
Vitamin D Increases Irisin Serum Levels and the Expression of Its Precursor in Skeletal Muscle




 ,
,
Abstract
1. Introduction
2. Results
2.1. Patients and Data Collection
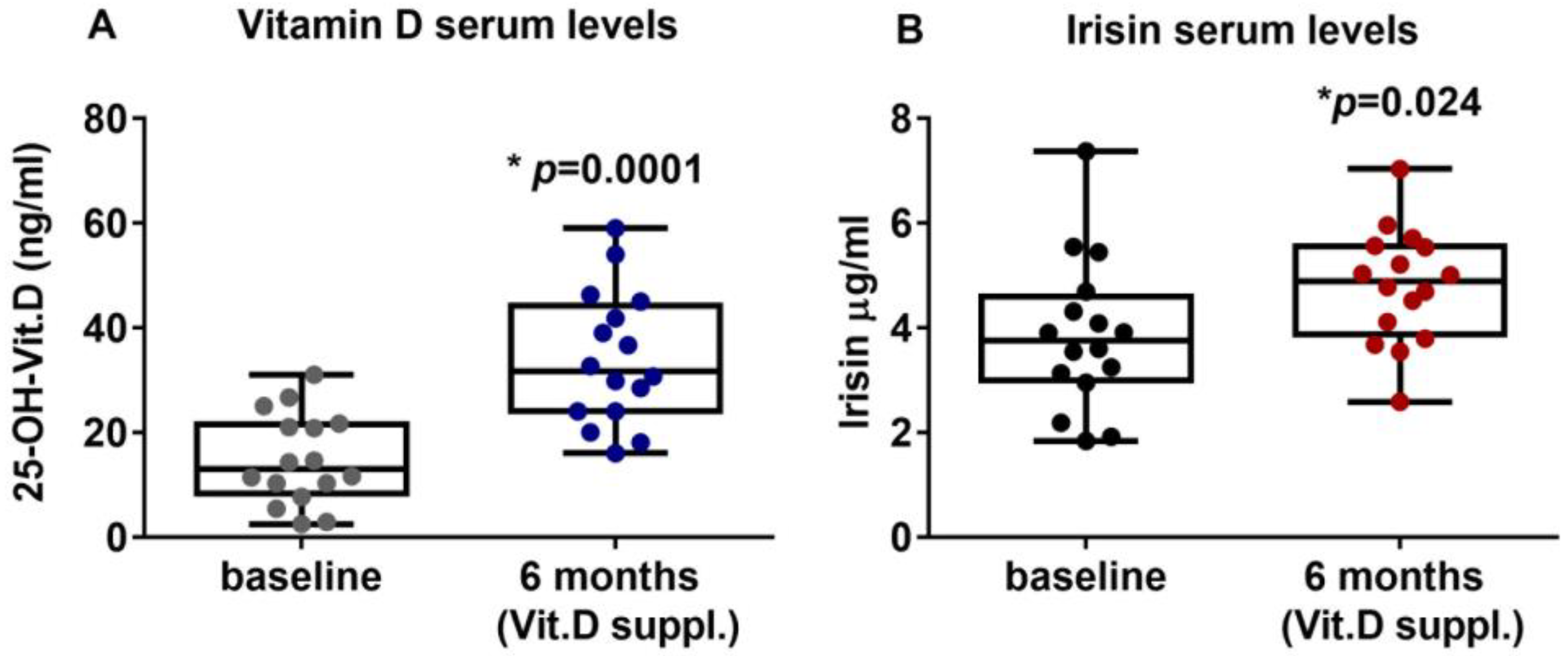
2.2. Vitamin D Supplementation Increases Irisin Serum Levels in PHPT Patients
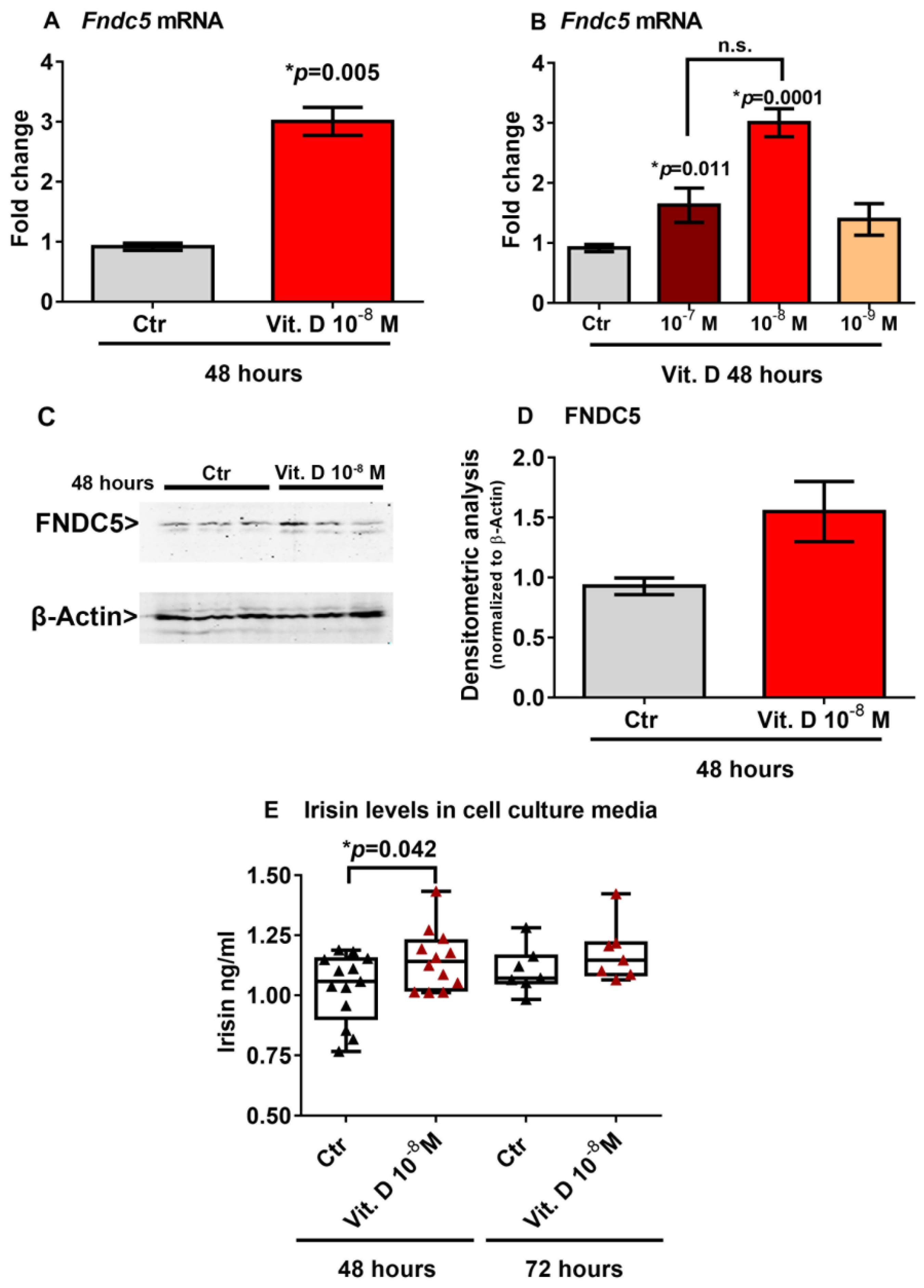
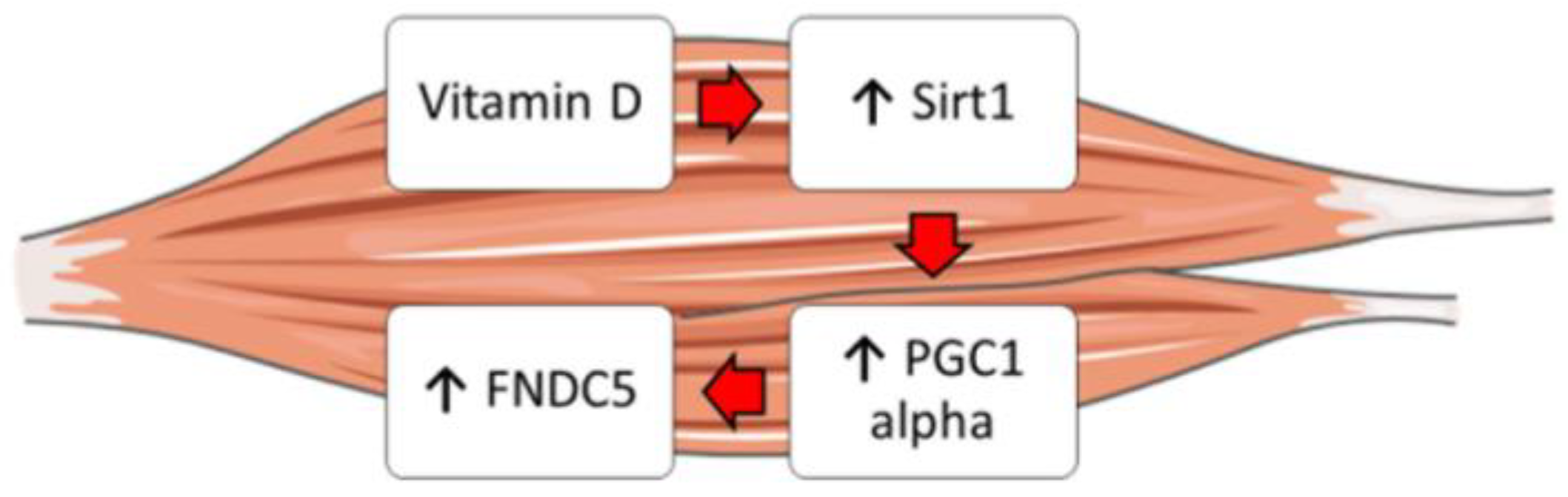
2.3. Vitamin D Increases Fndc5 Expression in Myoblasts
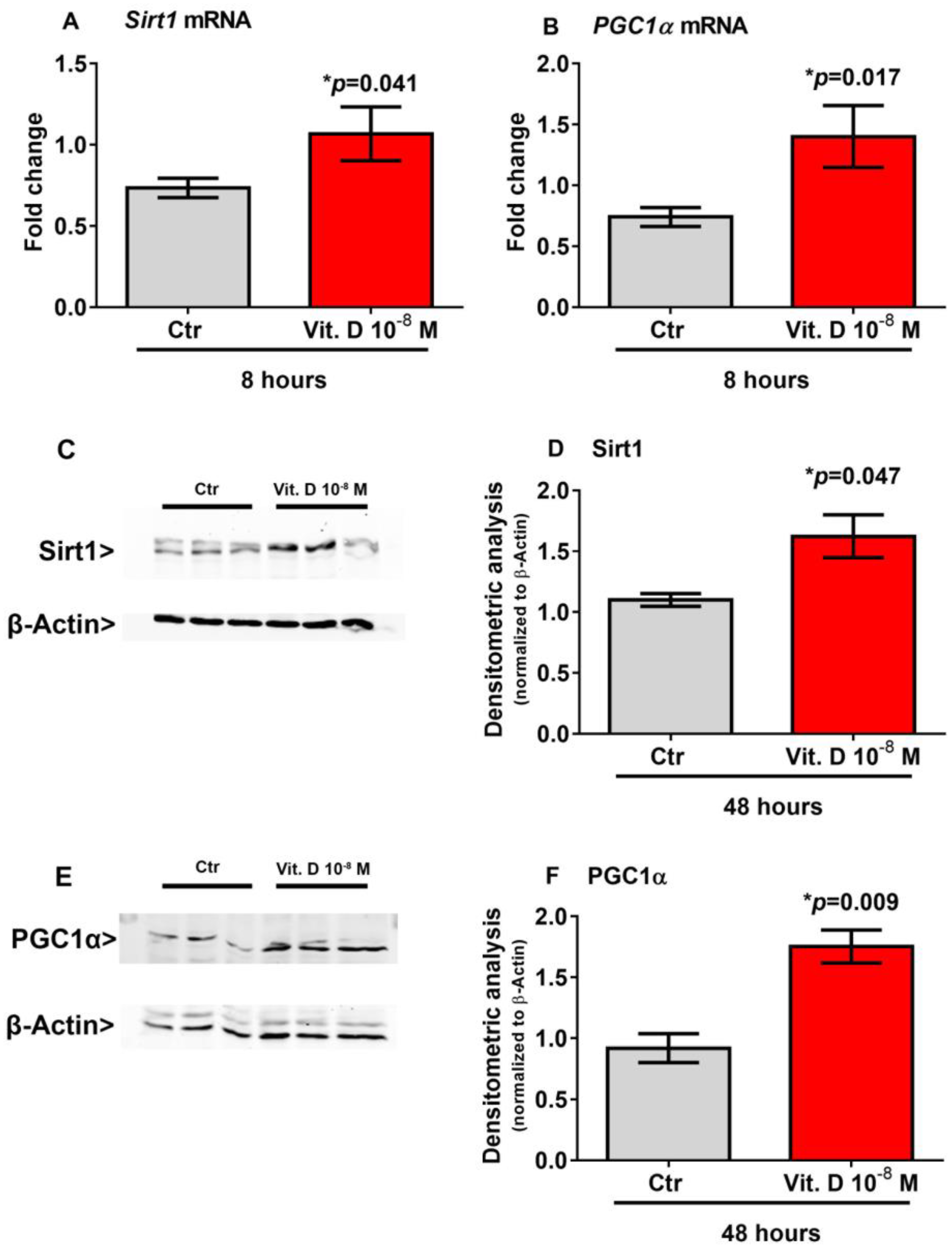
2.4. Vitamin D Increase the Expression of Sirt 1 and Pgc1α
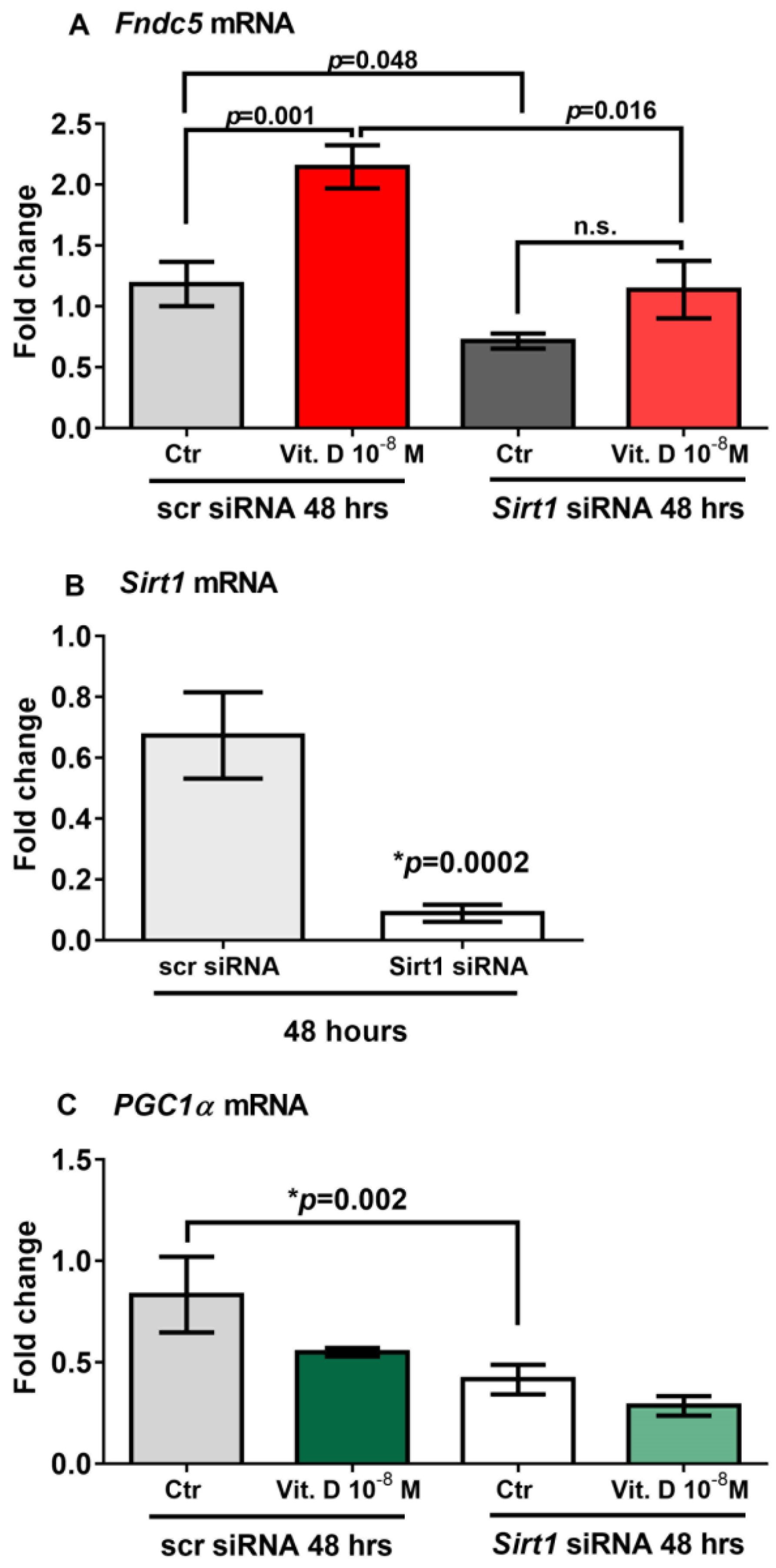
2.5. Sirt1 Is Involved in Vitamin-D-Mediated Upregulation of Fndc5
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Biochemical Measurement
4.3. Cell Culture
4.4. Vitamin D Treatment
4.5. Real Time PCR
4.6. Western Blot
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Colaianni, G.; Storlino, G.; Sanesi, L.; Colucci, S.; Grano, M. Myokines and Osteokines in the Pathogenesis of Muscle and Bone Diseases. Curr. Osteoporos. Rep. 2020, 18, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-Dependent Myokine That Drives Brown-Fat-Like Development of White Fat and Thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Colaianni, G.; Sanesi, L.; Storlino, G.; Brunetti, G.; Colucci, S.; Grano, M. Irisin and Bone: From Preclinical Studies to the Evaluation of Its Circulating Levels in Different Populations of Human Subjects. Cells 2019, 8, 451. [Google Scholar] [CrossRef] [PubMed]
- Zerlotin, R.; Oranger, A.; Pignataro, P.; Dicarlo, M.; Maselli, F.; Mori, G.; Colucci, S.C.; Grano, M.; Colaianni, G. Irisin and Secondary Osteoporosis in Humans. Int. J. Mol. Sci. 2022, 23, 690. [Google Scholar] [CrossRef]
- Colaianni, G.; Cuscito, C.; Mongelli, T.; Pignataro, P.; Buccoliero, C.; Liu, P.; Lu, P.; Sartini, L.; Di Comite, M.; Mori, G.; et al. The Myokine Irisin Increases Cortical Bone Mass. Proc. Natl. Acad. Sci. USA 2015, 112, 12157–12162. [Google Scholar] [CrossRef]
- Colaianni, G.; Mongelli, T.; Cuscito, C.; Pignataro, P.; Lippo, L.; Spiro, G.; Notarnicola, A.; Severi, I.; Passeri, G.; Mori, G.; et al. Irisin Prevents and Restores Bone Loss and Muscle Atrophy in Hind-Limb Suspended Mice. Sci. Rep. 2017, 7, 2811. [Google Scholar] [CrossRef]
- Colaianni, G.; Oranger, A.; Dicarlo, M.; Lovero, R.; Storlino, G.; Pignataro, P.; Fontana, A.; Di Serio, F.; Ingravallo, A.; Caputo, G.; et al. Irisin Serum Levels and Skeletal Muscle Assessment in a Cohort of Charcot-Marie-Tooth Patients. Front. Endocrinol. 2022, 13, 886243. [Google Scholar] [CrossRef]
- Faienza, M.F.; Brunetti, G.; Sanesi, L.; Colaianni, G.; Celi, M.; Piacente, L.; D’Amato, G.; Schipani, E.; Colucci, S.; Grano, M. High Irisin Levels Are Associated with Better Glycemic Control and Bone Health in Children With Type 1 diabetes. Diabetes Res. Clin. Pract. 2018, 141, 10–17. [Google Scholar] [CrossRef]
- Faienza, M.F.; Brunetti, G.; Grugni, G.; Fintini, D.; Convertino, A.; Pignataro, P.; Crinò, A.; Colucci, S.; Grano, M. The Genetic Background and Vitamin D Supplementation Can Affect Irisin Levels in Prader-Willi Syndrome. J. Endocrinol. Investig. 2021, 44, 2261–2271. [Google Scholar] [CrossRef]
- Safarpour, P.; Daneshi-Maskooni, M.; Vafa, M.; Nourbakhsh, M.; Janani, L.; Maddah, M.; Amiri, F.S.; Mohammadi, F.; Sadeghi, H. Vitamin D Supplementation Improves SIRT1, Irisin, and Glucose Indices in Overweight or Obese Type 2 Diabetic Patients: A Double-Blind Randomized Placebo-Controlled Clinical Trial. BMC Fam. Pract. 2020, 21, 26. [Google Scholar] [CrossRef]
- Colaianni, G.; Lippo, L.; Sanesi, L.; Brunetti, G.; Celi, M.; Cirulli, N.; Passeri, G.; Reseland, J.; Schipani, E.; Faienza, M.F.; et al. Deletion of the Transcription Factor PGC-1α in Mice Negatively Regulates Bone Mass. Calcif. Tissue Int. 2018, 103, 638–652. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, T.; Hosoyama, T.; Tomida, M.; Yamamoto, Y.; Nakamichi, Y.; Kato, S.; Kawai-Takaishi, M.; Ishizuka, S.; Nishita, Y.; Tange, C.; et al. Influence of Vitamin D on Sarcopenia Pathophysiology: A Longitudinal Study in Humans and Basic Research in Knockout Mice. J. Cachexia Sarcopenia Muscle 2022, 13, 2961–2973. [Google Scholar] [CrossRef]
- Muresan, G.C.; Hedesiu, M.; Lucaciu, O.; Boca, S.; Petrescu, N. Effect of Vitamin D on Bone Regeneration: A Review. Medicina 2022, 58, 1337. [Google Scholar] [CrossRef]
- Souberbielle, J.C.; Bienaimé, F.; Cavalier, E.; Cormier, C. Vitamin D and Primary Hyperparathyroidism (PHPT). Ann. D’endocrinologie 2012, 73, 165–169. [Google Scholar] [CrossRef]
- Moosgaard, B.; Vestergaard, P.; Heickendorff, L.; Melsen, F.; Christiansen, P.; Mosekilde, L. Vitamin D Status, Seasonal Variations, Parathyroid Adenoma Weight and Bone Mineral Density in Primary Hyperparathyroidism. Clin. Endocrinol. 2005, 63, 506–513. [Google Scholar] [CrossRef]
- Minisola, S.; Romagnoli, E.; Scillitani, A.; Rao, S.D. Hypovitaminosis D in Primary Hyperparathyroidism: To Treat or Not to Treat? That Is the Question. J. Endocrinol. Investig. 2014, 37, 413–414. [Google Scholar] [CrossRef]
- Scillitani, A.; Guarnieri, V.; Battista, C.; De Geronimo, S.; Muscarella, L.A.; Chiodini, I.; Cignarelli, M.; Minisola, S.; Bertoldo, F.; Francucci, C.M.; et al. Primary Hyperparathyroidism and the Presence Of Kidney Stones Are Associated with Different Haplotypes of the Calcium-Sensing Receptor. J. Clin. Endocrinol. Metab. 2007, 92, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, J.T.; Lerin, C.; Haas, W.; Gygi, S.P.; Spiegelman, B.M.; Puigserver, P. Nutrient Control of Glucose Homeostasis through a Complex of PGC-1alpha and SIRT1. Nature 2005, 434, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, S.; Fergusson, M.M.; Finkel, T. SIRT1 Functionally Interacts with the Metabolic Regulator and Transcriptional Coactivator PGC-1{alpha}. J. Biol. Chem. 2005, 280, 16456–16460. [Google Scholar] [CrossRef]
- Abulmeaty, M.M.A.; Almajwal, A.M.; Alam, I.; Razak, S.; ElSadek, M.F.; Aljuraiban, G.S.; Hussein, K.S.; Malash, A.M. Relationship of Vitamin D-Deficient Diet and Irisin, and Their Impact on Energy Homeostasis in Rats. Front. Physiol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Nadimi, H.; Djazayery, A.; Javanbakht, M.H.; Dehpour, A.; Ghaedi, E.; Derakhshanian, H.; Mohammadi, H.; Zarei, M.; Djalali, M. The Effect of Vitamin D Supplementation on Serum and Muscle Irisin Levels, and FNDC5 Expression in Diabetic Rats. Rep. Biochem. Mol. Biol. 2019, 8, 236–243. [Google Scholar]
- Chang, E.; Kim, Y. Vitamin D Ameliorates Fat Accumulation with AMPK/SIRT1 Activity in C2C12 Skeletal Muscle Cells. Nutrients 2019, 11, 2806. [Google Scholar] [CrossRef]
- Cantó, C.; Jiang, L.Q.; Deshmukh, A.S.; Mataki, C.; Coste, A.; Lagouge, M.; Zierath, J.R.; Auwerx, J. Interdependence of AMPK and SIRT1 for Metabolic Adaptation to Fasting and Exercise in Skeletal Muscle. Cell Metab. 2010, 11, 213–219. [Google Scholar] [CrossRef]
- Price, N.L.; Gomes, A.P.; Ling, A.J.; Duarte, F.V.; Martin-Montalvo, A.; North, B.J.; Agarwal, B.; Ye, L.; Ramadori, G.; Teodoro, J.S.; et al. SIRT1 Is Required for AMPK Activation and the Beneficial Effects of Resveratrol on Mitochondrial Function. Cell Metab. 2012, 15, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, B.M. Transcriptional Control of Mitochondrial Energy Metabolism through the PGC1 Coactivators. Novartis Found. Symp. 2007, 287, 60–63; discussion 63–69. [Google Scholar] [PubMed]
- Cannon, B.; Nedergaard, J. Brown Adipose Tissue: Function and Physiological Significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef] [PubMed]
- Braga, M.; Simmons, Z.; Norris, K.C.; Ferrini, M.G.; Artaza, J.N. Vitamin D Induces Myogenic Differentiation in Skeletal Muscle Derived Stem Cells. Endocr. Connect. 2017, 6, 139–150. [Google Scholar] [CrossRef]
- Reza, M.M.; Subramaniyam, N.; Sim, C.M.; Ge, X.; Sathiakumar, D.; McFarlane, C.; Sharma, M.; Kambadur, R. Irisin Is a Pro-Myogenic Factor That Induces Skeletal Muscle Hypertrophy and Rescues Denervation-Induced Atrophy. Nat. Commun. 2017, 8, 1104. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All Patients (n = 16) | |
|---|---|
| Age (years) | 58.69 ± 7.54 |
| BMI (kg/m2) | 26.05 (24.13–30.95) |
| Ca (mg/dL) | 9.76 ± 0.55 |
| PTH (pg/mL) | 116 (76.75–134.5) |
| 25(OH)-Vit D (ng/mL) | 14.82 ± 8.66 |
| irisin (mg/mL) | 3.85 ± 1.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanesi, L.; Dicarlo, M.; Pignataro, P.; Zerlotin, R.; Pugliese, F.; Columbu, C.; Carnevale, V.; Tunnera, S.; Scillitani, A.; Grano, M.; et al. Vitamin D Increases Irisin Serum Levels and the Expression of Its Precursor in Skeletal Muscle. Int. J. Mol. Sci. 2023, 24, 4129. https://doi.org/10.3390/ijms24044129
Sanesi L, Dicarlo M, Pignataro P, Zerlotin R, Pugliese F, Columbu C, Carnevale V, Tunnera S, Scillitani A, Grano M, et al. Vitamin D Increases Irisin Serum Levels and the Expression of Its Precursor in Skeletal Muscle. International Journal of Molecular Sciences. 2023; 24(4):4129. https://doi.org/10.3390/ijms24044129
Chicago/Turabian StyleSanesi, Lorenzo, Manuela Dicarlo, Patrizia Pignataro, Roberta Zerlotin, Flavia Pugliese, Carla Columbu, Vincenzo Carnevale, Silvia Tunnera, Alfredo Scillitani, Maria Grano, and et al. 2023. "Vitamin D Increases Irisin Serum Levels and the Expression of Its Precursor in Skeletal Muscle" International Journal of Molecular Sciences 24, no. 4: 4129. https://doi.org/10.3390/ijms24044129
APA StyleSanesi, L., Dicarlo, M., Pignataro, P., Zerlotin, R., Pugliese, F., Columbu, C., Carnevale, V., Tunnera, S., Scillitani, A., Grano, M., Colaianni, G., & Colucci, S. (2023). Vitamin D Increases Irisin Serum Levels and the Expression of Its Precursor in Skeletal Muscle. International Journal of Molecular Sciences, 24(4), 4129. https://doi.org/10.3390/ijms24044129