider Levorse, V.G.; Carlessi, L.P.; et al. Therapeutic effect of uridine phosphorylase 1 (UPP1) inhibitor on liver fibrosis in vitro and in vivo. Eur. J. Pharmacol. 2021, 890, 173670. [Google Scholar] [CrossRef]
- Zhang, J.; Zheng, Y.; Wang, Y.; Wang, J.; Sang, A.; Song, X.; Li, X. YAP1 alleviates sepsis-induced acute lung injury via inhibiting ferritinophagy-mediated ferroptosis. Front. Immunol. 2022, 13, 884362. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, D.; Xie, H.; Jia, M.; Sun, X.; Peng, F.; Guo, F.; Tang, D. AUF1 protects against ferroptosis to alleviate sepsis-induced acute lung injury by regulating NRF2 and ATF3. Cell. Mol. Life Sci. 2022, 79, 228. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Liu, B.; Xiong, R.; Geng, B.; Meng, H.; Lin, W.; Hao, B.; Zhang, L.; Wang, W.; Jiang, W.; et al. Itaconate inhibits ferroptosis of macrophage via Nrf2 pathways against sepsis-induced acute lung injury. Cell Death Discov. 2022, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Krylova, I.B.; Bulion, V.V.; Selina, E.N.; Mironova, G.D.; Sapronov, N.S. Effect of uridine on energy metabolism, LPO, and antioxidant system in the myocardium under conditions of acute coronary insufficiency. Bull. Exp. Biol. Med. 2012, 153, 644–646. [Google Scholar] [CrossRef]
- Li, Y.; Feng, D.; Wang, Z.; Zhao, Y.; Sun, R.; Tian, D.; Liu, D.; Zhang, F.; Ning, S.; Yao, J.; et al. Ischemia-induced ACSL4 activation contributes to ferroptosis-mediated tissue injury in intestinal ischemia/reperfusion. Cell Death Differ. 2019, 26, 2284–2299. [Google Scholar] [CrossRef] [PubMed]
- Doll, S.; Proneth, B.; Tyurina, Y.Y.; Panzilius, E.; Kobayashi, S.; Ingold, I.; Irmler, M.; Beckers, J.; Aichler, M.; Walch, A.; et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat. Chem. Biol. 2017, 13, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xie, C.; Zhai, Z.; Deng, Z.Y.; De Jonge, H.R.; Wu, X.; Ruan, Z. Uridine attenuates obesity, ameliorates hepatic lipid accumulation and modifies the gut microbiota composition in mice fed with a high-fat diet. Food Funct. 2021, 12, 1829–1840. [Google Scholar] [CrossRef]
- Luo, Y.; Chen, H.; Huang, R.; Wu, Q.; Li, Y.; He, Y. Guanosine and uridine alleviate airway inflammation via inhibition of the MAPK and NF-κB signals in OVA-induced asthmatic mice. Pulm. Pharmacol. Ther. 2021, 69, 102049. [Google Scholar] [CrossRef]
- Park, I.; Kim, M.; Choe, K.; Song, E.; Seo, H.; Hwang, Y.; Ahn, J.; Lee, S.H.; Lee, J.H.; Jo, Y.H.; et al. Neutrophils disturb pulmonary microcirculation in sepsis-induced acute lung injury. Eur. Respir. J. 2019, 53, 1800786. [Google Scholar] [CrossRef]
- Li, Y.; Cao, Y.; Xiao, J.; Shang, J.; Tan, Q.; Ping, F.; Huang, W.; Wu, F.; Zhang, H.; Zhang, X. Inhibitor of apoptosis-stimulating protein of p53 inhibits ferroptosis and alleviates intestinal ischemia/reperfusion-induced acute lung injury. Cell Death Differ. 2020, 27, 2635–2650. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene | Forward Primer | Reverse Primer |
|---|---|---|---|
| Mice | TNF-α | ACTGAACTTCGGGGTGATCGGT | TGGTTTGCTACGACGTGGGCTA |
| Mice | IL-6 | CCCCAATTTCCAATGCTCTCC | CGCACTAGGTTTGCCGAGTA |
| Mice | IL-1β | AATGAAGGAACGGAGGAGCC | CTCCAGCCAAGCTTCCTTGT |
| Mice | GPX4 | GCTGGGAAATGCCATCAAAT | TCCTTCTCTATCACCTGGGGCT |
| Mice | SLC7A11 | GAAATATCAGGTCATTGGTGGAGA | ATGCTCCTGCTTGAGTATGTCG |
| Mice | HO1 | GCCAGCAACAAAGTGCAAGA | TAAGGACCCATCGGAGAAGC |
| Mice | UPP1 | ATCCCAACATCTGTGCAGGC | ACCTGGCATGGTACAGCATC |
| Mice | β-actin | GTCCACCGCAAATGCTTCTA | TGCTGTCACCTTCACCGTTC |
| Human | TNF-α | TCCAGGCGGTGCTTGTTC | GCTTGTCACTCGGGGTTC |
| Human | IL-6 | ATGAGGAGACTTGCCTGGTGAA | CTCTGGCTTGTTCCTCACTACTCTC |
| Human | IL-1β | CGGGACTCACAGCAAAAAA | TTCAACACGCAGGACAGGT |
| Human | GPX4 | TGAAGATCCAACCCAAGGGC | GACGGTGTCCAAACTTGGTG |
| Human | SLC7A11 | GGCAGTTGCTGGGCTGATTTA | GATGACGAAGCCAATCCCTGT |
| Human | HO1 | ATGCCCCAGGATTTGTCAGAG | GGAAGTAGACAGGGGCGAAGAC |
| Human | UPP1 | CACCACTAGCAGACACAATTTCC | AATGCCCATACCATGACTGACAG |
| Human | β-actin | CACCCAGCACAATGAAGATCAAGAT | CCAGTTTTTAAATCCTGAGTCAAGC |
| Dataset Number | Platform | LPS Group/Sepsis | Control Group/Healthy | Annotation | Sample Type |
|---|---|---|---|---|---|
| GSE130936 | GPL339 | 6 | 4 | Affymetrix Mouse Expression 430A Array | Murine lung |
| GSE15379 | GPL1261 | 3 | 3 | Affymetrix Mouse Genome 430 2.0 Array | Murine lung |
| GSE2411 | GPL339 | 6 | 6 | Affymetrix Mouse Expression 430A Array | Murine lung |
| GSE1871 | GPL1261 | 3 | 3 | Affymetrix Mouse Genome 430 2.0 Array | Murine lung |
| GSE211210 | GPL24676 | 5 | 5 | Illumina NovaSeq 6000 (Homo sapiens) | Human Blood |
| GSE137340 | GPL10558 | 45 | 12 | Illumina HumanHT-12 V4.0 expression beadchip | Human Blood |
| GSE185263 | GPL16791 | 348 | 44 | Illumina HiSeq 2500 (Homo sapiens) | Human Blood |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, K.; Song, C.; Gao, M.; Deng, Y.; Lu, Z.; Li, N.; Geng, Q. Uridine Alleviates Sepsis-Induced Acute Lung Injury by Inhibiting Ferroptosis of Macrophage. Int. J. Mol. Sci. 2023, 24, 5093. https://doi.org/10.3390/ijms24065093
Lai K, Song C, Gao M, Deng Y, Lu Z, Li N, Geng Q. Uridine Alleviates Sepsis-Induced Acute Lung Injury by Inhibiting Ferroptosis of Macrophage. International Journal of Molecular Sciences. 2023; 24(6):5093. https://doi.org/10.3390/ijms24065093
Chicago/Turabian StyleLai, Kai, Congkuan Song, Minglang Gao, Yu Deng, Zilong Lu, Ning Li, and Qing Geng. 2023. "Uridine Alleviates Sepsis-Induced Acute Lung Injury by Inhibiting Ferroptosis of Macrophage" International Journal of Molecular Sciences 24, no. 6: 5093. https://doi.org/10.3390/ijms24065093
APA StyleLai, K., Song, C., Gao, M., Deng, Y., Lu, Z., Li, N., & Geng, Q. (2023). Uridine Alleviates Sepsis-Induced Acute Lung Injury by Inhibiting Ferroptosis of Macrophage. International Journal of Molecular Sciences, 24(6), 5093. https://doi.org/10.3390/ijms24065093