
Phenotypic and Genetic Links between Body Fat Measurements and Primary Open-Angle Glaucoma


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
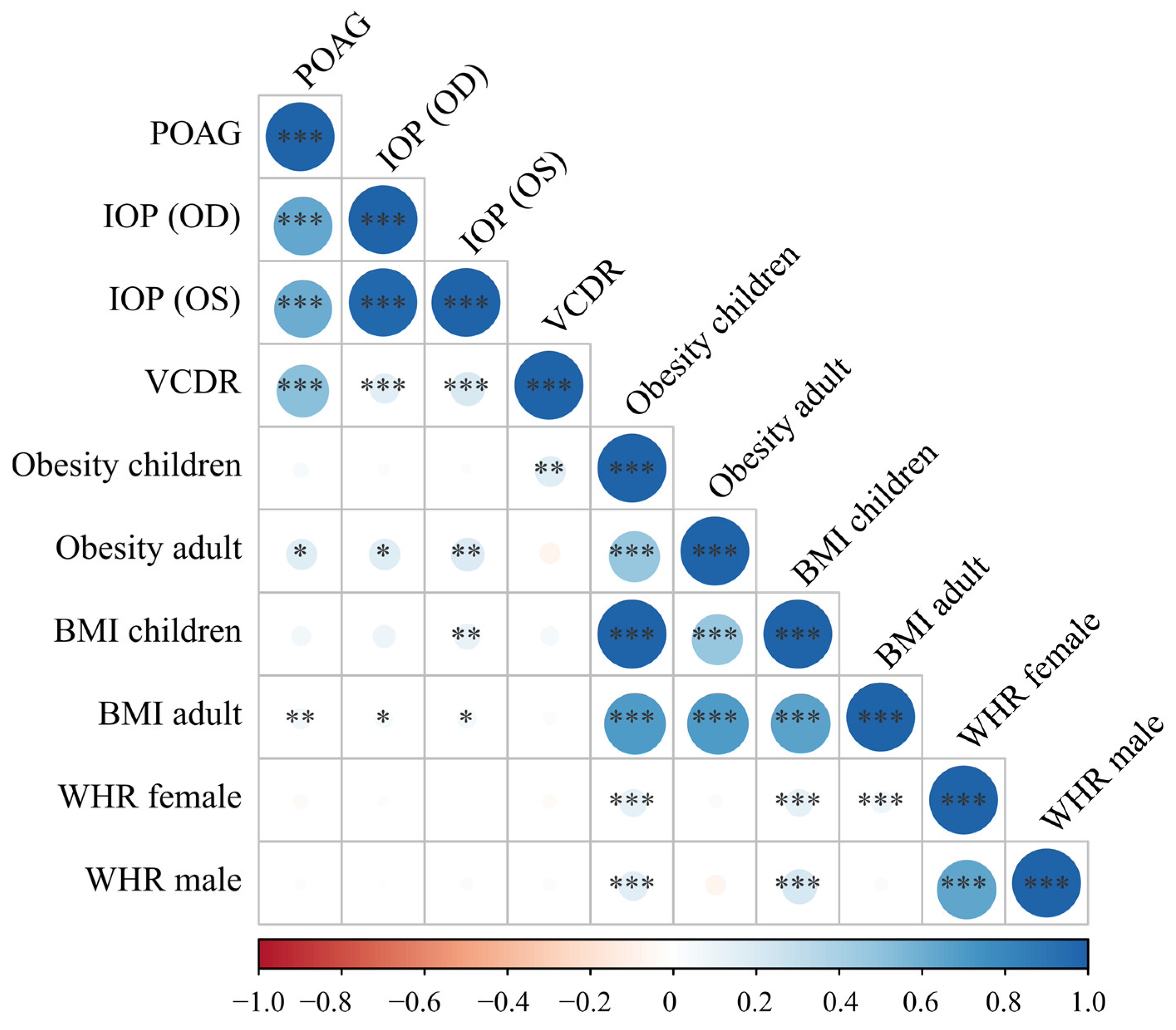
2. Results
2.1. Both Obesity and Underweight Increases POAG Risk in Longitudinal Epidemiology Studies
2.2. Genome-Wide Association Studies
2.3. Genetic Correlation
2.4. Shared Genetic Factors between Body Fat and POAG
2.4.1. Conditional Q-Q Plot
2.4.2. Shared Genetic Loci between POAG/IOP and BMI
2.4.3. Functional Annotations to the Identified Genomic Loci
3. Discussion
4. Materials and Methods
4.1. Meta-Analysis
4.2. GWAS Quality Control and Imputation
4.3. Genetic Correlation Analysis Using Linkage Disequilibrium Score Regressions
4.4. Pleiotropy Analysis
4.4.1. Conditional Quantile-Quantile Plots
4.4.2. Conditional and Conjunction False Discovery Rates
4.5. Functional Annotation of Shared Loci and Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Newcastle–Ottawa Quality Assessment Scale for Cohort Studies
- Selection
- (1)
- Representativeness of the exposed cohort
- (a)
- Truly representative of the average _______________ (describe) in the community *
- (b)
- Somewhat representative of the average ______________ in the community *
- (c)
- Selected group of users (e.g., nurses, volunteers)
- (d)
- No description of the derivation of the cohort
- (2)
- Selection of the nonexposed cohort
- (a)
- Drawn from the same community as the exposed cohort *
- (b)
- Drawn from a different source
- (c)
- No description of the derivation of the non exposed cohort
- (3)
- Ascertainment of exposure
- (a)
- Secure record (e.g., surgical records) *
- (b)
- Structured interview *
- (c)
- Written self-report
- (d)
- No description
- (4)
- Demonstration that outcome of interest was not present at start of study
- (a)
- Yes *
- (b)
- No
- Comparability
- (1)
- Comparability of cohorts on the basis of the design or analysis
- (a)
- Study controls for _____________ (select the most important factor) *
- (b)
- Study controls for any additional factor * (these criteria could be modified to indicate specific control for a second important factor)
- Outcome
- (1)
- Assessment of outcome
- (a)
- Independent blind assessment *
- (b)
- Record linkage *
- (c)
- Self-report
- (d)
- No description
- (2)
- Was follow-up long enough for outcomes to occur
- (a)
- Yes (select an adequate follow-up period for outcome of interest) *
- (b)
- No
- (3)
- Adequacy of follow-up of cohorts
- (a)
- Complete follow-up—all subjects accounted for *
- (b)
- Subjects lost to follow-up unlikely to introduce bias—small number lost—>____% (select an adequate %) follow-up, or description provided of those lost) *
- (c)
- Follow-up rate <____% (select an adequate %) and no description of those lost
- (d)
- No statement
References
- WHO. Obesity and Overweight. Fact Sheet. Available online: https://www.who.int/en/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 19 December 2022).
- Garvey, W.T.; Mechanick, J.I.; Brett, E.M.; Garber, A.J.; Hurley, D.L.; Jastreboff, A.M.; Nadolsky, K.; Pessah-Pollack, R.; Plodkowski, R. American Association of Clinical Endocrinologists and American College of Endocrinology Comprehensive Clinical Practice Guidelines for Medical Care of Patients with Obesity. Endocr. Pract. 2016, 22 (Suppl. S3), 1–203. [Google Scholar] [CrossRef] [PubMed]
- Yuen, M.; Lui, D.; Kaplan, L. A Systematic Review and Evaluation of Current Evidence Reveals 195 Obesity-Associated Disorders (OBAD). In Proceedings of the Obesity Week 2016, New Orleans, LA, USA, 31 October–4 November 2016. Poster T-P-3166. [Google Scholar]
- Jung, Y.; Han, K.; Park, H.Y.L.; Lee, S.H.; Park, C.K. Metabolic Health, Obesity, and the Risk of Developing Open-Angle Glaucoma: Metabolically Healthy Obese Patients versus Metabolically Unhealthy but Normal Weight Patients. Diabetes Metab. J. 2020, 44, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Choi, H.J.; Jeoung, J.W.; Park, K.H.; Kim, D.M. Five-year incidence of primary open-angle glaucoma and rate of progression in health center-based Korean population: The Gangnam Eye Study. PLoS ONE 2014, 9, e114058. [Google Scholar] [CrossRef] [PubMed]
- Marshall, H.; Berry, E.C.; Torres, S.D.; Mullany, S.; Schmidt, J.; Thomson, D.; Nguyen, T.T.; Knight, L.S.; Hollitt, G.; Qassim, A.; et al. Association Between Body Mass Index and Primary Open Angle Glaucoma in Three Cohorts. Am. J. Ophthalmol. 2022, 245, 126–133. [Google Scholar] [CrossRef]
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef]
- WHO. Blindness and Vision Impairment. Available online: https://www.who.int/news-room/fact-sheets/detail/blindness-and-visual-impairment (accessed on 19 December 2022).
- Jonas, J.B.; Aung, T.; Bourne, R.R.; Bron, A.M.; Ritch, R.; Panda-Jonas, S. Glaucoma. Lancet 2017, 390, 2183–2193. [Google Scholar] [CrossRef]
- Chen, W.D.; Lai, L.J.; Lee, K.L.; Chen, T.J.; Liu, C.Y.; Yang, Y.H. Is Obesity a Risk or Protective Factor for Open-Angle Glaucoma in Adults? A Two-Database, Asian, Matched-Cohort Study. J. Clin. Med. 2021, 10, 4021. [Google Scholar] [CrossRef]
- Newman-Casey, P.A.; Talwar, N.; Nan, B.; Musch, D.C.; Stein, J.D. The relationship between components of metabolic syndrome and open-angle glaucoma. Ophthalmology 2011, 118, 1318–1326. [Google Scholar] [CrossRef]
- Burgansky-Eliash, Z.; Achiron, A.; Hecht, I.; Shimonov, M. Reduction of intraocular pressure after bariatric surgery. Acta Ophthalmol. 2018, 96, e592–e595. [Google Scholar] [CrossRef]
- Viljanen, A.; Hannukainen, J.C.; Soinio, M.; Karlsson, H.K.; Salminen, P.; Nuutila, P.; Vesti, E. The effect of bariatric surgery on intraocular pressure. Acta Ophthalmol. 2018, 96, 849–852. [Google Scholar] [CrossRef]
- Na, K.S.; Kim, J.H.; Paik, J.S.; Cho, W.K.; Ha, M.; Park, Y.G.; Yang, S.W. Underweight increases the risk of primary open-angle glaucoma in diabetes patients: A Korean nationwide cohort study. Medicine 2020, 99, e19285. [Google Scholar] [CrossRef] [PubMed]
- Pasquale, L.R.; Willett, W.C.; Rosner, B.A.; Kang, J.H. Anthropometric measures and their relation to incident primary open-angle glaucoma. Ophthalmology 2010, 117, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Varma, R.; Wu, S.; Torres, M.; Azen, S.P.; Francis, B.A.; Chopra, V.; Nguyen, B.B. Baseline risk factors that predict the development of open-angle glaucoma in a population: The Los Angeles Latino Eye Study. Ophthalmology 2012, 119, 2245–2253. [Google Scholar] [CrossRef] [PubMed]
- Ramdas, W.D.; Wolfs, R.C.; Hofman, A.; de Jong, P.T.; Vingerling, J.R.; Jansonius, N.M. Lifestyle and risk of developing open-angle glaucoma: The Rotterdam study. Arch. Ophthalmol. 2011, 129, 767–772. [Google Scholar] [CrossRef]
- Herrera, B.M.; Lindgren, C.M. The genetics of obesity. Curr. Diab. Rep. 2010, 10, 498–505. [Google Scholar] [CrossRef]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef]
- Fall, T.; Ingelsson, E. Genome-wide association studies of obesity and metabolic syndrome. Mol. Cell. Endocrinol. 2014, 382, 740–757. [Google Scholar] [CrossRef]
- Wiggs, J.L.; Pasquale, L.R. Genetics of glaucoma. Hum. Mol. Genet. 2017, 26, R21–R27. [Google Scholar] [CrossRef]
- Aboobakar, I.F.; Wiggs, J.L. The genetics of glaucoma: Disease associations, personalised risk assessment and therapeutic opportunities-A review. Clin. Exp. Ophthalmol. 2022, 50, 143–162. [Google Scholar] [CrossRef]
- Wang, Z.; Wiggs, J.L.; Aung, T.; Khawaja, A.P.; Khor, C.C. The genetic basis for adult onset glaucoma: Recent advances and future directions. Prog. Retin. Eye Res. 2022, 90, 101066. [Google Scholar] [CrossRef]
- Lin, Y.; Zhu, X.; Luo, W.; Jiang, B.; Lin, Q.; Tang, M.; Li, X.; Xie, L. The Causal Association Between Obesity and Primary Open-Angle Glaucoma: A Two-Sample Mendelian Randomization Study. Front. Genet. 2022, 13, 835524. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R.; Liu, K.; Cai, Y.; He, F.; Xiao, X.; Zou, J. Body shape and risk of glaucoma: A Mendelian randomization. Front. Med. 2022, 9, 999974. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yu, K.F. What’s the relative risk? A method of correcting the odds ratio in cohort studies of common outcomes. JAMA 1998, 280, 1690–1691. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.M.; Reid, J.E. The hazard ratio is interpretable as an odds or a probability under the assumption of proportional hazards. arXiv 2021, arXiv:2109.12207. [Google Scholar]
- Gharahkhani, P.; Jorgenson, E.; Hysi, P.; Khawaja, A.P.; Pendergrass, S.; Han, X.; Ong, J.S.; Hewitt, A.W.; Segre, A.V.; Rouhana, J.M.; et al. Genome-wide meta-analysis identifies 127 open-angle glaucoma loci with consistent effect across ancestries. Nat. Commun. 2021, 12, 1258. [Google Scholar] [CrossRef]
- Pan-UKB_Team. Pan-UK Biobank. Available online: https://pan.ukbb.broadinstitute.org/ (accessed on 1 December 2022).
- Springelkamp, H.; Iglesias, A.I.; Mishra, A.; Hohn, R.; Wojciechowski, R.; Khawaja, A.P.; Nag, A.; Wang, Y.X.; Wang, J.J.; Cuellar-Partida, G.; et al. New insights into the genetics of primary open-angle glaucoma based on meta-analyses of intraocular pressure and optic disc characteristics. Hum. Mol. Genet. 2017, 26, 438–453. [Google Scholar] [CrossRef]
- Bradfield, J.P.; Taal, H.R.; Timpson, N.J.; Scherag, A.; Lecoeur, C.; Warrington, N.M.; Hypponen, E.; Holst, C.; Valcarcel, B.; Thiering, E.; et al. A genome-wide association meta-analysis identifies new childhood obesity loci. Nat. Genet. 2012, 44, 526–531. [Google Scholar] [CrossRef]
- Jiang, L.; Zheng, Z.; Fang, H.; Yang, J. A generalized linear mixed model association tool for biobank-scale data. Nat. Genet. 2021, 53, 1616–1621. [Google Scholar] [CrossRef]
- Yengo, L.; Sidorenko, J.; Kemper, K.E.; Zheng, Z.; Wood, A.R.; Weedon, M.N.; Frayling, T.M.; Hirschhorn, J.; Yang, J.; Visscher, P.M.; et al. Meta-analysis of genome-wide association studies for height and body mass index in approximately 700000 individuals of European ancestry. Hum. Mol. Genet. 2018, 27, 3641–3649. [Google Scholar] [CrossRef]
- Vogelezang, S.; Bradfield, J.P.; Ahluwalia, T.S.; Curtin, J.A.; Lakka, T.A.; Grarup, N.; Scholz, M.; van der Most, P.J.; Monnereau, C.; Stergiakouli, E.; et al. Novel loci for childhood body mass index and shared heritability with adult cardiometabolic traits. PLoS Genet. 2020, 16, e1008718. [Google Scholar] [CrossRef]
- Pulit, S.L.; Stoneman, C.; Morris, A.P.; Wood, A.R.; Glastonbury, C.A.; Tyrrell, J.; Yengo, L.; Ferreira, T.; Marouli, E.; Ji, Y.; et al. Meta-analysis of genome-wide association studies for body fat distribution in 694 649 individuals of European ancestry. Hum. Mol. Genet. 2019, 28, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Auburger, G.; Sen, N.E.; Meierhofer, D.; Basak, A.N.; Gitler, A.D. Efficient Prevention of Neurodegenerative Diseases by Depletion of Starvation Response Factor Ataxin-2. Trends Neurosci. 2017, 40, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Ju, W.K.; Perkins, G.A.; Kim, K.Y.; Bastola, T.; Choi, W.Y.; Choi, S.H. Glaucomatous optic neuropathy: Mitochondrial dynamics, dysfunction and protection in retinal ganglion cells. Prog. Retin. Eye Res. 2022, 84, 101136. [Google Scholar] [CrossRef] [PubMed]
- Meierhofer, D.; Halbach, M.; Sen, N.E.; Gispert, S.; Auburger, G. Ataxin-2 (Atxn2)-Knock-Out Mice Show Branched Chain Amino Acids and Fatty Acids Pathway Alterations. Mol. Cell. Proteom. MCP 2016, 15, 1728–1739. [Google Scholar] [CrossRef] [PubMed]
- Darbandi, M.; Pasdar, Y.; Moradi, S.; Mohamed, H.J.J.; Hamzeh, B.; Salimi, Y. Discriminatory Capacity of Anthropometric Indices for Cardiovascular Disease in Adults: A Systematic Review and Meta-Analysis. Prev. Chronic Dis. 2020, 17, E131. [Google Scholar] [CrossRef] [PubMed]
- Bennett, W.L.; Wilson, R.F.; Zhang, A.; Tseng, E.; Knapp, E.A.; Kharrazi, H.; Stuart, E.A.; Shogbesan, O.; Bass, E.B.; Cheskin, L.J. Methods for Evaluating Natural Experiments in Obesity: Systematic Evidence Review. Comp. Eff. Rev. 2018, 168, 791–800. [Google Scholar] [CrossRef]
- Kmet, L.M.; Lee, R.C.; Cook, L.S. Standard Quality Assessment Criteria for Evaluating Primary Research Papers from a Variety of Fields; Alberta Heritage Foundation for Medical Research: Edmonton, AB, Canada, 2004; p. 22. [Google Scholar]
- Khan, K.S.; Riet, G.t.; Popay, J.; Nixon, J.; Kleijnen, J. Conducting the review: Phase 5 study quality assessment. In Undertaking Systematic Reviews of Research Effectiveness CDC’s Guidance for Those Carrying Out or Commissioning Reviews; Centre for Reviews and Dissemination (CRD) at the University of York: York, UK, 2001; p. 20. [Google Scholar]
- Rong, S.S.; Lee, B.Y.; Kuk, A.K.; Yu, X.T.; Li, S.S.; Li, J.; Guo, Y.; Yin, Y.; Osterbur, D.L.; Yam, J.C.S.; et al. Comorbidity of dementia and age-related macular degeneration calls for clinical awareness: A meta-analysis. Br. J. Ophthalmol. 2019, 103. [Google Scholar] [CrossRef]
- Rong, S.S.; Tang, F.Y.; Chu, W.K.; Ma, L.; Yam, J.C.; Tang, S.M.; Li, J.; Gu, H.; Young, A.L.; Tham, C.C.; et al. Genetic Associations of Primary Angle-Closure Disease: A Systematic Review and Meta-analysis. Ophthalmology 2016, 123, 1211–1221. [Google Scholar] [CrossRef]
- Rong, S.S.; Peng, Y.; Liang, Y.B.; Cao, D.; Jhanji, V. Does cigarette smoking alter the risk of pterygium? A systematic review and meta-analysis. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6235–6243. [Google Scholar] [CrossRef]
- DerSimonian, R.; Laird, N. Meta-analysis in clinical trials. Control. Clin. Trials 1986, 7, 177–188. [Google Scholar] [CrossRef]
- Higgins, J.P.; Thompson, S.G. Quantifying heterogeneity in a meta-analysis. Stat. Med. 2002, 21, 1539–1558. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.; Thompson, S.G.; Deeks, J.J.; Altman, D.G. Measuring inconsistency in meta-analyses. BMJ 2003, 327, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.L.; Sutton, A.J.; Jones, D.R.; Abrams, K.R.; Rushton, L. Contour-enhanced meta-analysis funnel plots help distinguish publication bias from other causes of asymmetry. J. Clin. Epidemiol. 2008, 61, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.; Thomas, J.; Chandler, J.; Cumpston, M.; Li, T.; Page, M.J.; Welch, V.A. (Eds.) Cochrane Handbook for Systematic Reviews of Interventions; John Wiley & Sons Ltd.: Oxford, UK, 2019; Available online: www.cochrane-handbook.org (accessed on 20 November 2022).
- Sterne, J.A.; Gavaghan, D.; Egger, M. Publication and related bias in meta-analysis: Power of statistical tests and prevalence in the literature. J. Clin. Epidemiol. 2000, 53, 1119–1129. [Google Scholar] [CrossRef]
- Bulik-Sullivan, B.; Finucane, H.K.; Anttila, V.; Gusev, A.; Day, F.R.; Loh, P.R.; ReproGen Consortium; Psychiatric Genomics Consortium; Genetic Consortium for Anorexia Nervosa of the Wellcome Trust Case Control Consortium 3; Duncan, L.; et al. An atlas of genetic correlations across human diseases and traits. Nat. Genet. 2015, 47, 1236–1241. [Google Scholar] [CrossRef]
- Sudlow, C.; Gallacher, J.; Allen, N.; Beral, V.; Burton, P.; Danesh, J.; Downey, P.; Elliott, P.; Green, J.; Landray, M.; et al. UK biobank: An open access resource for identifying the causes of a wide range of complex diseases of middle and old age. PLoS Med. 2015, 12, e1001779. [Google Scholar] [CrossRef]
- UK Biobank. Available online: http://www.nealelab.is/uk-biobank (accessed on 2 November 2022).
- Genomes Project, C.; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef]
- Schorsch, E. LDSC (LD SCore). Available online: https://github.com/bulik/ldsc (accessed on 24 July 2022).
- Andreassen, O.A.; Thompson, W.K.; Schork, A.J.; Ripke, S.; Mattingsdal, M.; Kelsoe, J.R.; Kendler, K.S.; O’Donovan, M.C.; Rujescu, D.; Werge, T.; et al. Improved detection of common variants associated with schizophrenia and bipolar disorder using pleiotropy-informed conditional false discovery rate. PLoS Genet. 2013, 9, e1003455. [Google Scholar] [CrossRef]
- Smeland, O.B.; Frei, O.; Shadrin, A.; O’Connell, K.; Fan, C.C.; Bahrami, S.; Holland, D.; Djurovic, S.; Thompson, W.K.; Dale, A.M.; et al. Discovery of shared genomic loci using the conditional false discovery rate approach. Hum. Genet. 2020, 139, 85–94. [Google Scholar] [CrossRef]
- Desikan, R.S.; Schork, A.J.; Wang, Y.; Thompson, W.K.; Dehghan, A.; Ridker, P.M.; Chasman, D.I.; McEvoy, L.K.; Holland, D.; Chen, C.H.; et al. Polygenic Overlap Between C-Reactive Protein, Plasma Lipids, and Alzheimer Disease. Circulation 2015, 131, 2061–2069. [Google Scholar] [CrossRef]
- Devlin, B.; Roeder, K. Genomic control for association studies. Biometrics 1999, 55, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Schork, A.J.; Thompson, W.K.; Pham, P.; Torkamani, A.; Roddey, J.C.; Sullivan, P.F.; Kelsoe, J.R.; O’Donovan, M.C.; Furberg, H.; The Tobacco and Genetics Consortium; et al. All SNPs are not created equal: Genome-wide association studies reveal a consistent pattern of enrichment among functionally annotated SNPs. PLoS Genet. 2013, 9, e1003449. [Google Scholar] [CrossRef]
- Price, A.L.; Weale, M.E.; Patterson, N.; Myers, S.R.; Need, A.C.; Shianna, K.V.; Ge, D.; Rotter, J.I.; Torres, E.; Taylor, K.D.; et al. Long-range LD can confound genome scans in admixed populations. Am. J. Hum. Genet. 2008, 83, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Liley, J.; Wallace, C. A pleiotropy-informed Bayesian false discovery rate adapted to a shared control design finds new disease associations from GWAS summary statistics. PLoS Genet. 2015, 11, e1004926. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Liu, S.; Zhang, X.; Hu, Y.; Shang, X.; Zhu, Z.; Huang, Y.; Wu, G.; Xiao, Y.; Du, Z.; et al. Shared genetic architecture between the two neurodegenerative diseases: Alzheimer’s disease and glaucoma. Front. Aging Neurosci. 2022, 14, 880576. [Google Scholar] [CrossRef] [PubMed]
- Witoelar, A.; Jansen, I.E.; Wang, Y.; Desikan, R.S.; Gibbs, J.R.; Blauwendraat, C.; Thompson, W.K.; Hernandez, D.G.; Djurovic, S.; Schork, A.J.; et al. Genome-wide Pleiotropy Between Parkinson Disease and Autoimmune Diseases. JAMA Neurol. 2017, 74, 780–792. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, S.; Shadrin, A.; Frei, O.; O’Connell, K.S.; Bettella, F.; Krull, F.; Fan, C.C.; Rossberg, J.I.; Hindley, G.; Ueland, T.; et al. Genetic loci shared between major depression and intelligence with mixed directions of effect. Nat. Hum. Behav. 2021, 5, 795–801. [Google Scholar] [CrossRef]
- Watanabe, K.; Umicevic Mirkov, M.; de Leeuw, C.A.; van den Heuvel, M.P.; Posthuma, D. Genetic mapping of cell type specificity for complex traits. Nat. Commun. 2019, 10, 3222. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with the PANTHER classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rong, S.S.; Yu, X. Phenotypic and Genetic Links between Body Fat Measurements and Primary Open-Angle Glaucoma. Int. J. Mol. Sci. 2023, 24, 3925. https://doi.org/10.3390/ijms24043925
Rong SS, Yu X. Phenotypic and Genetic Links between Body Fat Measurements and Primary Open-Angle Glaucoma. International Journal of Molecular Sciences. 2023; 24(4):3925. https://doi.org/10.3390/ijms24043925
Chicago/Turabian StyleRong, Shi Song, and Xinting Yu. 2023. "Phenotypic and Genetic Links between Body Fat Measurements and Primary Open-Angle Glaucoma" International Journal of Molecular Sciences 24, no. 4: 3925. https://doi.org/10.3390/ijms24043925
APA StyleRong, S. S., & Yu, X. (2023). Phenotypic and Genetic Links between Body Fat Measurements and Primary Open-Angle Glaucoma. International Journal of Molecular Sciences, 24(4), 3925. https://doi.org/10.3390/ijms24043925