
On the Oral Microbiome of Oral Potentially Malignant and Malignant Disorders: Dysbiosis, Loss of Diversity, and Pathogens Enrichment

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Participant Characteristics and Sequencing Data Summary
2.2. Differences in Microbiome Diversity
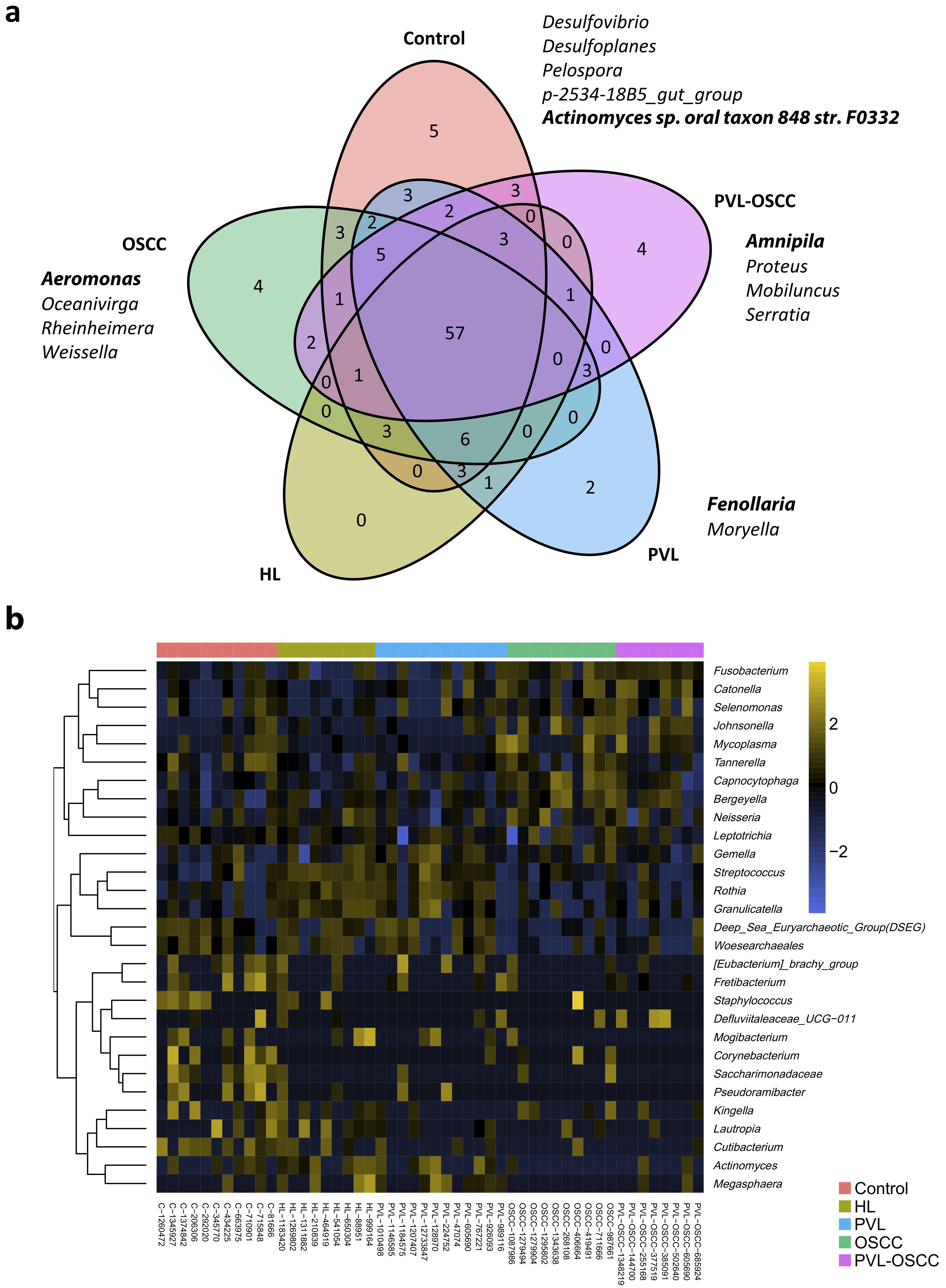
2.3. Differences in Microbiome Composition
2.4. Community Structure Reveals Differently Abundant ASVs in Oral Disorders
3. Discussion
4. Materials and Methods
4.1. Patients and Tissue Samples
4.2. DNA Extraction and 16S rDNA Gene Sequencing
4.3. Bioinformatic Analysis and Taxonomic Annotation
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Warnakulasuriya, S.; Kujan, O.; Aguirre-Urizar, J.M.; Bagan, J.V.; González-Moles, M.Á.; Kerr, A.R.; Lodi, G.; Mello, F.W.; Monteiro, L.; Ogden, G.R.; et al. Oral potentially malignant disorders: A consensus report from an international seminar on nomenclature and classification, convened by the WHO Collaborating Centre for Oral Cancer. Oral Dis. 2021, 27, 1862–1880. [Google Scholar] [CrossRef] [PubMed]
- Warnakulasuriya, S.; Johnson, N.W.; Van Der Waal, I. Nomenclature and classification of potentially malignant disorders of the oral mucosa. J. Oral Pathol. Med. 2007, 36, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Urizar, J.M.; Lafuente-Ibáñez de Mendoza, I.; Warnakulasuriya, S. Malignant transformation of oral leukoplakia: Systematic review and meta-analysis of the last 5 years. Oral Dis. 2021, 27, 1881–1895. [Google Scholar] [CrossRef] [PubMed]
- Bagan, J.; Martorell, M.; Cebrián, J.L.; Rubert, A.; Bagán, L.; Mezquida, C.; Hervás, D. Effect of clinical and histologic features on time to malignancy in 224 cases of oral leukoplakia treated by surgery. Clin. Oral Investig. 2022, 26, 5181–5188. [Google Scholar] [CrossRef]
- Ramos-García, P.; González-Moles, M.Á.; Mello, F.W.; Bagan, J.V.; Warnakulasuriya, S. Malignant transformation of oral proliferative verrucous leukoplakia: A systematic review and meta-analysis. Oral Dis. 2021, 27, 1896–1907. [Google Scholar] [CrossRef]
- Villa, A.; Menon, R.S.; Kerr, A.R.; De Abreu Alves, F.; Guollo, A.; Ojeda, D.; Woo, S.B. Proliferative leukoplakia: Proposed new clinical diagnostic criteria. Oral Dis. 2018, 24, 749–760. [Google Scholar] [CrossRef]
- Bagan, J.; Murillo-Cortes, J.; Poveda-Roda, R.; Leopoldo-Rodado, M.; Bagan, L. Second primary tumors in proliferative verrucous leukoplakia: A series of 33 cases. Clin. Oral Investig. 2020, 24, 1963–1969. [Google Scholar] [CrossRef]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Prim. 2020, 6, 92. [Google Scholar] [CrossRef]
- Chen, T.; Yu, W.-H.; Izard, J.; Baranova, O.V.; Lakshmanan, A.; Dewhirst, F.E. The Human Oral Microbiome Database: A web accessible resource for investigating oral microbe taxonomic and genomic information. Database 2010, 2010, baq013. [Google Scholar] [CrossRef]
- Ganesan, S.M.; Joshi, V.; Fellows, M.; Dabdoub, S.M.; Nagaraja, H.N.; O’Donnell, B.; Deshpande, N.R.; Kumar, P.S. A tale of two risks: Smoking, diabetes and the subgingival microbiome. ISME J. 2017, 11, 2075–2089. [Google Scholar] [CrossRef]
- Long, J.; Cai, Q.; Steinwandel, M.; Hargreaves, M.K.; Bordenstein, S.R.; Blot, W.J.; Zheng, W.; Shu, X.O. Association of oral microbiome with type 2 diabetes risk. J. Periodontal Res. 2017, 52, 636–643. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Kong, C.; Yang, Y.; Cai, S.; Li, X.; Cai, G.; Ma, Y. Human oral microbiome dysbiosis as a novel non-invasive biomarker in detection of colorectal cancer. Theranostics 2020, 10, 11595–11606. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-R.; Xu, Q.; Xiao, J.; Deng, Y.-M.; Tang, Z.-H.; Tang, Y.-L.; Liu, L.S. Role of oral microbiota in atherosclerosis. Clin. Chim. Acta 2020, 506, 191–195. [Google Scholar] [CrossRef]
- Ruff, W.E.; Greiling, T.M.; Kriegel, M.A. Host–microbiota interactions in immune-mediated diseases. Nat. Rev. Microbiol. 2020, 18, 521–538. [Google Scholar] [CrossRef] [PubMed]
- Johannsen, A.; Susin, C.; Gustafsson, A. Smoking and inflammation: Evidence for a synergistic role in chronic disease. Periodontol. 2000 2014, 64, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Bullon, P.; Newman, H.N.; Battino, M. Obesity, diabetes mellitus, atherosclerosis and chronic periodontitis: A shared pathology via oxidative stress and mitochondrial dysfunction? Periodontol. 2000 2014, 64, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Tezal, M.; Sullivan, M.A.; Hyland, A.; Marshall, J.R.; Stoler, D.; Reid, M.E.; Loree, T.R.; Rigual, N.R.; Merzianu, M.; Hauck, L.; et al. Chronic periodontitis and the incidence of head and neck squamous cell carcinoma. Cancer Epidemiol. Biomark. Prev. 2009, 18, 2406–2412. [Google Scholar] [CrossRef]
- Palefsky, J.M.; Silverman, S.J.; Abdel-Salaam, M.; Daniels, T.E.; Greenspan, J.S. Association between proliferative verrucous leukoplakia and infection with human papillomavirus type 16. J. Oral Pathol. Med. 1995, 24, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Campisi, G.; Giovannelli, L.; Ammatuna, P.; Capra, G.; Colella, G.; Di Liberto, C.; Gandolfo, S.; Pentenero, M.; Carrozzo, M.; Serpico, R.; et al. Proliferative verrucous vs conventional leukoplakia: No significantly increased risk of HPV infection. Oral Oncol. 2004, 40, 835–840. [Google Scholar] [CrossRef]
- García-López, R.; Moya, A.; Bagan, J.V.; Pérez-Brocal, V. Retrospective case-control study of viral pathogen screening in proliferative verrucous leukoplakia lesions. Clin. Otolaryngol. 2014, 39, 272–280. [Google Scholar] [CrossRef]
- Upadhyaya, J.D.; Fitzpatrick, S.G.; Islam, M.N.; Bhattacharyya, I.; Cohen, D.M. A Retrospective 20-Year Analysis of Proliferative Verrucous Leukoplakia and Its Progression to Malignancy and Association with High-risk Human Papillomavirus. Head Neck Pathol. 2018, 12, 500–510. [Google Scholar] [CrossRef]
- Herreros-Pomares, A.; Llorens, C.; Soriano, B.; Zhang, F.; Gallach, S.; Bagan, L.; Murillo, J.; Jantus-Lewintre, E.; Bagan, J. Oral microbiome in Proliferative Verrucous Leukoplakia exhibits loss of diversity and enrichment of pathogens. Oral Oncol. 2021, 120, 105404. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; van der Voort, M.; Schneider, J.H.M.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.; et al. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Gholizadeh, P.; Eslami, H.; Yousefi, M.; Asgharzadeh, M.; Aghazadeh, M.; Kafil, H.S. Role of oral microbiome on oral cancers, a review. Biomed. Pharmacother. 2016, 84, 552–558. [Google Scholar] [CrossRef]
- Michikawa, C.; Gopalakrishnan, V.; Harrandah, A.M.; Karpinets, T.V.; Garg, R.R.; Chu, R.A.; Park, Y.P.; Chukkapallia, S.S.; Yadlapalli, N.; Erikson-Carter, K.C.; et al. Fusobacterium is enriched in oral cancer and promotes induction of programmed death-ligand 1 (PD-L1). Neoplasia 2022, 31, 100813. [Google Scholar] [CrossRef]
- Xu, L.; Li, Y.; Sun, S.; Yue, J. Decrease of oral microbial diversity might correlate with radiation esophagitis in patients with esophageal cancer undergoing chemoradiation: A pilot study. Precis. Radiat. Oncol. 2020, 4, 81–88. [Google Scholar] [CrossRef]
- Debelius, J.W.; Huang, T.; Cai, Y.; Ploner, A.; Barrett, D.; Zhou, X.; Xiao, X.; Li, Y.; Liao, J.; Zheng, Y.; et al. Subspecies Niche Specialization in the Oral Microbiome Is Associated with Nasopharyngeal Carcinoma Risk. mSystems 2020, 5, e00065-20. [Google Scholar] [CrossRef] [PubMed]
- Verma, D.; Garg, P.K.; Dubey, A.K. Insights into the human oral microbiome. Arch. Microbiol. 2018, 200, 525–540. [Google Scholar] [CrossRef]
- Ismail, Y.; Mahendran, V.; Octavia, S.; Day, A.S.; Riordan, S.M.; Grimm, M.C.; Lan, R.; Lemberg, D.; Tran, T.A.; Zhang, L. Investigation of the enteric pathogenic potential of oral Campylobacter concisus strains isolated from patients with inflammatory bowel disease. PLoS ONE 2012, 7, e38217. [Google Scholar] [CrossRef]
- Poosari, A.; Nutravong, T.; Sa-ngiamwibool, P.; Namwat, W.; Chatrchaiwiwatana, S.; Ungareewittaya, P. Association between infection with Campylobacter species, poor oral health and environmental risk factors on esophageal cancer: A hospital-based case–control study in Thailand. Eur. J. Med. Res. 2021, 26, 82. [Google Scholar] [CrossRef]
- Su, S.-C.; Chang, L.-C.; Huang, H.-D.; Peng, C.-Y.; Chuang, C.-Y.; Chen, Y.-T.; Lu, M.Y.; Chiu, Y.W.; Chen, P.Y.; Yang, S.F. Oral microbial dysbiosis and its performance in predicting oral cancer. Carcinogenesis 2021, 42, 127–135. [Google Scholar] [CrossRef]
- Amer, A.; Galvin, S.; Healy, C.M.; Moran, G.P. The Microbiome of Potentially Malignant Oral Leukoplakia Exhibits Enrichment for Fusobacterium, Leptotrichia, Campylobacter, and Rothia Species. Front. Microbiol. 2017, 8, 2391. [Google Scholar] [CrossRef]
- He, Z.; Gharaibeh, R.Z.; Newsome, R.C.; Pope, J.L.; Dougherty, M.W.; Tomkovich, S.; Pons, B.; Mirey, G.; Vignard, J.; Hendrixson, D.R.; et al. Campylobacter jejuni promotes colorectal tumorigenesis through the action of cytolethal distending toxin. Gut 2019, 68, 289–300. [Google Scholar] [CrossRef]
- Bik, E.M.; Long, C.D.; Armitage, G.C.; Loomer, P.; Emerson, J.; Mongodin, E.F.; Nelson, K.E.; Gill, S.R.; Fraser-Liggett, C.M.; Relman, D.A. Bacterial diversity in the oral cavity of 10 healthy individuals. ISME J. 2010, 4, 962–974. [Google Scholar] [CrossRef]
- Li, J.; Quinque, D.; Horz, H.-P.; Li, M.; Rzhetskaya, M.; Raff, J.A.; Hayes, M.G.; Stoneking, M. Comparative analysis of the human saliva microbiome from different climate zones: Alaska, Germany, and Africa. BMC Microbiol. 2014, 14, 316. [Google Scholar] [CrossRef]
- Gopinath, D.; Kunnath Menon, R.; Chun Wie, C.; Banerjee, M.; Panda, S.; Mandal, D.; Behera, P.K.; Roychoudhury, S.; Kheur, S.; George Botelho, M.; et al. Salivary bacterial shifts in oral leukoplakia resemble the dysbiotic oral cancer bacteriome. J. Oral Microbiol. 2020, 13, 1857998. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Shen, W.; Wang, J.; Xu, Y.; Zhai, R.; Zhang, J.; Wang, M.; Wang, M.; Liu, L. Capnocytophaga gingivalis is a potential tumor promotor in oral cancer. Oral Dis. 2022, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; He, P.; Zhou, M.; Li, S.; Zhang, J.; Tao, X.; Wang, A.; Wu, X. Variations in oral microbiome and its predictive functions between tumorous and healthy individuals. J. Med. Microbiol. 2022, 71, 001568. [Google Scholar] [CrossRef] [PubMed]
- Saxena, R.; PK, V.P.; Gupta, S.V.; Gupta, S.; Waiker, P.; Samaiya, A.; Sharma, A.K. Assessing the Effect of Smokeless Tobacco Consumption on Oral Microbiome in Healthy and Oral Cancer Patients. Front. Cell. Infect. Microbiol. 2022, 12, 841465. [Google Scholar] [CrossRef]
- Kim, Y.K.; Kwon, E.J.; Yu, Y.; Kim, J.; Woo, S.-Y.; Choi, H.-S.; Kwon, M.; Jung, K.; Kim, H.S.; Park, H.R.; et al. Microbial and molecular differences according to the location of head and neck cancers. Cancer Cell Int. 2022, 22, 135. [Google Scholar] [CrossRef]
- Siqueira, J.F.J.; Rôças, I.N. Catonella morbi and Granulicatella adiacens: New species in endodontic infections. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2006, 102, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, Y.; Zheng, H.J.; Zhang, C.P. The Oral Microbiota May Have Influence on Oral Cancer. Front. Cell. Infect. Microbiol. 2019, 9, 476. [Google Scholar] [CrossRef] [PubMed]
- Moritani, K.; Takeshita, T.; Shibata, Y.; Ninomiya, T.; Kiyohara, Y.; Yamashita, Y. Acetaldehyde production by major oral microbes. Oral Dis. 2015, 21, 748–754. [Google Scholar] [CrossRef]
- Pushalkar, S.; Ji, X.; Li, Y.; Estilo, C.; Yegnanarayana, R.; Singh, B.; Li, X.; Saxena, D. Comparison of oral microbiota in tumor and non-tumor tissues of patients with oral squamous cell carcinoma. BMC Microbiol. 2012, 12, 144. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, S.; Takeshita, T.; Takeuchi, K.; Asakawa, M.; Matsumi, R.; Furuta, M.; Shibata, Y.; Nagai, K.; Ikebe, M.; Morita, M.; et al. Characteristics of the Salivary Microbiota in Patients With Various Digestive Tract Cancers. Front. Microbiol. 2019, 10, 1780. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.; Zhang, H.; Zheng, X.; Wang, J.; Jia, X.; Peng, X.; Xie, Q.; Zou, J.; Zheng, L.; et al. Probiotic Streptococcus salivarius K12 Alleviates Radiation-Induced Oral Mucositis in Mice. Front. Immunol. 2021, 12, 684824. [Google Scholar] [CrossRef]
- Yang, C.-Y.; Yeh, Y.-M.; Yu, H.-Y.; Chin, C.-Y.; Hsu, C.-W.; Liu, H.; Huang, P.J.; Hu, S.N.; Liao, C.T.; Chang, K.P.; et al. Oral Microbiota Community Dynamics Associated With Oral Squamous Cell Carcinoma Staging. Front. Microbiol. 2018, 9, 862. [Google Scholar] [CrossRef]
- Perera, M.; Al-Hebshi, N.N.; Perera, I.; Ipe, D.; Ulett, G.C.; Speicher, D.J.; Chen, T.; Johnson, N.W. Inflammatory Bacteriome and Oral Squamous Cell Carcinoma. J. Dent. Res. 2018, 97, 725–732. [Google Scholar] [CrossRef]
- Coker, O.O.; Dai, Z.; Nie, Y.; Zhao, G.; Cao, L.; Nakatsu, G.; Wu, W.K.; Wong, S.H.; Chen, Z.; Sung, J.J.Y.; et al. Mucosal microbiome dysbiosis in gastric carcinogenesis. Gut 2018, 67, 1024–1032. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Z.; Qi, Y.; Wen, X.; Zhang, L. Metagenomic Analysis Reveals a Changing Microbiome Associated With the Depth of Invasion of Oral Squamous Cell Carcinoma. Front. Microbiol. 2022, 13, 795777. [Google Scholar] [CrossRef]
- Mager, D.L.; Haffajee, A.D.; Devlin, P.M.; Norris, C.M.; Posner, M.R.; Goodson, J.M. The salivary microbiota as a diagnostic indicator of oral cancer: A descriptive, non-randomized study of cancer-free and oral squamous cell carcinoma subjects. J. Transl. Med. 2005, 3, 27. [Google Scholar] [CrossRef] [PubMed]
- Fredricks, D.N.; Schubert, M.M.; Myerson, D. Molecular identification of an invasive gingival bacterial community. Clin. Infect. Dis. 2005, 41, e1–e4. [Google Scholar] [CrossRef] [PubMed]
- Funada, H.; Machi, T.; Yoneyama, H.; Matsuda, T.; Miura, H.; Ezaki, T.; Yokota, Y. Capnocytophaga sputigena bacteremia associated with acute leukemia. Kansenshogaku zasshi. J. Jpn. Assoc. Infect. Dis. 1993, 67, 622–628. [Google Scholar]
- Mizuki, H.; Abe, R.; Mikami, T. Ultrastructural Changes during the Life Cycle of Mycoplasma salivarium in Oral Biopsies from Patients with Oral Leukoplakia. Front. Cell. Infect. Microbiol. 2017, 7, 403. [Google Scholar] [CrossRef]
- Henrich, B.; Rumming, M.; Sczyrba, A.; Velleuer, E.; Dietrich, R.; Gerlach, W.; Gombert, M.; Rahn, S.; Stoye, J.; Borkhardt, A.; et al. Mycoplasma salivarium as a dominant coloniser of Fanconi anaemia associated oral carcinoma. PLoS ONE 2014, 9, e92297. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.K.; Panda, M.; Das, A.K.; Rahman, T.; Das, R.; Das, K.; Sarma, A.; Kataki, A.C.; Chattopadhyay, I. Dysbiosis of salivary microbiome and cytokines influence oral squamous cell carcinoma through inflammation. Arch. Microbiol. 2021, 203, 137–152. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Genus | Control | HL | PVL | OSCC | PVL-OSCC | p-Value |
|---|---|---|---|---|---|---|---|
| Fusobacteriota | 24% | 25% | 19% | 34% | 30% | 0.021 * | |
| Fusobacterium | 14% | 19% | 14% | 25% | 26% | 0.006 * | |
| Leptotrichia | 10% | 5% | 5% | 9% | 4% | 0.056 | |
| Bacteroidota | 21% | 14% | 20% | 25% | 13% | 0.195 | |
| Porphyromonas | 9% | 3% | 6% | 3% | 3% | 0.433 | |
| Prevotella | 3% | 8% | 9% | 5% | 4% | 0.689 | |
| Tannerella | 3% | 0.002 * | |||||
| Capnocytophaga | 2% | 1% | 13% | 1% | 0.012 * | ||
| Alloprevotella | 1% | 1% | 2% | 2% | 0.422 | ||
| Bergeyella | 2% | 1% | 0.034 * | ||||
| Proteobacteria | 20% | 25% | 11% | 19% | 11% | 0.708 | |
| Haemophilus | 15% | 19% | 7% | 7% | 4% | 0.459 | |
| Neisseria | 2% | 5% | 1% | 3% | 0.518 | ||
| Aggregatibacter | 1% | 1% | 1% | 0.671 | |||
| Pseudomonas | 2% | 0.297 | |||||
| Firmicutes | 19% | 26% | 28% | 14% | 21% | 0.011 * | |
| Streptococcus | 10% | 19% | 13% | 3% | 3% | 0.002 * | |
| Gemella | 1% | 1% | 2% | 0.103 | |||
| Dialister | 1% | 0.398 | |||||
| Selenomonas | 1% | 3% | 0.236 | ||||
| Veillonella | 2% | 2% | 2% | 0.287 | |||
| Granulicatella | 1% | 1% | 0.182 | ||||
| Fenollaria | 1% | 0.546 | |||||
| Catonella | 1% | 4% | 0.001 * | ||||
| Mycoplasma | 3% | 2% | 0.229 | ||||
| Spirochaetota | 5% | 5% | 1% | 10% | 0.020 * | ||
| Treponema | 5% | 5% | 1% | 10% | 0.022 * | ||
| Campylobacterota | 2% | 3% | 11% | 4% | 14% | 0.002 * | |
| Campylobacter | 2% | 3% | 11% | 4% | 14% | 0.003 * | |
| Synergistota | 1% | 0.029 * | |||||
| Fretibacterium | 1% | 0.020 * | |||||
| Actinobacteriota | 1% | 2% | 2% | 0.059 | |||
| Rothia | 1% | 1% | 0.018 * | ||||
| Others | 7% | 5% | 4% | 3% | -- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herreros-Pomares, A.; Hervás, D.; Bagan-Debón, L.; Jantus-Lewintre, E.; Gimeno-Cardona, C.; Bagan, J. On the Oral Microbiome of Oral Potentially Malignant and Malignant Disorders: Dysbiosis, Loss of Diversity, and Pathogens Enrichment. Int. J. Mol. Sci. 2023, 24, 3466. https://doi.org/10.3390/ijms24043466
Herreros-Pomares A, Hervás D, Bagan-Debón L, Jantus-Lewintre E, Gimeno-Cardona C, Bagan J. On the Oral Microbiome of Oral Potentially Malignant and Malignant Disorders: Dysbiosis, Loss of Diversity, and Pathogens Enrichment. International Journal of Molecular Sciences. 2023; 24(4):3466. https://doi.org/10.3390/ijms24043466
Chicago/Turabian StyleHerreros-Pomares, Alejandro, David Hervás, Leticia Bagan-Debón, Eloísa Jantus-Lewintre, Concepción Gimeno-Cardona, and José Bagan. 2023. "On the Oral Microbiome of Oral Potentially Malignant and Malignant Disorders: Dysbiosis, Loss of Diversity, and Pathogens Enrichment" International Journal of Molecular Sciences 24, no. 4: 3466. https://doi.org/10.3390/ijms24043466
APA StyleHerreros-Pomares, A., Hervás, D., Bagan-Debón, L., Jantus-Lewintre, E., Gimeno-Cardona, C., & Bagan, J. (2023). On the Oral Microbiome of Oral Potentially Malignant and Malignant Disorders: Dysbiosis, Loss of Diversity, and Pathogens Enrichment. International Journal of Molecular Sciences, 24(4), 3466. https://doi.org/10.3390/ijms24043466