

Ovariectomy Exacerbates Acute Ethanol-Induced Tachycardia: Role of Nitric Oxide and NMDA Receptors in the Rostral Ventrolateral Medulla



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
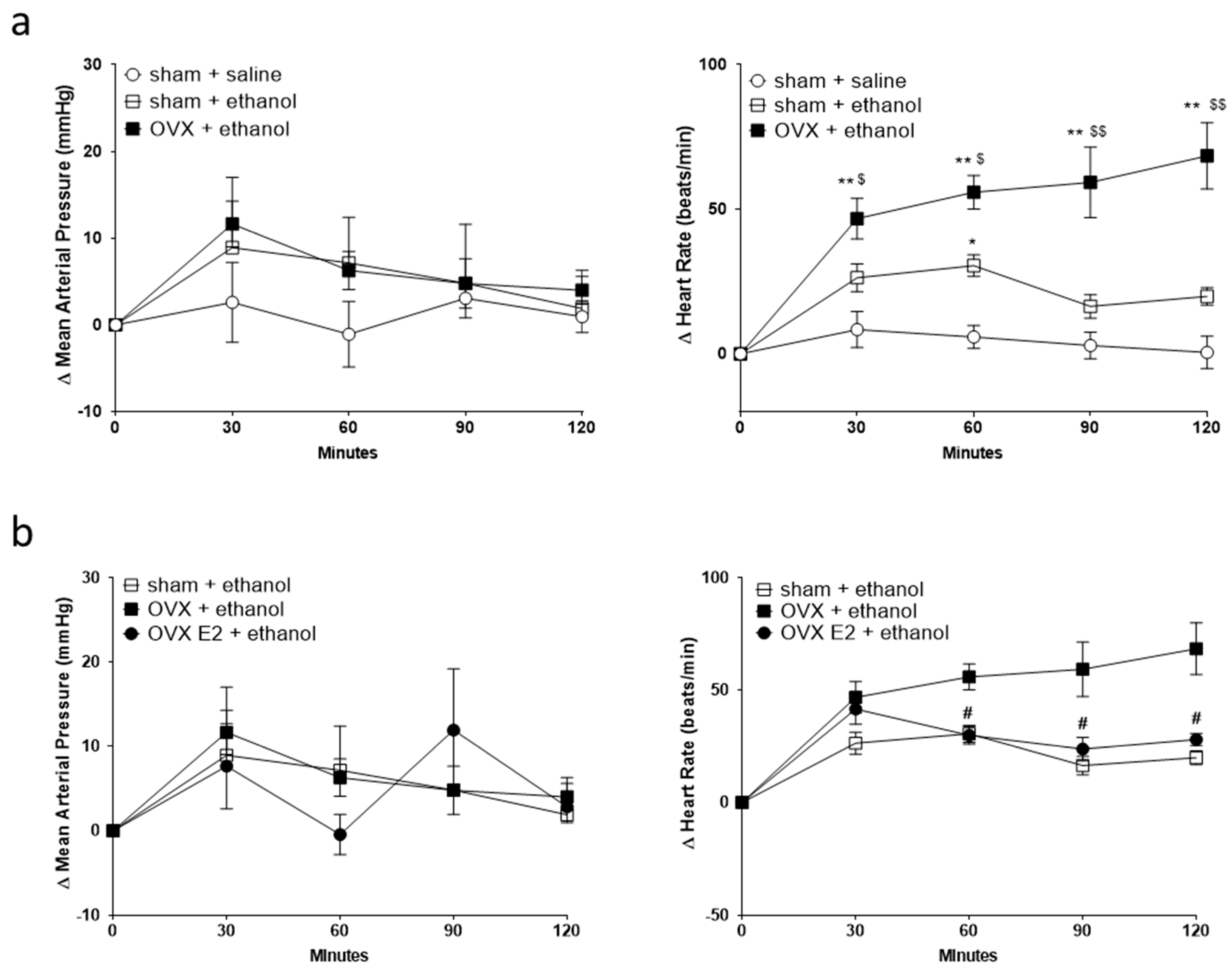
2.1. Ovariectomy Increased Ethanol-Induced Tachycardia, and Estradiol Treatment Attenuated the Tachycardia
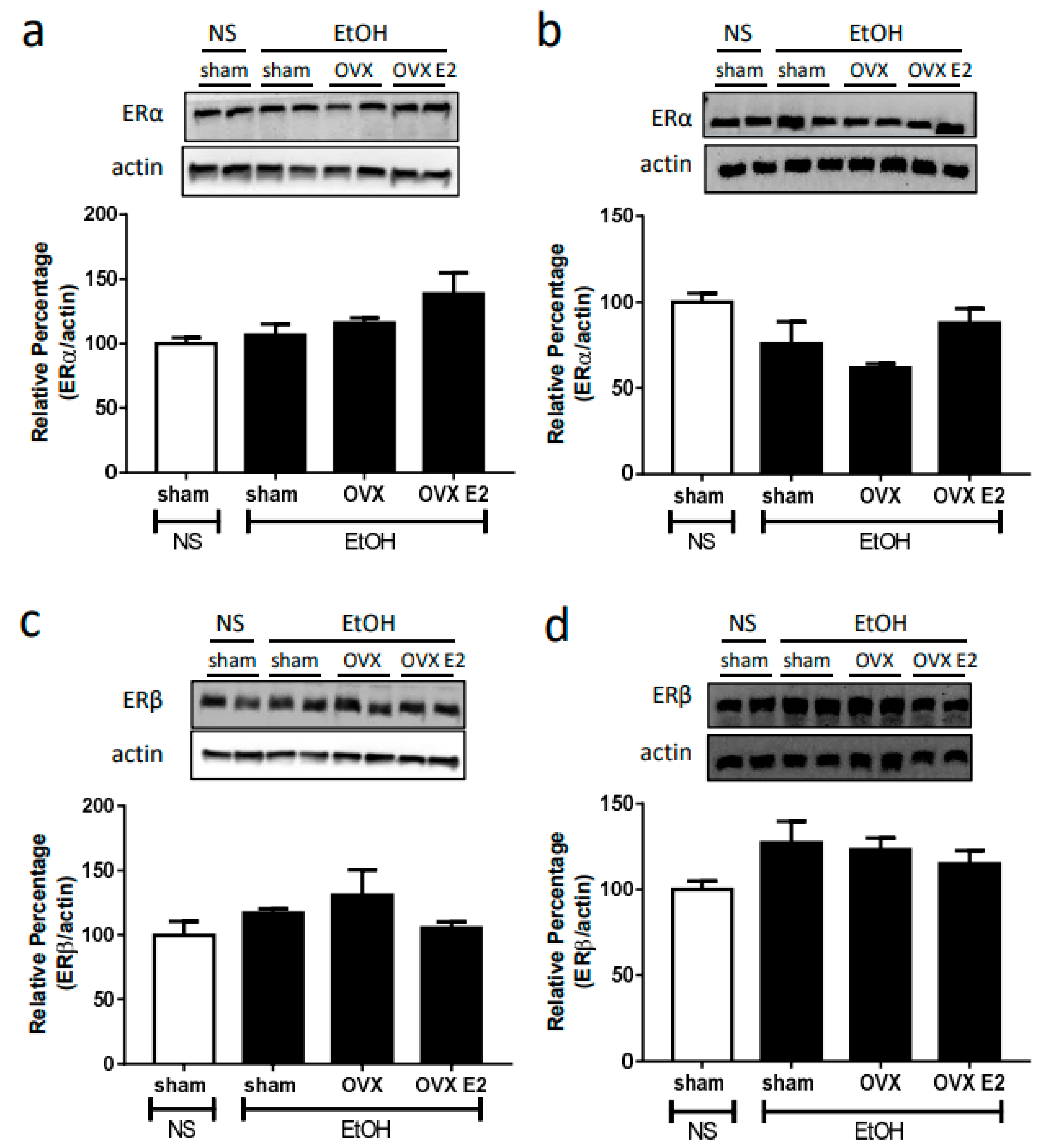
2.2. Ovariectomy and E2 Treatment Did Not Alter NOS and ER Expression in RVLM and NA
2.3. Ovariectomy Decreased NO Levels in the RVLM
2.4. Changes in the Immunoreactivity of NMDA Receptor Subunit Phosphorylation in the RVLM
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemicals
4.3. Ovariectomy (OVX)
4.4. Ethanol or Saline Administration
4.5. Blood Pressure and Heart Rate Measurement
4.6. Western Blot Analysis
4.7. Immunohistochemistry
4.8. Determination of Total Nitrate-Nitrite
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pareek, M.; Olsen, M.H. Alcohol and blood pressure. Lancet Public Health 2017, 2, e63–e64. [Google Scholar] [CrossRef]
- Ristuccia, R.C.; Spear, L.P. Adolescent and adult heart rate responses to self-administered ethanol. Alcohol. Clin. Exp. Res. 2008, 32, 1807–1815. [Google Scholar] [CrossRef] [PubMed]
- Brunner, S.; Herbel, R.; Drobesch, C.; Peters, A.; Massberg, S.; Kaab, S.; Sinner, M.F. Alcohol consumption, sinus tachycardia, and cardiac arrhythmias at the Munich Octoberfest: Results from the Munich Beer Related Electrocardiogram Workup Study (MunichBREW). Eur. Heart J. 2017, 38, 2100–2106. [Google Scholar] [CrossRef]
- Da Silva, J.S.; Montagnoli, T.L.; de Sá, M.P.L.; Zapata-Sudo, G. Heart Failure in Menopause: Treatment and New Approaches. Int. J. Mol. Sci. 2022, 23, 15140. [Google Scholar] [CrossRef] [PubMed]
- Babic, G.M.; Markovic, S.D.; Varjacic, M.; Djordjevic, N.Z.; Nikolic, T.; Stojic, I.; Jakovljevic, V. Estradiol decreases blood pressure in association with redox regulation in preeclampsia. Clin. Exp. Hypertens. 2018, 40, 281–286. [Google Scholar] [CrossRef]
- El-Mas, M.M.; Abdel-Rahman, A.A. Estrogen enhances baroreflex control of heart rate in conscious ovariectomized rats. Can. J. Physiol. Pharmacol. 1998, 76, 381–386. [Google Scholar] [CrossRef]
- Pinkham, M.I.; Barrett, C.J. Estradiol alters the chemosensitive cardiac afferent reflex in female rats by augmenting sympathoinhibition and attenuating sympathoexcitation. Clin. Exp. Pharmacol. Physiol. 2015, 42, 622–631. [Google Scholar] [CrossRef]
- Varga, K.; Kunos, G. Ethanol inhibition of baroreflex bradycardia: Role of brainstem GABA receptors. Br. J. Pharmacol. 1990, 101, 773–775. [Google Scholar] [CrossRef]
- Mao, L.; Abdel-Rahman, A.A. Blockade of L-glutamate receptors in the rostral ventrolateral medulla contributes to ethanol-evoked impairment of baroreflexes in conscious rats. Brain Res. Bull. 1995, 37, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.; Lin, H.H.; Chen, J.K.; Situmorang, J.H.; Lai, C.C. Involvement of NMDA Receptors, Nitric Oxide, and GABA in Rostral Ventrolateral Medulla in Acute Ethanol-Induced Cardiovascular Responses in Rats. Alcohol. Clin. Exp. Res. 2018, 42, 1418–1430. [Google Scholar] [CrossRef]
- Lin, H.H.; Cheng, T.T.; Lo, H.; Lin, Y.C.; Lai, C.-C. Spontaneously hypertensive rats exhibit higher sensitivity to ethanol-induced hypotensive effects: Role of NMDA receptors and nitric oxide in rostral ventrolateral medulla. Alcohol 2018, 73, 25–35. [Google Scholar] [CrossRef]
- Situmorang, J.H.; Lin, H.H.; Lo, H.; Lai, C.C. Role of neuronal nitric oxide synthase (nNOS) at medulla in tachycardia induced by repeated administration of ethanol in conscious rats. J. Biomed. Sci. 2018, 25, 1–11. [Google Scholar] [CrossRef]
- El-Mas, M.M.; Abdel-Rahman, A.A. Estrogen-dependent hypotensive effects of ethanol in conscious female rats. Alcohol. Clin. Exp. Res. 1999, 23, 624–632. [Google Scholar] [CrossRef]
- Li, G.; Abdel-Rahman, A.A. Estrogen-dependent enhancement of NO production in the nucleus tractus solitarius contributes to ethanol-induced hypotension in conscious female rats. Alcohol. Clin. Exp. Res. 2009, 33, 366–374. [Google Scholar] [CrossRef]
- Yao, F.; Abdel-Rahman, A.A. Estrogen Receptors alpha and beta Play Major Roles in Ethanol-Evoked Myocardial Oxidative Stress and Dysfunction in Conscious Ovariectomized Rats. Alcohol. Clin. Exp. Res. 2017, 41, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Doucet, M.V.; Harkin, A.; Dev, K.K. The PSD-95/nNOS complex: New drugs for depression? Pharmacol. Ther. 2012, 133, 218–229. [Google Scholar] [CrossRef]
- Wang, J.Q.; Guo, M.L.; Jin, D.Z.; Xue, B.; Fibuch, E.E.; Mao, L.M. Roles of subunit phosphorylation in regulating glutamate receptor function. Eur. J. Pharmacol. 2014, 728, 183–187. [Google Scholar] [CrossRef] [PubMed]
- McGregor, C.; Riordan, A.; Thornton, J. Estrogens and the cognitive symptoms of schizophrenia: Possible neuroprotective mechanisms. Front. Neuroendocrinol. 2017, 47, 19–33. [Google Scholar] [CrossRef]
- Liu, S.B.; Zhao, M.G. Neuroprotective effect of estrogen: Role of nonsynaptic NR2B-containing NMDA receptors. Brain Res. Bull. 2013, 93, 27–31. [Google Scholar] [CrossRef]
- Waters, E.M.; Mazid, S.; Dodos, M.; Puri, R.; Janssen, W.G.; Morrison, J.H.; McEwen, B.S.; Milner, T.A. Effects of estrogen and aging on synaptic morphology and distribution of phosphorylated Tyr1472 NR2B in the female rat hippocampus. Neurobiol. Aging 2019, 73, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Jumrussirikul, P.; Dinerman, J.; Dawson, T.M.; Dawson, V.L.; Ekelund, U.; Georgakopoulos, D.; Schramm, L.P.; Calkins, H.; Snyder, S.H.; Hare, J.M.; et al. Interaction between neuronal nitric oxide synthase and inhibitory G protein activity in heart rate regulation in conscious mice. J. Clin. Investig. 1998, 102, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Martins-Pinge, M.C.; Baraldi-Passy, I.; Lopes, O.U. Excitatory effects of nitric oxide within the rostral ventrolateral medulla of freely moving rats. Hypertension 1997, 30 Pt 2, 704–707. [Google Scholar] [CrossRef]
- Mayorov, D.N. Selective sensitization by nitric oxide of sympathetic baroreflex in rostral ventrolateral medulla of conscious rabbits. Hypertension 2005, 45, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, P.; Battaglia, A.; Ermirio, R.; Grossini, E.; Molinari, C.; Mary, D.A.; Vacca, G. Role of nitric oxide in the control of the heart rate within the nucleus ambiguus of rats. Neuroreport 2000, 11, 481–485. [Google Scholar] [CrossRef] [PubMed]
- McNeill, A.M.; Zhang, C.; Stanczyk, F.Z.; Duckles, S.P.; Krause, D.N. Estrogen increases endothelial nitric oxide synthase via estrogen receptors in rat cerebral blood vessels: Effect preserved after concurrent treatment with medroxyprogesterone acetate or progesterone. Stroke 2002, 33, 1685–1691. [Google Scholar] [CrossRef]
- Nevzati, E.; Shafighi, M.; Bakhtian, K.D.; Treiber, H.; Fandino, J.; Fathi, A.R. Estrogen induces nitric oxide production via nitric oxide synthase activation in endothelial cells. Acta Neurochir. Suppl. 2015, 120, 141–145. [Google Scholar]
- Robert, R.; Spitzer, J.A. Effects of female hormones (17beta-estradiol and progesterone) on nitric oxide production by alveolar macrophages in rats. Nitric Oxide 1997, 1, 453–462. [Google Scholar] [CrossRef]
- Chen, B.S.; Roche, K.W. Regulation of NMDA receptors by phosphorylation. Neuropharmacology 2007, 53, 362–368. [Google Scholar] [CrossRef]
- Tang, B.; Ji, Y.; Traub, R.J. Estrogen alters spinal NMDA receptor activity via a PKA signaling pathway in a visceral pain model in the rat. Pain 2008, 137, 540–549. [Google Scholar] [CrossRef]
- Ji, Y.; Bai, G.; Cao, D.Y.; Traub, R.J. Estradiol modulates visceral hyperalgesia by increasing thoracolumbar spinal GluN2B subunit activity in female rats. Neurogastroenterol. Motil. 2015, 27, 775–786. [Google Scholar] [CrossRef]
- Dedek, A.; Xu, J.; Lorenzo, L.; Godin, A.G.; Kandegedara, C.M.; Glavina, G.; Landrigan, J.A.; Lombroso, P.J.; De Koninck, Y.; Tsai, E.C.; et al. Sexual dimorphism in a neuronal mechanism of spinal hyperexcitability across rodent and human models of pathological pain. Brain 2022, 145, 1124–1138. [Google Scholar] [CrossRef]
- Kumar, S.; Lane, B.M.; Morrow, A.L. Differential effects of systemic ethanol administration on protein kinase cepsilon, gamma, and beta isoform expression, membrane translocation, and target phosphorylation: Reversal by chronic ethanol exposure. J. Pharmacol. Exp. Ther. 2006, 319, 1366–1375. [Google Scholar] [CrossRef]
- Li, H.F.; Mochly-Rosen, D.; Kendig, J.J. Protein kinase Cgamma mediates ethanol withdrawal hyper-responsiveness of NMDA receptor currents in spinal cord motor neurons. Br. J. Pharmacol. 2005, 144, 301–307. [Google Scholar] [CrossRef]
- Lai, C.-C.; Hsu, J.-W.; Cheng, Y.-S.; Lin, H.-H. Involvement of metabotropic glutamate receptor 5 in ethanol regulation of NMDA receptor activity in rat substantia gelatinosa neurons. Life Sci. 2019, 233, 116729. [Google Scholar] [CrossRef] [PubMed]
- Reneau, J.; Reyland, M.E.; Popp, R.L. Acute ethanol exposure prevents PMA-mediated augmentation of N-methyl-D-aspartate receptor function in primary cultured cerebellar granule cells. Alcohol 2011, 45, 595–605. [Google Scholar] [CrossRef]
- Brenner, G.J.; Ji, R.R.; Shaffer, S.; Woolf, C.J. Peripheral noxious stimulation induces phosphorylation of the NMDA receptor NR1 subunit at the PKC-dependent site, serine-896, in spinal cord dorsal horn neurons. Eur. J. Neurosci. 2004, 20, 375–384. [Google Scholar] [CrossRef]
- Chiu, H.-Y.; Lin, H.-H.; Lai, C.-C. Potentiation of spinal NMDA-mediated nociception by cocaine- and amphetamine-regulated transcript peptide via PKA and PKC signaling pathways in rats. Regul. Pept. 2009, 158, 77–85. [Google Scholar] [CrossRef]
- Vanderhorst, V.G.; Gustafsson, J.A.; Ulfhake, B. Estrogen receptor-alpha and -beta immunoreactive neurons in the brainstem and spinal cord of male and female mice: Relationships to monoaminergic, cholinergic, and spinal projection systems. J. Comp. Neurol. 2005, 488, 152–179. [Google Scholar] [CrossRef] [PubMed]
- Brailoiu, E.; Dun, S.L.; Brailoiu, G.C.; Mizuo, K.; Sklar, L.A.; Oprea, T.I.; Prossnitz, E.R.; Dun, N.J. Distribution and characterization of estrogen receptor G protein-coupled receptor 30 in the rat central nervous system. J. Endocrinol. 2007, 193, 311–321. [Google Scholar] [CrossRef]
- Parkash, J.; d’Anglemont de Tassigny, X.; Bellefontaine, N.; Campagne, C.; Mazure, D.; Buee-Scherrer, V.; Prevot, V. Phosphorylation of N-methyl-D-aspartic acid receptor-associated neuronal nitric oxide synthase depends on estrogens and modulates hypothalamic nitric oxide production during the ovarian cycle. Endocrinology 2010, 151, 2723–2735. [Google Scholar] [CrossRef] [PubMed]
- Shih, C.D. Activation of estrogen receptor beta-dependent nitric oxide signaling mediates the hypotensive effects of estrogen in the rostral ventrolateral medulla of anesthetized rats. J. Biomed. Sci. 2009, 16, 60. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.-C.; Yuan, Z.-F.; Chu, L.-Y.; Chuang, K.-T.; Lin, H.-H. Roles of cocaine- and amphetamine-regulated transcript peptide in the rostral ventrolateral medulla in cardiovascular regulation in rats. Brain Res. 2019, 1710, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, W.-K.; Lin, H.-H.; Lai, C.-C. Involvement of protein kinase C and Src tyrosine kinase in acute tolerance to ethanol inhibition of spinal NMDA-induced pressor responses in rats. Br. J. Pharmacol. 2009, 158, 806–818. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Situmorang, J.H.; Lin, H.-H.; Islam, M.S.; Lai, C.-C. Ovariectomy Exacerbates Acute Ethanol-Induced Tachycardia: Role of Nitric Oxide and NMDA Receptors in the Rostral Ventrolateral Medulla. Int. J. Mol. Sci. 2023, 24, 5087. https://doi.org/10.3390/ijms24065087
Situmorang JH, Lin H-H, Islam MS, Lai C-C. Ovariectomy Exacerbates Acute Ethanol-Induced Tachycardia: Role of Nitric Oxide and NMDA Receptors in the Rostral Ventrolateral Medulla. International Journal of Molecular Sciences. 2023; 24(6):5087. https://doi.org/10.3390/ijms24065087
Chicago/Turabian StyleSitumorang, Jiro Hasegawa, Hsun-Hsun Lin, Md Sharyful Islam, and Chih-Chia Lai. 2023. "Ovariectomy Exacerbates Acute Ethanol-Induced Tachycardia: Role of Nitric Oxide and NMDA Receptors in the Rostral Ventrolateral Medulla" International Journal of Molecular Sciences 24, no. 6: 5087. https://doi.org/10.3390/ijms24065087
APA StyleSitumorang, J. H., Lin, H.-H., Islam, M. S., & Lai, C.-C. (2023). Ovariectomy Exacerbates Acute Ethanol-Induced Tachycardia: Role of Nitric Oxide and NMDA Receptors in the Rostral Ventrolateral Medulla. International Journal of Molecular Sciences, 24(6), 5087. https://doi.org/10.3390/ijms24065087