

Towards a New Biomarker for Diabetic Retinopathy: Exploring RBP3 Structure and Retinoids Binding for Functional Imaging of Eyes In Vivo


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Diabetic Retinopathy
1.2. Eye Imaging
2. Diabetic Retinopathy
2.1. Clinical Progression
2.2. Molecular Component
2.3. Biomarkers
2.4. Non-Invasive Tracing of Disease Manifestations
3. RBP3/Two-Photon
3.1. RBP3
3.1.1. Historical
3.1.2. Gene Duplication
3.1.3. Ligand Binding
3.1.4. Link to DR
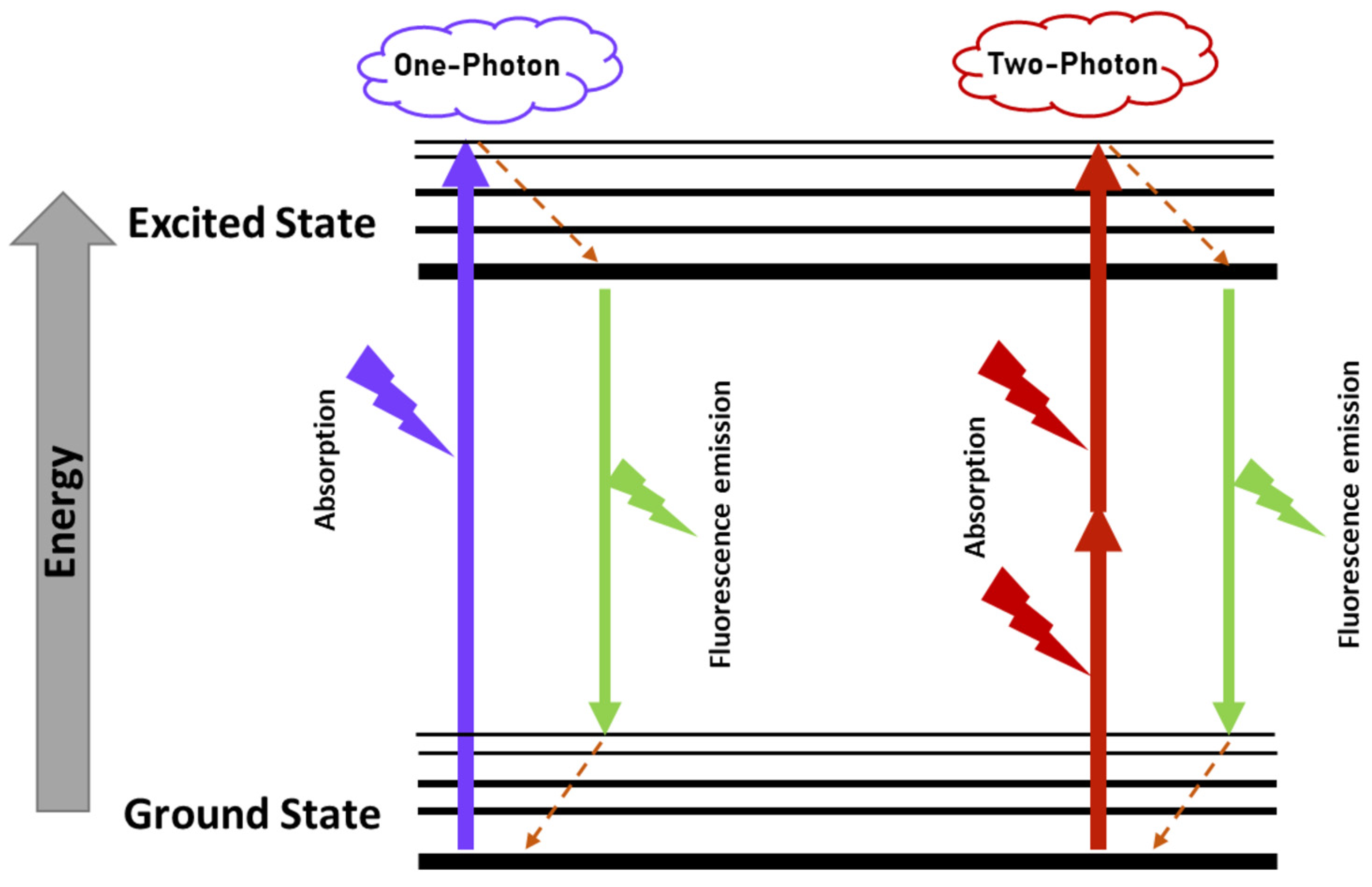
3.2. Two-Photon Excitation Fluorescence
3.2.1. Historical
3.2.2. Clinical Applications
3.2.3. Retinoids/Phasor
3.2.4. Possible Applications in a DR Context
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Frudd, K.; Sivaprasad, S.; Raman, R.; Krishnakumar, S.; Revathy, Y.R.; Turowski, P. Diagnostic circulating biomarkers to detect vision-threatening diabetic retinopathy: Potential screening tool of the future? Acta Ophthalmol. 2022, 100, e648–e668. [Google Scholar] [CrossRef]
- Berkowitz, B.A. Preventing diabetic retinopathy by mitigating subretinal space oxidative stress in vivo. Vis. Neurosci. 2020, 37, E002. [Google Scholar] [CrossRef]
- Jenkins, A.J.; Joglekar, M.V.; Hardikar, A.A.; Keech, A.C.; O’Neal, D.N.; Januszewski, A.S. Biomarkers in Diabetic Retinopathy. Rev. Diabet. Stud. 2015, 12, 159–195. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; He, M.; Congdon, N. The worldwide epidemic of diabetic retinopathy. Indian J. Ophthalmol. 2012, 60, 428–431. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lo, A.C.Y. Diabetic Retinopathy: Pathophysiology and Treatments. Int. J. Mol. Sci. 2018, 19, 1816. [Google Scholar] [CrossRef] [PubMed]
- Jampol, L.M.; Glassman, A.R.; Sun, J. Evaluation and Care of Patients with Diabetic Retinopathy. N. Engl. J. Med. 2020, 382, 1629–1637. [Google Scholar] [CrossRef] [PubMed]
- Mansour, S.E.; Browning, D.J.; Wong, K.; Flynn, H.W., Jr.; Bhavsar, A.R. The Evolving Treatment of Diabetic Retinopathy. Clin. Ophthalmol. 2020, 14, 653–678. [Google Scholar] [CrossRef]
- Wells, J.A.; Glassman, A.R.; Ayala, A.R.; Jampol, L.M.; Bressler, N.M.; Bressler, S.B.; Brucker, A.J.; Ferris, F.L.; Hampton, G.R.; Jhaveri, C.; et al. Aflibercept, Bevacizumab, or Ranibizumab for Diabetic Macular Edema: Two-Year Results from a Comparative Effectiveness Randomized Clinical Trial. Ophthalmology 2016, 123, 1351–1359. [Google Scholar] [CrossRef]
- Cecilia, O.M.; Jose Alberto, C.G.; Jose, N.P.; Ernesto German, C.M.; Ana Karen, L.C.; Luis Miguel, R.P.; Ricardo Raul, R.R.; Adolfo Daniel, R.C. Oxidative Stress as the Main Target in Diabetic Retinopathy Pathophysiology. J. Diabetes Res. 2019, 2019, 8562408. [Google Scholar] [CrossRef]
- Gardner, T.W.; Chew, E.Y. Future opportunities in diabetic retinopathy research. Curr. Opin. Endocrinol. Diabetes Obes. 2016, 23, 91–96. [Google Scholar] [CrossRef]
- Ting, D.S.; Tan, K.A.; Phua, V.; Tan, G.S.; Wong, C.W.; Wong, T.Y. Biomarkers of Diabetic Retinopathy. Curr. Diabet. Rep. 2016, 16, 125. [Google Scholar] [CrossRef] [PubMed]
- Antonetti, D.A.; Barber, A.J.; Bronson, S.K.; Freeman, W.M.; Gardner, T.W.; Jefferson, L.S.; Kester, M.; Kimball, S.R.; Krady, J.K.; LaNoue, K.F.; et al. Diabetic retinopathy: Seeing beyond glucose-induced microvascular disease. Diabetes 2006, 55, 2401–2411. [Google Scholar] [CrossRef] [PubMed]
- Safi, H.; Safi, S.; Hafezi-Moghadam, A.; Ahmadieh, H. Early detection of diabetic retinopathy. Surv. Ophthalmol. 2018, 63, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Malechka, V.V.; Moiseyev, G.; Takahashi, Y.; Shin, Y.; Ma, J.X. Impaired Rhodopsin Generation in the Rat Model of Diabetic Retinopathy. Am. J. Pathol. 2017, 187, 2222–2231. [Google Scholar] [CrossRef]
- Parisi, V.; Uccioli, L. Visual electrophysiological responses in persons with type 1 diabetes. Diabetes Metab. Res. Rev. 2001, 17, 12–18. [Google Scholar] [CrossRef]
- Bearse, M.A., Jr.; Han, Y.; Schneck, M.E.; Barez, S.; Jacobsen, C.; Adams, A.J. Local multifocal oscillatory potential abnormalities in diabetes and early diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3259–3265. [Google Scholar] [CrossRef]
- Han, Y.; Bearse, M.A., Jr.; Schneck, M.E.; Barez, S.; Jacobsen, C.H.; Adams, A.J. Multifocal electroretinogram delays predict sites of subsequent diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2004, 45, 948–954. [Google Scholar] [CrossRef]
- Adams, A.J.; Bearse, M.A., Jr. Retinal neuropathy precedes vasculopathy in diabetes: A function-based opportunity for early treatment intervention? Clin. Exp. Optom. 2012, 95, 256–265. [Google Scholar] [CrossRef]
- McAnany, J.J.; Park, J.C. Temporal Frequency Abnormalities in Early-Stage Diabetic Retinopathy Assessed by Electroretinography. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4871–4879. [Google Scholar] [CrossRef] [PubMed]
- McAnany, J.J.; Park, J.C.; Chau, F.Y.; Leiderman, Y.I.; Lim, J.I.; Blair, N.P. Amplitude Loss of the High-Frequency Flicker Electroretinogram in Early Diabetic Retinopathy. Retina 2019, 39, 2032–2039. [Google Scholar] [CrossRef]
- Daley, M.L.; Watzke, R.C.; Riddle, M.C. Early loss of blue-sensitive color vision in patients with type I diabetes. Diabetes Care 1987, 10, 777–781. [Google Scholar] [CrossRef] [PubMed]
- Henson, D.B.; North, R.V. Dark adaptation in diabetes mellitus. Br. J. Ophthalmol. 1979, 63, 539–541. [Google Scholar] [CrossRef] [PubMed]
- Lieth, E.; Gardner, T.W.; Barber, A.J.; Antonetti, D.A. Retinal neurodegeneration: Early pathology in diabetes. Clin. Exp. Ophthalmol. 2000, 28, 3–8. [Google Scholar] [CrossRef]
- Sohn, E.H.; van Dijk, H.W.; Jiao, C.; Kok, P.H.; Jeong, W.; Demirkaya, N.; Garmager, A.; Wit, F.; Kucukevcilioglu, M.; van Velthoven, M.E.; et al. Retinal neurodegeneration may precede microvascular changes characteristic of diabetic retinopathy in diabetes mellitus. Proc. Natl. Acad. Sci. USA 2016, 113, E2655–E2664. [Google Scholar] [CrossRef]
- Simo, R.; Stitt, A.W.; Gardner, T.W. Neurodegeneration in diabetic retinopathy: Does it really matter? Diabetologia 2018, 61, 1902–1912. [Google Scholar] [CrossRef] [PubMed]
- Pusparajah, P.; Lee, L.H.; Abdul Kadir, K. Molecular Markers of Diabetic Retinopathy: Potential Screening Tool of the Future? Front. Physiol. 2016, 7, 200. [Google Scholar] [CrossRef]
- Catalani, E.; Cervia, D. Diabetic retinopathy: A matter of retinal ganglion cell homeostasis. Neural Regen. Res. 2020, 15, 1253–1254. [Google Scholar] [CrossRef]
- Potilinski, M.C.; Lorenc, V.; Perisset, S.; Gallo, J.E. Mechanisms behind Retinal Ganglion Cell Loss in Diabetes and Therapeutic Approach. Int. J. Mol. Sci. 2020, 21, 2351. [Google Scholar] [CrossRef]
- Tonade, D.; Kern, T.S. Photoreceptor cells and RPE contribute to the development of diabetic retinopathy. Prog. Retin. Eye Res. 2021, 83, 100919. [Google Scholar] [CrossRef]
- Zeng, S.; Zhang, T.; Madigan, M.C.; Fernando, N.; Aggio-Bruce, R.; Zhou, F.; Pierce, M.; Chen, Y.; Huang, L.; Natoli, R.; et al. Interphotoreceptor Retinoid-Binding Protein (IRBP) in Retinal Health and Disease. Front. Cell NeuroSci. 2020, 14, 577935. [Google Scholar] [CrossRef]
- Garcia-Ramirez, M.; Hernandez, C.; Villarroel, M.; Canals, F.; Alonso, M.A.; Fortuny, R.; Masmiquel, L.; Navarro, A.; Garcia-Arumi, J.; Simo, R. Interphotoreceptor retinoid-binding protein (IRBP) is downregulated at early stages of diabetic retinopathy. Diabetologia 2009, 52, 2633–2641. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, F.; Ghosh, D. Focus on Molecules: Interphotoreceptor retinoid-binding protein (IRBP). Exp. Eye Res. 2008, 86, 169–170. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Li, S.; Nusinowitz, S.; Lloyd, M.; Hu, J.; Radu, R.A.; Bok, D.; Travis, G.H. The role of interphotoreceptor retinoid-binding protein on the translocation of visual retinoids and function of cone photoreceptors. J. NeuroSci. 2009, 29, 1486–1495. [Google Scholar] [CrossRef] [PubMed]
- Kusakabe, T.G.; Takimoto, N.; Jin, M.; Tsuda, M. Evolution and the origin of the visual retinoid cycle in vertebrates. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2897–2910. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Fernandez, F. Interphotoreceptor retinoid-binding protein--an old gene for new eyes. Vis. Res. 2003, 43, 3021–3036. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Blakeley, L.R.; Cornwall, M.C.; Crouch, R.K.; Wiggert, B.N.; Koutalos, Y. Interphotoreceptor retinoid-binding protein is the physiologically relevant carrier that removes retinol from rod photoreceptor outer segments. Biochemistry 2007, 46, 8669–8679. [Google Scholar] [CrossRef]
- Yokomizo, H.; Maeda, Y.; Park, K.; Clermont, A.C.; Hernandez, S.L.; Fickweiler, W.; Li, Q.; Wang, C.H.; Paniagua, S.M.; Simao, F.; et al. Retinol binding protein 3 is increased in the retina of patients with diabetes resistant to diabetic retinopathy. Sci. Transl. Med. 2019, 11, eaau6627. [Google Scholar] [CrossRef]
- Vujosevic, S.; Aldington, S.J.; Silva, P.; Hernandez, C.; Scanlon, P.; Peto, T.; Simo, R. Screening for diabetic retinopathy: New perspectives and challenges. Lancet Diabetes Endocrinol. 2020, 8, 337–347. [Google Scholar] [CrossRef]
- Aspelund, T.; Thornorisdottir, O.; Olafsdottir, E.; Gudmundsdottir, A.; Einarsdottir, A.B.; Mehlsen, J.; Einarsson, S.; Palsson, O.; Einarsson, G.; Bek, T.; et al. Individual risk assessment and information technology to optimise screening frequency for diabetic retinopathy. Diabetologia 2011, 54, 2525–2532. [Google Scholar] [CrossRef]
- Andersson, E.; Persson, S.; Hallen, N.; Ericsson, A.; Thielke, D.; Lindgren, P.; Steen Carlsson, K.; Jendle, J. Costs of diabetes complications: Hospital-based care and absence from work for 392,200 people with type 2 diabetes and matched control participants in Sweden. Diabetologia 2020, 63, 2582–2594. [Google Scholar] [CrossRef]
- Entenberg, D.; Oktay, M.H.; Condeelis, J.S. Intravital imaging to study cancer progression and metastasis. Nat. Rev. Cancer 2023, 23, 25–42. [Google Scholar] [CrossRef]
- Li, H.; Yu, J.; Zhang, R.; Li, X.; Zheng, W. Two-photon excitation fluorescence lifetime imaging microscopy: A promising diagnostic tool for digestive tract tumors. J. Innov. Opt. Health Sci. 2019, 12, 1930009. [Google Scholar] [CrossRef]
- Helmchen, F.; Denk, W. Deep tissue two-photon microscopy. Nat. Methods 2005, 2, 932–940. [Google Scholar] [CrossRef] [PubMed]
- Denk, W.; Strickler, J.H.; Webb, W.W. Two-photon laser scanning fluorescence microscopy. Science 1990, 248, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Noh, C.K.; Lim, C.S.; Lee, G.H.; Cho, M.K.; Lee, H.W.; Roh, J.; Kim, Y.B.; Lee, E.; Park, B.; Kim, H.M.; et al. A Diagnostic Method for Gastric Cancer Using Two-Photon Microscopy with Enzyme-Selective Fluorescent Probes: A Pilot Study. Front. Oncol. 2021, 11, 634219. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Zhang, C.; Li, Y.; Yan, J.; Xu, Y.; Pan, Y.; Hu, R.; Liu, L.; Qu, J. Human serum albumin gradient in serous ovarian cancer cryosections measured by fluorescence lifetime. BioMed. Opt. Express 2021, 12, 1195–1204. [Google Scholar] [CrossRef]
- Cho, M.K.; Lim, C.S.; Sarkar, A.R.; Lee, H.W.; Choi, H.J.; Noh, C.-K.; Shin, S.J.; Kim, H.M. A two-photon ratiometric probe for detection of hNQO1 enzyme activity in human colon tissue. Sens. Actuators B Chem. 2018, 272, 203–210. [Google Scholar] [CrossRef]
- Palczewska, G.; Dong, Z.; Golczak, M.; Hunter, J.J.; Williams, D.R.; Alexander, N.S.; Palczewski, K. Noninvasive two-photon microscopy imaging of mouse retina and retinal pigment epithelium through the pupil of the eye. Nat. Med. 2014, 20, 785–789. [Google Scholar] [CrossRef]
- Maeda, A.; Palczewska, G.; Golczak, M.; Kohno, H.; Dong, Z.; Maeda, T.; Palczewski, K. Two-photon microscopy reveals early rod photoreceptor cell damage in light-exposed mutant mice. Proc. Natl. Acad. Sci. USA 2014, 111, E1428–E1437. [Google Scholar] [CrossRef]
- Wang, B.G.; Konig, K.; Halbhuber, K.J. Two-photon microscopy of deep intravital tissues and its merits in clinical research. J. Microsc. 2010, 238, 1–20. [Google Scholar] [CrossRef]
- Palczewska, G.; Stremplewski, P.; Suh, S.; Alexander, N.; Salom, D.; Dong, Z.; Ruminski, D.; Choi, E.H.; Sears, A.E.; Kern, T.S.; et al. Two-photon imaging of the mammalian retina with ultrafast pulsing laser. JCI Insight 2018, 3, e121555. [Google Scholar] [CrossRef] [PubMed]
- Palczewska, G.; Boguslawski, J.; Stremplewski, P.; Kornaszewski, L.; Zhang, J.; Dong, Z.; Liang, X.X.; Gratton, E.; Vogel, A.; Wojtkowski, M.; et al. Noninvasive two-photon optical biopsy of retinal fluorophores. Proc. Natl. Acad. Sci. USA 2020, 117, 22532–22543. [Google Scholar] [CrossRef] [PubMed]
- Boguslawski, J.; Palczewska, G.; Tomczewski, S.; Milkiewicz, J.; Kasprzycki, P.; Stachowiak, D.; Komar, K.; Marzejon, M.J.; Sikorski, B.L.; Hudzikowski, A.; et al. In vivo imaging of the human eye using a 2-photon-excited fluorescence scanning laser ophthalmoscope. J. Clin. Investig. 2022, 132. [Google Scholar] [CrossRef] [PubMed]
- Kang, Q.; Yang, C. Oxidative stress and diabetic retinopathy: Molecular mechanisms, pathogenetic role and therapeutic implications. Redox Biol. 2020, 37, 101799. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef]
- Mahajan, N.; Arora, P.; Sandhir, R. Perturbed Biochemical Pathways and Associated Oxidative Stress Lead to Vascular Dysfunctions in Diabetic Retinopathy. Oxid. Med. Cell Longev. 2019, 2019, 8458472. [Google Scholar] [CrossRef]
- Cheung, N.; Mitchell, P.; Wong, T.Y. Diabetic retinopathy. Lancet 2010, 376, 124–136. [Google Scholar] [CrossRef]
- Wu, M.Y.; Yiang, G.T.; Lai, T.T.; Li, C.J. The Oxidative Stress and Mitochondrial Dysfunction during the Pathogenesis of Diabetic Retinopathy. Oxid. Med. Cell Longev. 2018, 2018, 3420187. [Google Scholar] [CrossRef]
- Antonetti, D.A.; Silva, P.S.; Stitt, A.W. Current understanding of the molecular and cellular pathology of diabetic retinopathy. Nat. Rev. Endocrinol. 2021, 17, 195–206. [Google Scholar] [CrossRef]
- Brownlee, M.; Cerami, A.; Vlassara, H. Advanced glycosylation end products in tissue and the biochemical basis of diabetic complications. N. Engl. J. Med. 1988, 318, 1315–1321. [Google Scholar] [CrossRef]
- Stitt, A.W. AGEs and diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4867–4874. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Chen, L.J.; Yu, J.; Wang, H.J.; Zhang, F.; Liu, Q.; Wu, J. Involvement of Advanced Glycation End Products in the Pathogenesis of Diabetic Retinopathy. Cell Physiol. Biochem. 2018, 48, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Packer, L.; Cadenas, E. Oxidants and antioxidants revisited. New concepts of oxidative stress. Free Radic. Res. 2007, 41, 951–952. [Google Scholar] [CrossRef]
- Al-Shabrawey, M.; Bartoli, M.; El-Remessy, A.B.; Ma, G.; Matragoon, S.; Lemtalsi, T.; Caldwell, R.W.; Caldwell, R.B. Role of NADPH oxidase and Stat3 in statin-mediated protection against diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3231–3238. [Google Scholar] [CrossRef]
- Al-Shabrawey, M.; Rojas, M.; Sanders, T.; Behzadian, A.; El-Remessy, A.; Bartoli, M.; Parpia, A.K.; Liou, G.; Caldwell, R.B. Role of NADPH oxidase in retinal vascular inflammation. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3239–3244. [Google Scholar] [CrossRef]
- Berkowitz, B.A.; Kern, T.S.; Bissig, D.; Patel, P.; Bhatia, A.; Kefalov, V.J.; Roberts, R. Systemic Retinaldehyde Treatment Corrects Retinal Oxidative Stress, Rod Dysfunction, and Impaired Visual Performance in Diabetic Mice. Investig. Ophthalmol. Vis. Sci. 2015, 56, 6294–6303. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, R.B.; Bartoli, M.; Behzadian, M.A.; El-Remessy, A.E.; Al-Shabrawey, M.; Platt, D.H.; Liou, G.I.; Caldwell, R.W. Vascular endothelial growth factor and diabetic retinopathy: Role of oxidative stress. Curr. Drug Targets 2005, 6, 511–524. [Google Scholar] [CrossRef]
- Du, Y.; Veenstra, A.; Palczewski, K.; Kern, T.S. Photoreceptor cells are major contributors to diabetes-induced oxidative stress and local inflammation in the retina. Proc. Natl. Acad. Sci. USA 2013, 110, 16586–16591. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, M.; Chan, P.S.; Kern, T.S.; Kowluru, R.A. Oxidative damage in the retinal mitochondria of diabetic mice: Possible protection by superoxide dismutase. Investig. Ophthalmol. Vis. Sci. 2007, 48, 3805–3811. [Google Scholar] [CrossRef]
- Kowluru, R.A.; Tang, J.; Kern, T.S. Abnormalities of retinal metabolism in diabetes and experimental galactosemia. VII. Effect of long-term administration of antioxidants on the development of retinopathy. Diabetes 2001, 50, 1938–1942. [Google Scholar] [CrossRef]
- Kowluru, R.A.; Kowluru, A.; Mishra, M.; Kumar, B. Oxidative stress and epigenetic modifications in the pathogenesis of diabetic retinopathy. Prog. Retin. Eye Res. 2015, 48, 40–61. [Google Scholar] [CrossRef] [PubMed]
- Berkowitz, B.A.; Gradianu, M.; Bissig, D.; Kern, T.S.; Roberts, R. Retinal ion regulation in a mouse model of diabetic retinopathy: Natural history and the effect of Cu/Zn superoxide dismutase overexpression. Investig. Ophthalmol. Vis. Sci. 2009, 50, 2351–2358. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; He, J.; Johnson, D.; Wei, Y.; Liu, Y.; Wang, S.; Lutty, G.A.; Duh, E.J.; Semba, R.D. Deletion of placental growth factor prevents diabetic retinopathy and is associated with Akt activation and HIF1alpha-VEGF pathway inhibition. Diabetes 2015, 64, 200–212. [Google Scholar] [CrossRef]
- Lupo, G.; Motta, C.; Giurdanella, G.; Anfuso, C.D.; Alberghina, M.; Drago, F.; Salomone, S.; Bucolo, C. Role of phospholipases A2 in diabetic retinopathy: In vitro and in vivo studies. Biochem. Pharmacol. 2013, 86, 1603–1613. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.I.; Hykin, P.G.; Gregor, Z.J.; Boulton, M.; Cree, I.A. Angiopoietin concentrations in diabetic retinopathy. Br. J. Ophthalmol. 2005, 89, 480–483. [Google Scholar] [CrossRef]
- Rangasamy, S.; Srinivasan, R.; Maestas, J.; McGuire, P.G.; Das, A. A potential role for angiopoietin 2 in the regulation of the blood-retinal barrier in diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3784–3791. [Google Scholar] [CrossRef]
- Li, X.; Cai, Y.; Wang, Y.S.; Shi, Y.Y.; Hou, W.; Xu, C.S.; Wang, H.Y.; Ye, Z.; Yao, L.B.; Zhang, J. Hyperglycaemia exacerbates choroidal neovascularisation in mice via the oxidative stress-induced activation of STAT3 signalling in RPE cells. PLoS ONE 2012, 7, e47600. [Google Scholar] [CrossRef]
- Liu, H.; Tang, J.; Du, Y.; Lee, C.A.; Golczak, M.; Muthusamy, A.; Antonetti, D.A.; Veenstra, A.A.; Amengual, J.; von Lintig, J.; et al. Retinylamine Benefits Early Diabetic Retinopathy in Mice. J. Biol. Chem. 2015, 290, 21568–21579. [Google Scholar] [CrossRef]
- Bailey, T.A.; Kanuga, N.; Romero, I.A.; Greenwood, J.; Luthert, P.J.; Cheetham, M.E. Oxidative stress affects the junctional integrity of retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 2004, 45, 675–684. [Google Scholar] [CrossRef]
- MacGregor, L.C.; Matschinsky, F.M. Treatment with aldose reductase inhibitor or with myo-inositol arrests deterioration of the electroretinogram of diabetic rats. J. Clin. Investig. 1985, 76, 887–889. [Google Scholar] [CrossRef]
- MacGregor, L.C.; Matschinsky, F.M. Altered retinal metabolism in diabetes. II. Measurement of sodium-potassium ATPase and total sodium and potassium in individual retinal layers. J. Biol. Chem. 1986, 261, 4052–4058. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, L.C.; Matschinsky, F.M. Experimental diabetes mellitus impairs the function of the retinal pigmented epithelium. Metabolism 1986, 35, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, A.; Kowluru, R.A.; Yamazaki, A. Functional alterations of G-proteins in diabetic rat retina: A possible explanation for the early visual abnormalities in diabetes mellitus. Diabetologia 1992, 35, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Ostroy, S.E.; Frede, S.M.; Wagner, E.F.; Gaitatzes, C.G.; Janle, E.M. Decreased rhodopsin regeneration in diabetic mouse eyes. Investig. Ophthalmol. Vis. Sci. 1994, 35, 3905–3909. [Google Scholar]
- Kern, T.S.; Engerman, R.L. Capillary lesions develop in retina rather than cerebral cortex in diabetes and experimental galactosemia. Arch. Ophthalmol. 1996, 114, 306–310. [Google Scholar] [CrossRef]
- Ostroy, S.E. Altered rhodopsin regeneration in diabetic mice caused by acid conditions within the rod photoreceptors. Curr. Eye Res. 1998, 17, 979–985. [Google Scholar] [CrossRef]
- Lahdenranta, J.; Pasqualini, R.; Schlingemann, R.O.; Hagedorn, M.; Stallcup, W.B.; Bucana, C.D.; Sidman, R.L.; Arap, W. An anti-angiogenic state in mice and humans with retinal photoreceptor cell degeneration. Proc. Natl. Acad. Sci. USA 2001, 98, 10368–10373. [Google Scholar] [CrossRef]
- Colantuoni, A.; Longoni, B.; Marchiafava, P.L. Retinal photoreceptors of Syrian hamsters undergo oxidative stress during streptozotocin-induced diabetes. Diabetologia 2002, 45, 121–124. [Google Scholar] [CrossRef]
- Phipps, J.A.; Fletcher, E.L.; Vingrys, A.J. Paired-flash identification of rod and cone dysfunction in the diabetic rat. Investig. Ophthalmol. Vis. Sci. 2004, 45, 4592–4600. [Google Scholar] [CrossRef]
- Wong, V.H.; Vingrys, A.J.; Bui, B.V. Glial and neuronal dysfunction in streptozotocin-induced diabetic rats. J. Ocul. Biol. Dis. Infor. 2011, 4, 42–50. [Google Scholar] [CrossRef]
- Berkowitz, B.A.; Grady, E.M.; Khetarpal, N.; Patel, A.; Roberts, R. Oxidative stress and light-evoked responses of the posterior segment in a mouse model of diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2015, 56, 606–615. [Google Scholar] [CrossRef]
- Du, Y.; Cramer, M.; Lee, C.A.; Tang, J.; Muthusamy, A.; Antonetti, D.A.; Jin, H.; Palczewski, K.; Kern, T.S. Adrenergic and serotonin receptors affect retinal superoxide generation in diabetic mice: Relationship to capillary degeneration and permeability. FASEB J. 2015, 29, 2194–2204. [Google Scholar] [CrossRef] [PubMed]
- Kern, T.S.; Berkowitz, B.A. Photoreceptors in diabetic retinopathy. J. Diabetes Investig. 2015, 6, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Samuels, I.S.; Bell, B.A.; Pereira, A.; Saxon, J.; Peachey, N.S. Early retinal pigment epithelium dysfunction is concomitant with hyperglycemia in mouse models of type 1 and type 2 diabetes. J. NeuroPhysiol. 2015, 113, 1085–1099. [Google Scholar] [CrossRef]
- Berkowitz, B.A.; Bissig, D.; Roberts, R. MRI of rod cell compartment-specific function in disease and treatment in vivo. Prog. Retin. Eye Res. 2016, 51, 90–106. [Google Scholar] [CrossRef] [PubMed]
- Kern, T.S. Do photoreceptor cells cause the development of retinal vascular disease? Vis. Res. 2017, 139, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Kern, T.S.; Song, B.; Stuebe, C. Mechanistic Insights into Pathological Changes in the Diabetic Retina: Implications for Targeting Diabetic Retinopathy. Am J. Pathol. 2017, 187, 9–19. [Google Scholar] [CrossRef]
- Tarchick, M.J.; Cutler, A.H.; Trobenter, T.D.; Kozlowski, M.R.; Makowski, E.R.; Holoman, N.; Shao, J.; Shen, B.; Anand-Apte, B.; Samuels, I.S. Endogenous insulin signaling in the RPE contributes to the maintenance of rod photoreceptor function in diabetes. Exp. Eye Res. 2019, 180, 63–74. [Google Scholar] [CrossRef]
- de Gooyer, T.E.; Stevenson, K.A.; Humphries, P.; Simpson, D.A.; Gardiner, T.A.; Stitt, A.W. Retinopathy is reduced during experimental diabetes in a mouse model of outer retinal degeneration. Investig. Ophthalmol. Vis. Sci. 2006, 47, 5561–5568. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, Y.S.; Noh, H.S.; Kang, S.S.; Cheon, E.W.; Park, S.K.; Lee, B.J.; Choi, W.S.; Cho, G.J. Changes in rhodopsin kinase and transducin in the rat retina in early-stage diabetes. Exp. Eye Res. 2005, 80, 753–760. [Google Scholar] [CrossRef]
- Liu, H.; Tang, J.; Du, Y.; Saadane, A.; Samuels, I.; Veenstra, A.; Kiser, J.Z.; Palczewski, K.; Kern, T.S. Transducin1, Phototransduction and the Development of Early Diabetic Retinopathy. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1538–1546. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, F.; Sung, D.; Haswell, K.M.; Tsin, A.; Ghosh, D. Thiol-dependent antioxidant activity of interphotoreceptor retinoid-binding protein. Exp. Eye Res. 2014, 120, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Bringmann, A.; Pannicke, T.; Grosche, J.; Francke, M.; Wiedemann, P.; Skatchkov, S.N.; Osborne, N.N.; Reichenbach, A. Muller cells in the healthy and diseased retina. Prog. Retin. Eye Res. 2006, 25, 397–424. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Nelson, K.C.; Wu, M.; Sternberg, P., Jr.; Jones, D.P. Oxidative damage and protection of the RPE. Prog. Retin. Eye Res. 2000, 19, 205–221. [Google Scholar] [CrossRef] [PubMed]
- Plafker, S.M.; O’Mealey, G.B.; Szweda, L.I. Mechanisms for countering oxidative stress and damage in retinal pigment epithelium. Int. Rev. Cell Mol. Biol. 2012, 298, 135–177. [Google Scholar] [CrossRef]
- Giordano, C.R.; Roberts, R.; Krentz, K.A.; Bissig, D.; Talreja, D.; Kumar, A.; Terlecky, S.R.; Berkowitz, B.A. Catalase therapy corrects oxidative stress-induced pathophysiology in incipient diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3095–3102. [Google Scholar] [CrossRef]
- Muir, E.R.; Chandra, S.B.; De La Garza, B.H.; Velagapudi, C.; Abboud, H.E.; Duong, T.Q. Layer-Specific Manganese-Enhanced MRI of the Diabetic Rat Retina in Light and Dark Adaptation at 11.7 Tesla. Investig. Ophthalmol. Vis. Sci. 2015, 56, 4006–4012. [Google Scholar] [CrossRef]
- Saliba, A.; Du, Y.; Liu, H.; Patel, S.; Roberts, R.; Berkowitz, B.A.; Kern, T.S. Photobiomodulation Mitigates Diabetes-Induced Retinopathy by Direct and Indirect Mechanisms: Evidence from Intervention Studies in Pigmented Mice. PLoS ONE 2015, 10, e0139003. [Google Scholar] [CrossRef]
- Berkowitz, B.A.; Podolsky, R.H.; Lins-Childers, K.M.; Li, Y.; Qian, H. Outer Retinal Oxidative Stress Measured In Vivo Using QUEnch-assiSTed (QUEST) OCT. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1566–1570. [Google Scholar] [CrossRef]
- Zhang, T.; Gillies, M.; Wang, Y.; Shen, W.; Bahrami, B.; Zeng, S.; Zhu, M.; Yao, W.; Zhou, F.; Murray, M.; et al. Simvastatin protects photoreceptors from oxidative stress induced by all-trans-retinal, through the up-regulation of interphotoreceptor retinoid binding protein. Br. J. Pharmacol. 2019, 176, 2063–2078. [Google Scholar] [CrossRef]
- Lee, M.; Li, S.; Sato, K.; Jin, M. Interphotoreceptor Retinoid-Binding Protein Mitigates Cellular Oxidative Stress and Mitochondrial Dysfunction Induced by All-trans-Retinal. Investig. Ophthalmol. Vis. Sci. 2016, 57, 1553–1562. [Google Scholar] [CrossRef]
- Wang, K.; Zhu, X.; Zhang, K.; Zhou, F.; Zhu, L. Neuroprotective effect of tetramethylpyrazine against all-trans-retinal toxicity in the differentiated Y-79 cells via upregulation of IRBP expression. Exp. Cell Res. 2017, 359, 120–128. [Google Scholar] [CrossRef] [PubMed]
- van Veen, T.; Katial, A.; Shinohara, T.; Barrett, D.J.; Wiggert, B.; Chader, G.J.; Nickerson, J.M. Retinal photoreceptor neurons and pinealocytes accumulate mRNA for interphotoreceptor retinoid-binding protein (IRBP). FEBS Lett. 1986, 208, 133–137. [Google Scholar] [CrossRef]
- Wisard, J.; Faulkner, A.; Chrenek, M.A.; Waxweiler, T.; Waxweiler, W.; Donmoyer, C.; Liou, G.I.; Craft, C.M.; Schmid, G.F.; Boatright, J.H.; et al. Exaggerated eye growth in IRBP-deficient mice in early development. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5804–5811. [Google Scholar] [CrossRef]
- Markand, S.; Baskin, N.L.; Chakraborty, R.; Landis, E.; Wetzstein, S.A.; Donaldson, K.J.; Priyadarshani, P.; Alderson, S.E.; Sidhu, C.S.; Boatright, J.H.; et al. IRBP deficiency permits precocious ocular development and myopia. Mol. Vis. 2016, 22, 1291–1308. [Google Scholar] [PubMed]
- Liou, G.I.; Fei, Y.; Peachey, N.S.; Matragoon, S.; Wei, S.; Blaner, W.S.; Wang, Y.; Liu, C.; Gottesman, M.E.; Ripps, H. Early onset photoreceptor abnormalities induced by targeted disruption of the interphotoreceptor retinoid-binding protein gene. J. NeuroSci. 1998, 18, 4511–4520. [Google Scholar] [CrossRef] [PubMed]
- Ooi, X.Y.; Khan, R.; Choudhury, A.; Elisarraras, F.X.; Grigsby, J.; Obregon, B.; Tsin, A. Recent advances on visual cycle protein research and progress on clinical translation. Arch Clin. Exp. Ophthalmol. 2020, 2, 73–76. [Google Scholar] [CrossRef]
- Ishikawa, M.; Sawada, Y.; Yoshitomi, T. Structure and function of the interphotoreceptor matrix surrounding retinal photoreceptor cells. Exp. Eye Res. 2015, 133, 3–18. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, F.; Lippincott, J.; Powell, A.; Coleman, T. Rat CRISPR/Cas9 knockout of Interphotoreceptor retinoid-binding protein (IRBP, RBP3) suggests role in maintaining lamellar disc outer segment stability. Investig. Ophthalmol. Vis. Sci. 2020, 61, 1528. [Google Scholar]
- Wald, G. Molecular basis of visual excitation. Science 1968, 162, 230–239. [Google Scholar] [CrossRef]
- Kiser, P.D.; Golczak, M.; Palczewski, K. Chemistry of the retinoid (visual) cycle. Chem. Rev. 2014, 114, 194–232. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.; Haswell, K.M.; Sprada, M.; Gonzalez-Fernandez, F. Structure of zebrafish IRBP reveals fatty acid binding. Exp. Eye Res. 2015, 140, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Saari, J.C.; Noy, N. Interactions of all-trans-retinol and long-chain fatty acids with interphotoreceptor retinoid-binding protein. Biochemistry 1993, 32, 11311–11318. [Google Scholar] [CrossRef] [PubMed]
- Marmorstein, A.D.; Marmorstein, L.Y.; Sakaguchi, H.; Hollyfield, J.G. Spectral profiling of autofluorescence associated with lipofuscin, Bruch’s Membrane, and sub-RPE deposits in normal and AMD eyes. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2435–2441. [Google Scholar]
- Radu, R.A.; Mata, N.L.; Nusinowitz, S.; Liu, X.; Sieving, P.A.; Travis, G.H. Treatment with isotretinoin inhibits lipofuscin accumulation in a mouse model of recessive Stargardt’s macular degeneration. Proc. Natl. Acad. Sci. USA 2003, 100, 4742–4747. [Google Scholar] [CrossRef]
- Wiggert, B.; van Veen, T.; Kutty, G.; Lee, L.; Nickerson, J.; Si, J.S.; Nilsson, S.E.; Chader, G.J.; Narfstrom, K. An early decrease in interphotoreceptor retinoid-binding protein gene expression in Abyssinian cats homozygous for hereditary rod-cone degeneration. Cell Tissue Res. 1994, 278, 291–298. [Google Scholar] [CrossRef]
- Narfstrom, K.; Nilsson, S.E.; Wiggert, B.; Lee, L.; Chader, G.J.; van Veen, T. Reduced level of interphotoreceptor retinoid-binding protein (IRBP), a possible cause for retinal degeneration in the Abyssinian cat. Cell Tissue Res. 1989, 257, 631–639. [Google Scholar] [CrossRef]
- Arno, G.; Hull, S.; Robson, A.G.; Holder, G.E.; Cheetham, M.E.; Webster, A.R.; Plagnol, V.; Moore, A.T. Lack of Interphotoreceptor Retinoid Binding Protein Caused by Homozygous Mutation of RBP3 Is Associated with High Myopia and Retinal Dystrophy. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2358–2365. [Google Scholar] [CrossRef]
- Li, S.; Yang, Z.; Hu, J.; Gordon, W.C.; Bazan, N.G.; Haas, A.L.; Bok, D.; Jin, M. Secretory defect and cytotoxicity: The potential disease mechanisms for the retinitis pigmentosa (RP)-associated interphotoreceptor retinoid-binding protein (IRBP). J. Biol. Chem. 2013, 288, 11395–11406. [Google Scholar] [CrossRef]
- den Hollander, A.I.; McGee, T.L.; Ziviello, C.; Banfi, S.; Dryja, T.P.; Gonzalez-Fernandez, F.; Ghosh, D.; Berson, E.L. A homozygous missense mutation in the IRBP gene (RBP3) associated with autosomal recessive retinitis pigmentosa. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1864–1872. [Google Scholar] [CrossRef]
- Zheng, Q.; Ren, Y.; Tzekov, R.; Hua, S.; Li, M.; Pang, J.; Qu, J.; Li, W. iTRAQ-Based Proteomic Analysis of Visual Cycle-Associated Proteins in RPE of rd12 Mice before and after RPE65 Gene Delivery. J. Ophthalmol. 2015, 2015, 918473. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.; Ulyanova, T.; Organisciak, D.T.; Bennett, S.; Lakins, J.; Arnold, J.M.; Kutty, R.K.; Tenniswood, M.; vanVeen, T.; Darrow, R.M.; et al. Expression of multiple forms of clusterin during light-induced retinal degeneration. Curr. Eye Res. 2001, 23, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Adler, A.J.; Severin, K.M. Proteins of the bovine interphotoreceptor matrix: Tissues of origin. Exp. Eye Res. 1981, 32, 755–769. [Google Scholar] [CrossRef] [PubMed]
- Liou, G.I.; Bridges, C.D.; Fong, S.L.; Alvarez, R.A.; Gonzalez-Fernandez, F. Vitamin A transport between retina and pigment epithelium--an interstitial protein carrying endogenous retinol (interstitial retinol-binding protein). Vis. Res. 1982, 22, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Adler, A.J.; Evans, C.D. Rapid isolation of bovine interphotoreceptor retinol-binding protein. Biochim. Biophys. Acta 1983, 761, 217–222. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, F. Interphotoreceptor retinoid binding protein; myths and mysteries. J. Ophthalmic. Vis. Res. 2012, 7, 100–104. [Google Scholar] [PubMed]
- Nickerson, J.M.; Frey, R.A.; Ciavatta, V.T.; Stenkamp, D.L. Interphotoreceptor retinoid-binding protein gene structure in tetrapods and teleost fish. Mol. Vis. 2006, 12, 1565–1585. [Google Scholar]
- Fei, Y.; Matragoon, S.; Smith, S.B.; Overbeek, P.A.; Chen, S.; Zack, D.J.; Liou, G.I. Functional dissection of the promoter of the interphotoreceptor retinoid-binding protein gene: The cone-rod-homeobox element is essential for photoreceptor-specific expression in vivo. J. Biochem. 1999, 125, 1189–1199. [Google Scholar] [CrossRef]
- Nishida, A.; Furukawa, A.; Koike, C.; Tano, Y.; Aizawa, S.; Matsuo, I.; Furukawa, T. Otx2 homeobox gene controls retinal photoreceptor cell fate and pineal gland development. Nat. NeuroSci. 2003, 6, 1255–1263. [Google Scholar] [CrossRef]
- Wang, J.S.; Kefalov, V.J. An alternative pathway mediates the mouse and human cone visual cycle. Curr. Biol. 2009, 19, 1665–1669. [Google Scholar] [CrossRef]
- Sato, S.; Kefalov, V.J. cis Retinol oxidation regulates photoreceptor access to the retina visual cycle and cone pigment regeneration. J. Physiol. 2016, 594, 6753–6765. [Google Scholar] [CrossRef]
- Mata, N.L.; Radu, R.A.; Clemmons, R.C.; Travis, G.H. Isomerization and oxidation of vitamin a in cone-dominant retinas: A novel pathway for visual-pigment regeneration in daylight. Neuron 2002, 36, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Tang, P.H.; Kono, M.; Koutalos, Y.; Ablonczy, Z.; Crouch, R.K. New insights into retinoid metabolism and cycling within the retina. Prog. Retin. Eye Res. 2013, 32, 48–63. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.S.; Fong, S.L.; Bridges, C.D. Retinoids bound to interstitial retinol-binding protein during light and dark-adaptation. Vis. Res. 1989, 29, 1699–1709. [Google Scholar] [CrossRef]
- Saari, J.C.; Teller, D.C.; Crabb, J.W.; Bredberg, L. Properties of an interphotoreceptor retinoid-binding protein from bovine retina. J. Biol. Chem. 1985, 260, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Scott, B.L.; Bazan, N.G. Membrane docosahexaenoate is supplied to the developing brain and retina by the liver. Proc. Natl. Acad. Sci. USA 1989, 86, 2903–2907. [Google Scholar] [CrossRef]
- Jastrzebska, B.; Debinski, A.; Filipek, S.; Palczewski, K. Role of membrane integrity on G protein-coupled receptors: Rhodopsin stability and function. Prog. Lipid. Res. 2011, 50, 267–277. [Google Scholar] [CrossRef]
- Semenova, E.M.; Converse, C.A. Comparison between oleic acid and docosahexaenoic acid binding to interphotoreceptor retinoid-binding protein. Vis. Res. 2003, 43, 3063–3067. [Google Scholar] [CrossRef]
- Putilina, T.; Sittenfeld, D.; Chader, G.J.; Wiggert, B. Study of a fatty acid binding site of interphotoreceptor retinoid-binding protein using fluorescent fatty acids. Biochemistry 1993, 32, 3797–3803. [Google Scholar] [CrossRef]
- Lin, Z.Y.; Li, G.R.; Takizawa, N.; Si, J.S.; Gross, E.A.; Richardson, K.; Nickerson, J.M. Structure-function relationships in interphotoreceptor retinoid-binding protein (IRBP). Mol. Vis. 1997, 3, 17. [Google Scholar]
- Uehara, F.; Matthes, M.T.; Yasumura, D.; LaVail, M.M. Light-evoked changes in the interphotoreceptor matrix. Science 1990, 248, 1633–1636. [Google Scholar] [CrossRef]
- Hollyfield, J.G.; Fliesler, S.J.; Rayborn, M.E.; Fong, S.L.; Landers, R.A.; Bridges, C.D. Synthesis and secretion of interstitial retinol-binding protein by the human retina. Investig. Ophthalmol. Vis. Sci. 1985, 26, 58–67. [Google Scholar]
- Huang, B.; Karwoski, C.J. Light-evoked expansion of subretinal space volume in the retina of the frog. J. NeuroSci. 1992, 12, 4243–4252. [Google Scholar] [CrossRef]
- Yamamoto, F.; Steinberg, R.H. Effects of intravenous acetazolamide on retinal pH in the cat. Exp. Eye Res. 1992, 54, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Govardovskii, V.I.; Li, J.D.; Dmitriev, A.V.; Steinberg, R.H. Mathematical model of TMA+ diffusion and prediction of light-dependent subretinal hydration in chick retina. Investig. Ophthalmol. Vis. Sci. 1994, 35, 2712–2724. [Google Scholar]
- Li, J.D.; Gallemore, R.P.; Dmitriev, A.; Steinberg, R.H. Light-dependent hydration of the space surrounding photoreceptors in chick retina. Investig. Ophthalmol. Vis. Sci. 1994, 35, 2700–2711. [Google Scholar]
- Wolfensberger, T.J.; Dmitriev, A.V.; Govardovskii, V.I. Inhibition of membrane-bound carbonic anhydrase decreases subretinal pH and volume. Doc. Ophthalmol. 1999, 97, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Bissig, D.; Berkowitz, B.A. Light-dependent changes in outer retinal water diffusion in rats in vivo. Mol. Vis. 2012, 18, 2561. [Google Scholar]
- Li, Y.; Fariss, R.N.; Qian, J.W.; Cohen, E.D.; Qian, H. Light-Induced Thickening of Photoreceptor Outer Segment Layer Detected by Ultra-High Resolution OCT Imaging. Investig. Ophthalmol. Vis. Sci. 2016, 57, OCT105–OCT111. [Google Scholar] [CrossRef] [PubMed]
- Hammer, M.; Roggan, A.; Schweitzer, D.; Muller, G. Optical properties of ocular fundus tissues--an in vitro study using the double-integrating-sphere technique and inverse Monte Carlo simulation. Phys. Med. Biol. 1995, 40, 963–978. [Google Scholar] [CrossRef]
- Imanishi, Y.; Batten, M.L.; Piston, D.W.; Baehr, W.; Palczewski, K. Noninvasive two-photon imaging reveals retinyl ester storage structures in the eye. J. Cell Biol. 2004, 164, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Imanishi, Y.; Gerke, V.; Palczewski, K. Retinosomes: New insights into intracellular managing of hydrophobic substances in lipid bodies. J. Cell Biol. 2004, 166, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Golczak, M.; Imanishi, Y.; Kuksa, V.; Maeda, T.; Kubota, R.; Palczewski, K. Lecithin:retinol acyltransferase is responsible for amidation of retinylamine, a potent inhibitor of the retinoid cycle. J. Biol. Chem. 2005, 280, 42263–42273. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Schwarz, C.; Hunter, J.J.; Palczewska, G.; Palczewski, K.; Williams, D.R. Formation and Clearance of All-Trans-Retinol in Rods Investigated in the Living Primate Eye with Two-Photon Ophthalmoscopy. Investig. Ophthalmol. Vis. Sci. 2017, 58, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kiser, P.D.; Badiee, M.; Palczewska, G.; Dong, Z.; Golczak, M.; Tochtrop, G.P.; Palczewski, K. Molecular pharmacodynamics of emixustat in protection against retinal degeneration. J. Clin. Investig. 2015, 125, 2781–2794. [Google Scholar] [CrossRef] [PubMed]
- Kiser, P.D.; Palczewski, K. Retinoids and Retinal Diseases. Annu. Rev. Vis. Sci. 2016, 2, 197–234. [Google Scholar] [CrossRef]
- Fereidouni, F.; Bader, A.N.; Gerritsen, H.C. Spectral phasor analysis allows rapid and reliable unmixing of fluorescence microscopy spectral images. Opt. Express 2012, 20, 12729–12741. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaushik, V.; Gessa, L.; Kumar, N.; Fernandes, H. Towards a New Biomarker for Diabetic Retinopathy: Exploring RBP3 Structure and Retinoids Binding for Functional Imaging of Eyes In Vivo. Int. J. Mol. Sci. 2023, 24, 4408. https://doi.org/10.3390/ijms24054408
Kaushik V, Gessa L, Kumar N, Fernandes H. Towards a New Biomarker for Diabetic Retinopathy: Exploring RBP3 Structure and Retinoids Binding for Functional Imaging of Eyes In Vivo. International Journal of Molecular Sciences. 2023; 24(5):4408. https://doi.org/10.3390/ijms24054408
Chicago/Turabian StyleKaushik, Vineeta, Luca Gessa, Nelam Kumar, and Humberto Fernandes. 2023. "Towards a New Biomarker for Diabetic Retinopathy: Exploring RBP3 Structure and Retinoids Binding for Functional Imaging of Eyes In Vivo" International Journal of Molecular Sciences 24, no. 5: 4408. https://doi.org/10.3390/ijms24054408
APA StyleKaushik, V., Gessa, L., Kumar, N., & Fernandes, H. (2023). Towards a New Biomarker for Diabetic Retinopathy: Exploring RBP3 Structure and Retinoids Binding for Functional Imaging of Eyes In Vivo. International Journal of Molecular Sciences, 24(5), 4408. https://doi.org/10.3390/ijms24054408