
Short Telomere Lesions with Dysplastic Metaplasia Histology May Represent Precancerous Lesions of Helicobacter pylori-Positive Gastric Mucosa


 ,
,
Abstract
1. Introduction
2. Results
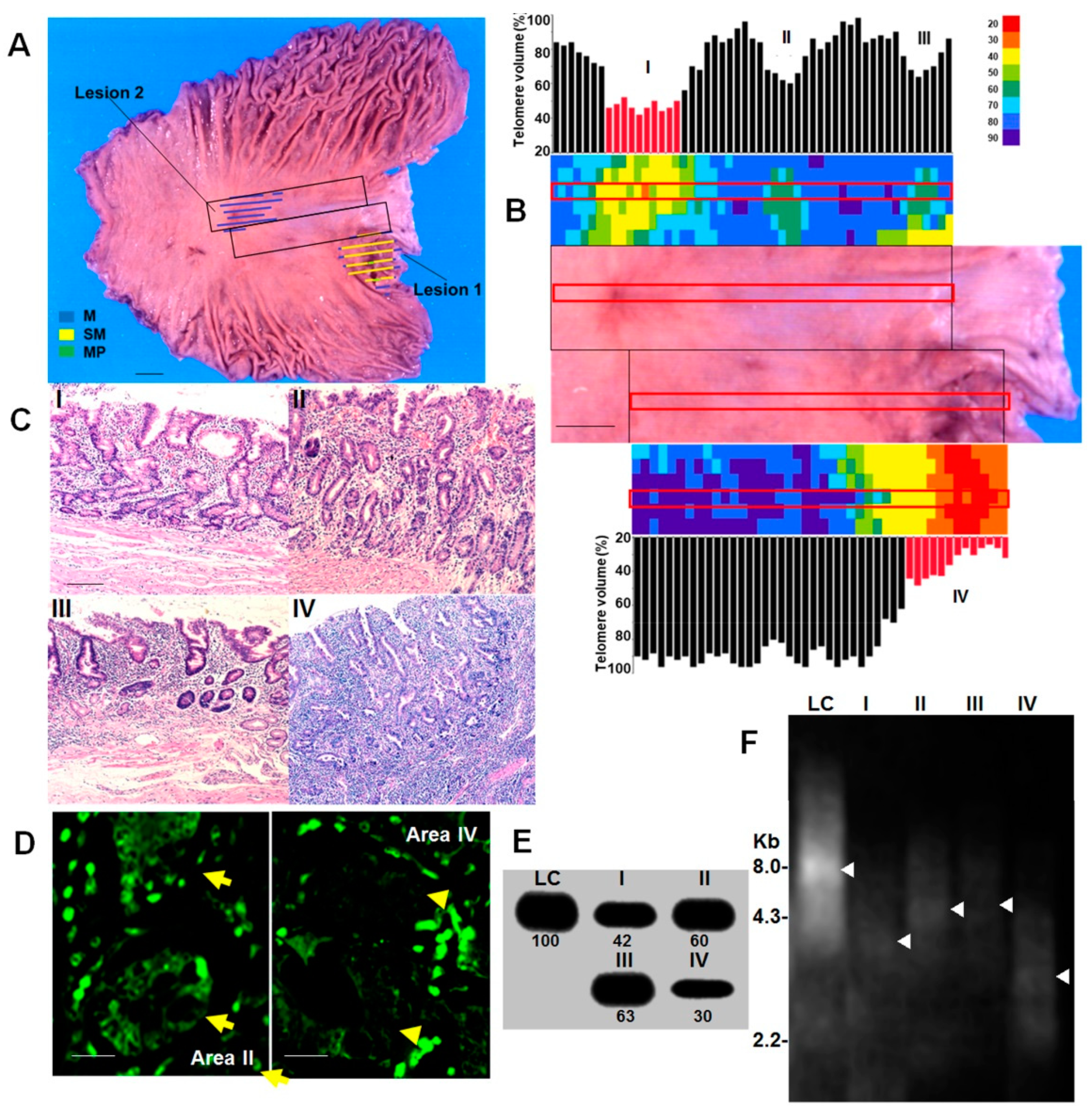
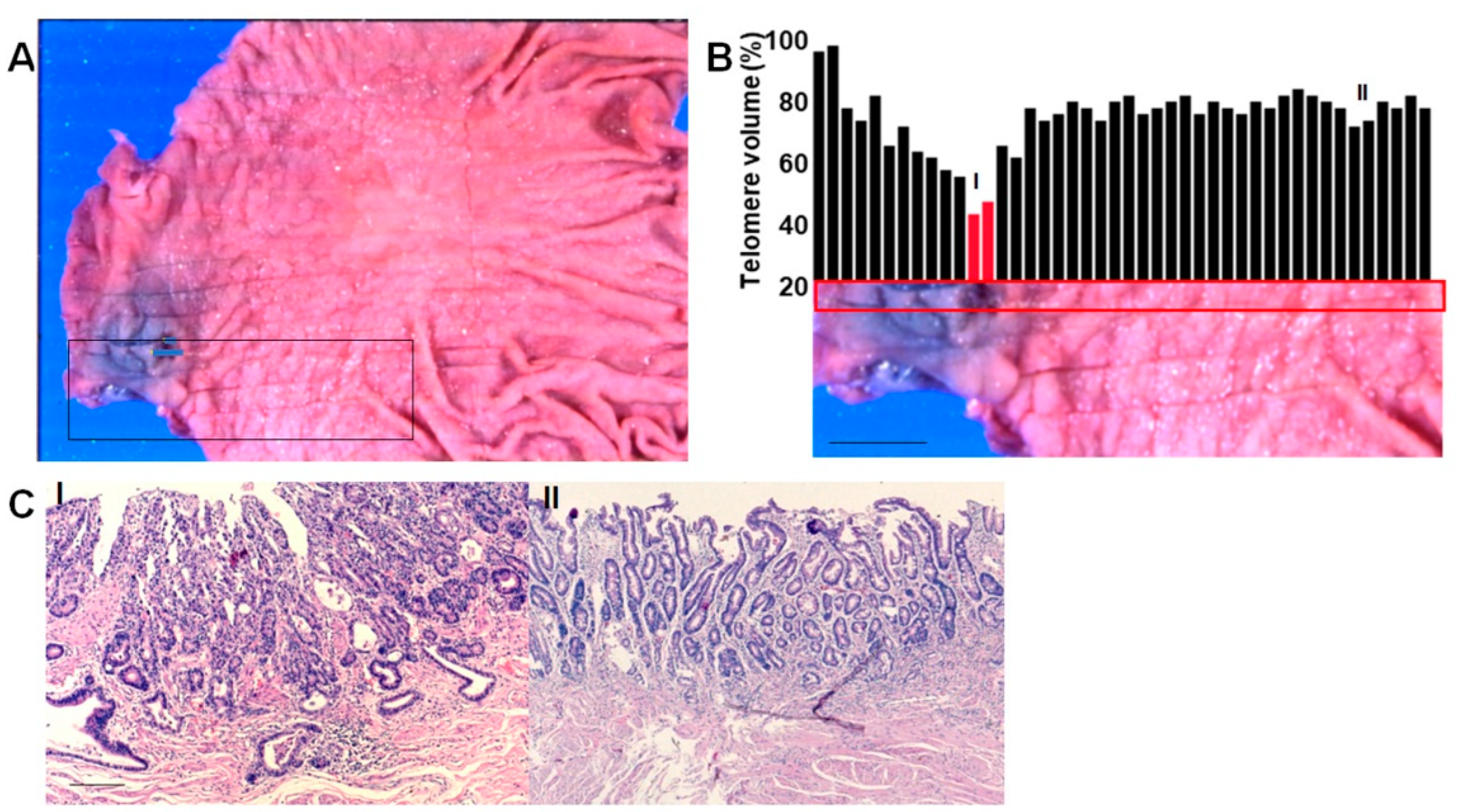
2.1. Short Telomere Lesions in Gastric Cancer Cases
2.2. Morphological Features of Short Telomere Lesions
2.3. Extraction of DM Cases
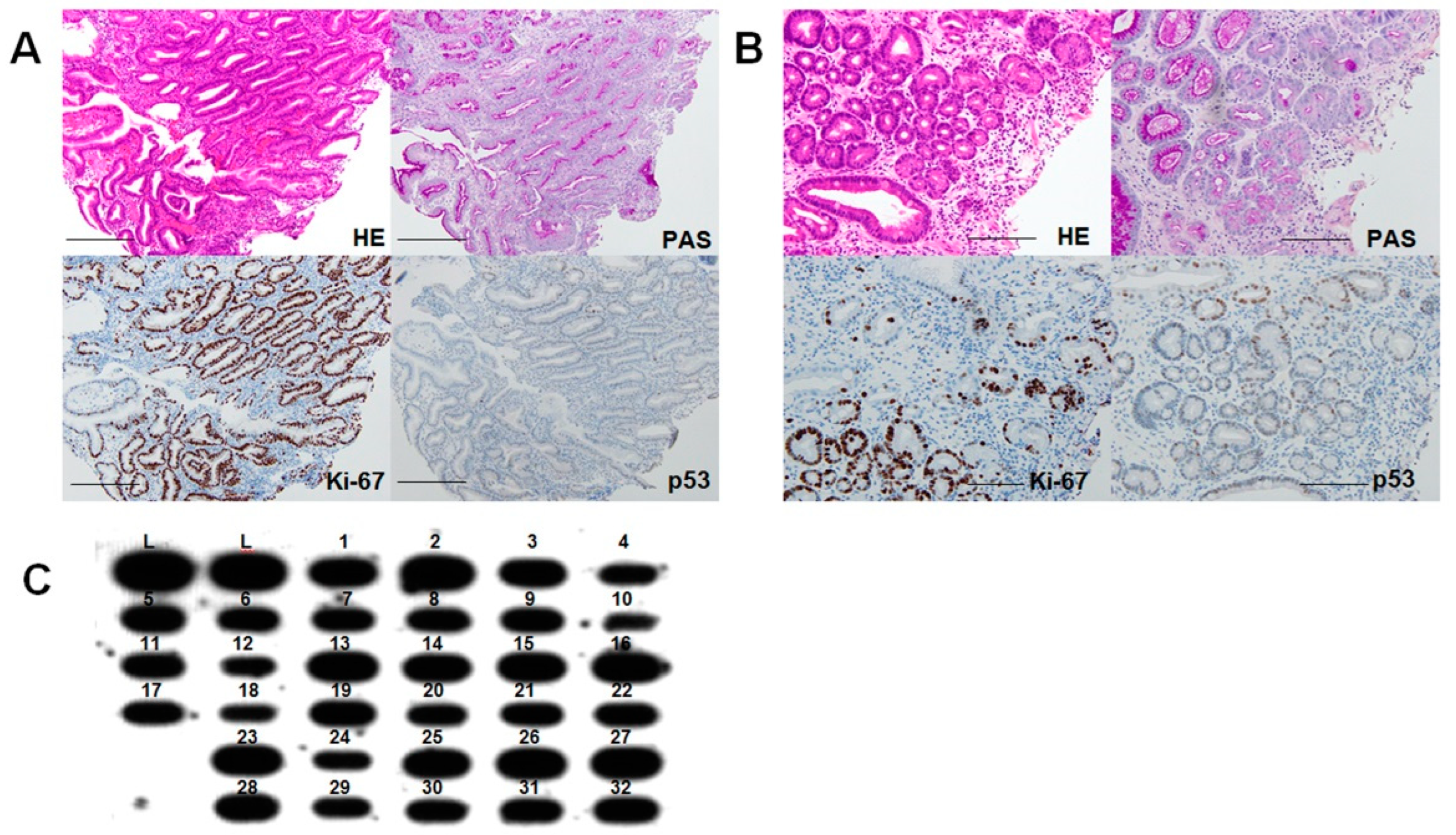
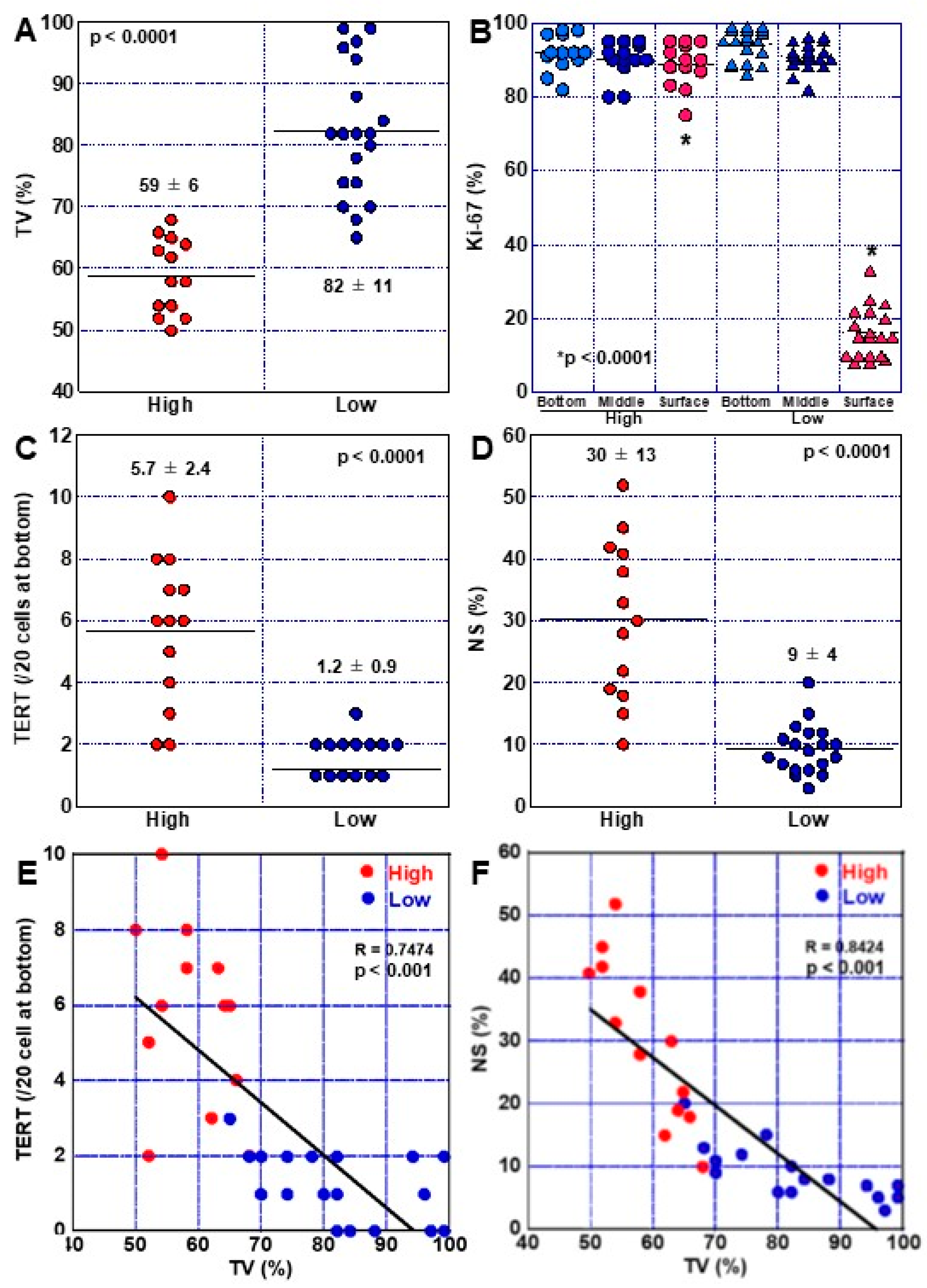
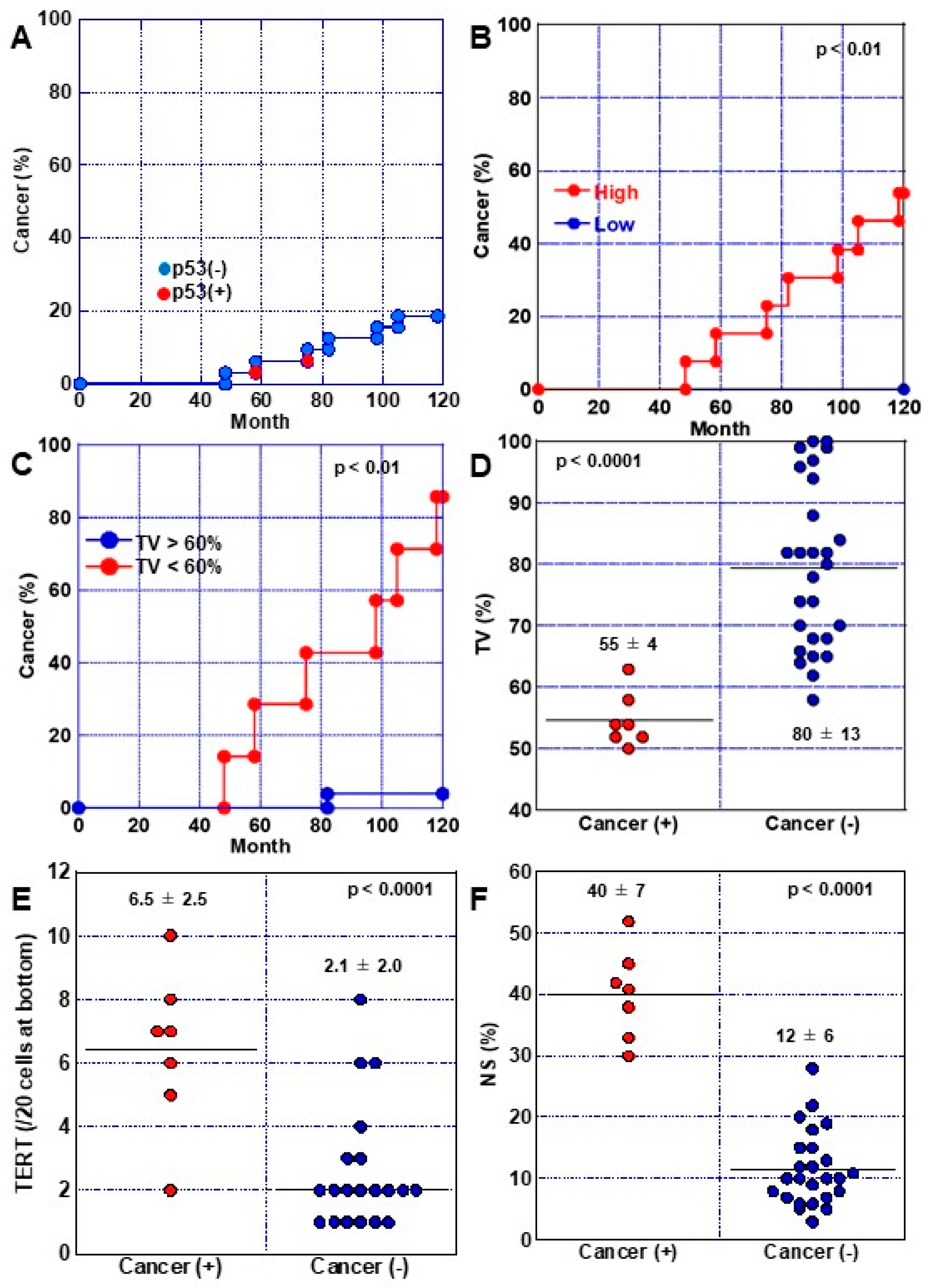
2.4. Phenotypes of High-Grade DM
2.5. Progression of DM to Cancer
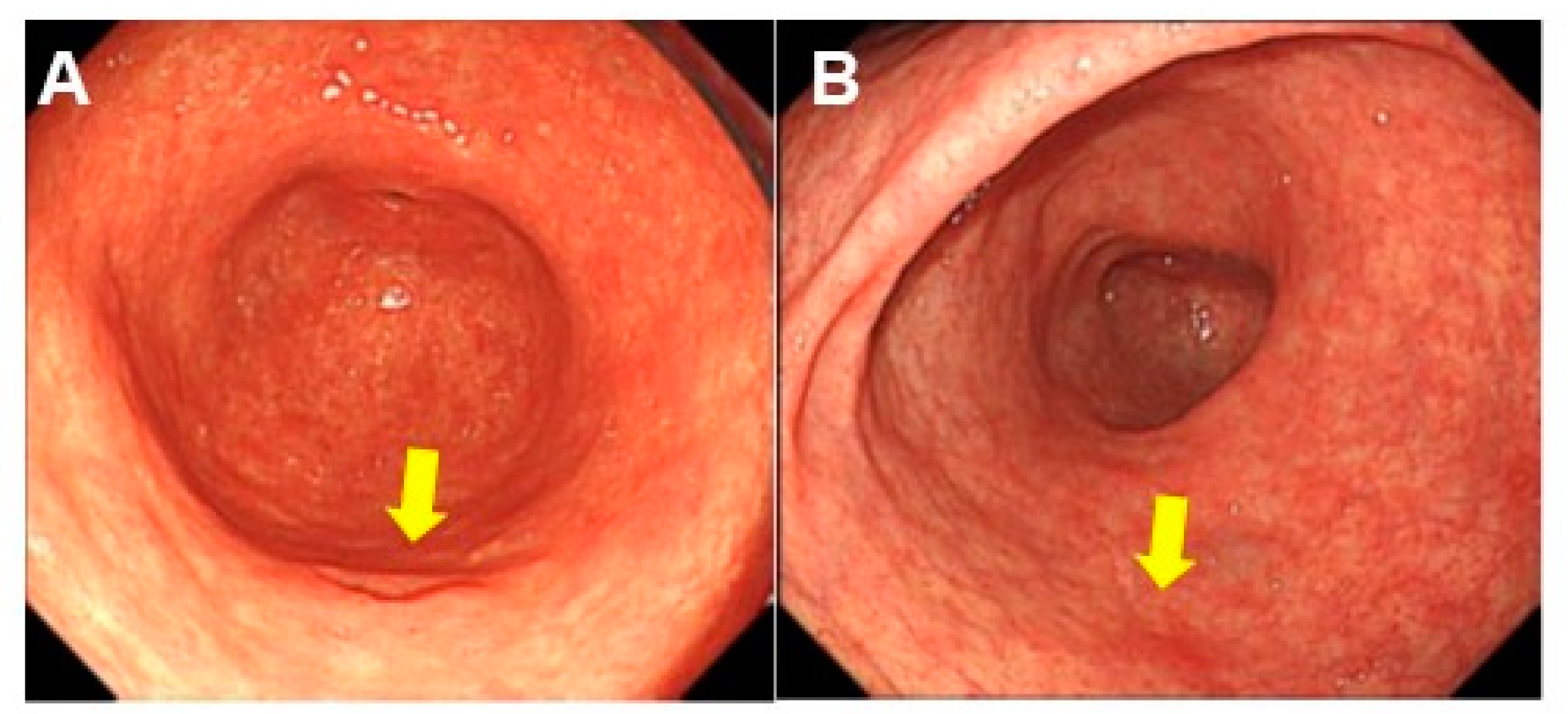
2.6. Endoscopic Characteristics Associated with DM
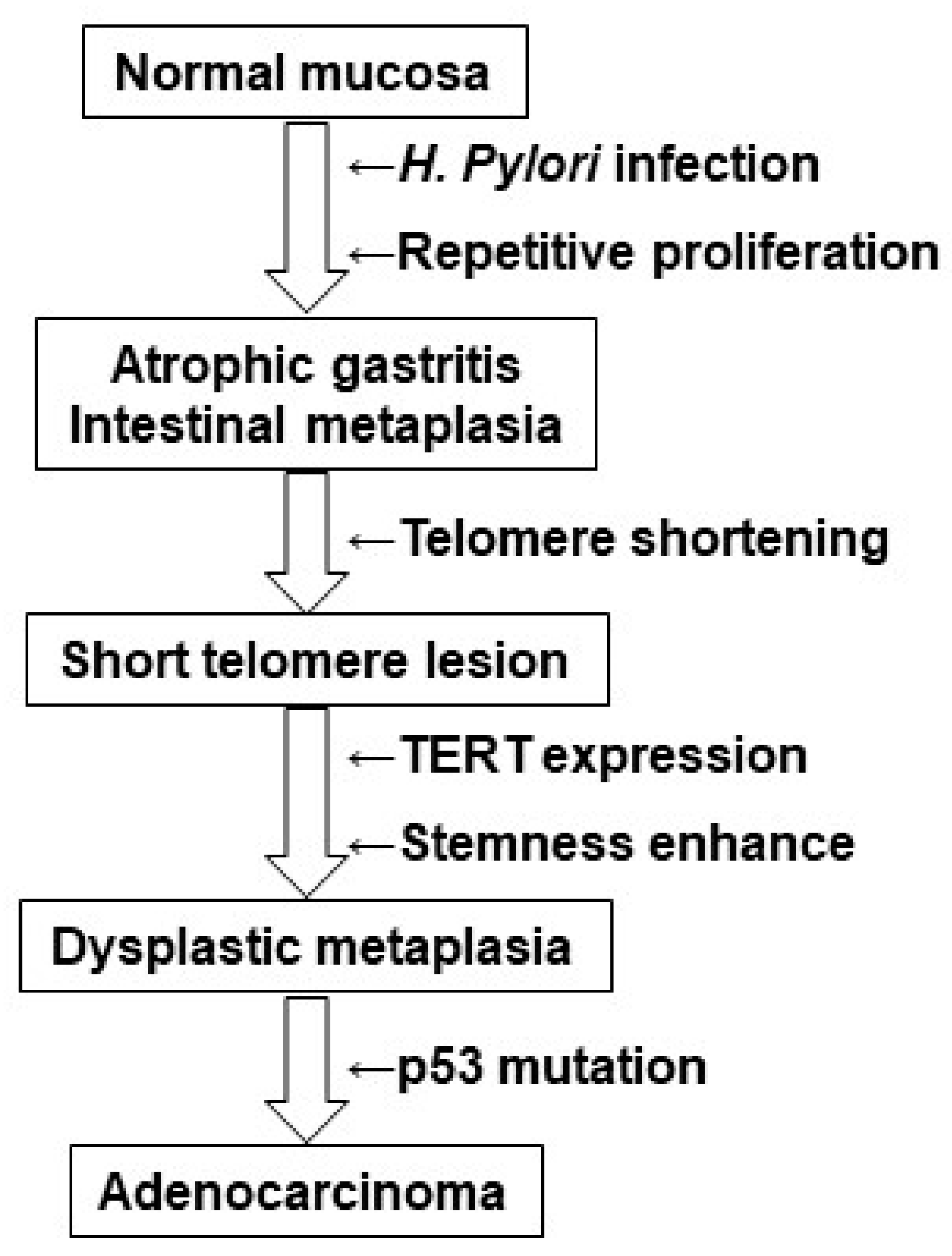
3. Discussion
4. Materials & Methods
4.1. Patients
4.2. Histological Analyses
4.3. Immunohistochemistry
4.4. Fluorescence In Situ Hybridization
4.5. Assessment of Telomere Volume
4.6. Laser Microdissection
4.7. Southern Blot Analysis
4.8. Slot Blot Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Chiang, T.H.; Chou, C.K.; Tu, Y.K.; Liao, W.C.; Wu, M.S.; Graham, D.Y. Association Between Helicobacter pylori Eradication and Gastric Cancer Incidence: A Systematic Review and Meta-analysis. Gastroenterology 2016, 150, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Kishikawa, H.; Ojiro, K.; Nakamura, K.; Katayama, T.; Arahata, K.; Takarabe, S.; Miura, S.; Kanai, T.; Nishida, J. Previous Helicobacter pylori infection-induced atrophic gastritis: A distinct disease entity in an understudied population without a history of eradication. Helicobacter 2020, 25, e12669. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, H.; Hayakawa, Y.; Koike, K. Metaplasia in the Stomach-Precursor of Gastric Cancer? Int. J. Mol. Sci. 2017, 18, 2063. [Google Scholar] [CrossRef]
- Battista, S.; Ambrosio, M.R.; Limarzi, F.; Gallo, G.; Saragoni, L. Molecular Alterations in Gastric Preneoplastic Lesions and Early Gastric Cancer. Int. J. Mol. Sci. 2021, 22, 6652. [Google Scholar] [CrossRef]
- Filipe, M.I.; Muñoz, N.; Matko, I.; Kato, I.; Pompe-Kirn, V.; Jutersek, A.; Teuchmann, S.; Benz, M.; Prijon, T. Intestinal metaplasia types and the risk of gastric cancer: A cohort study in Slovenia. Int. J. Cancer 1994, 57, 324–329. [Google Scholar] [CrossRef]
- Spence, A.D.; Cardwell, C.R.; McMenamin, Ú.C.; Hicks, B.M.; Johnston, B.T.; Murray, L.J.; Coleman, H.G. Adenocarcinoma risk in gastric atrophy and intestinal metaplasia: A systematic review. BMC Gastroenterol. 2017, 17, 157. [Google Scholar] [CrossRef]
- Laszkowska, M.; Truong, H.; Faye, A.S.; Kim, J.; Tan, S.X.; Lim, F.; Abrams, J.A.; Hur, C. Prevalence of Extensive and Limited Gastric Intestinal Metaplasia and Progression to Dysplasia and Gastric Cancer. Dig. Dis. Sci. 2022, 67, 3693–3701. [Google Scholar] [CrossRef]
- Kuniyasu, H.; Kitadai, Y.; Mieno, H.; Yasui, W. Helicobactor pylori infection is closely associated with telomere reduction in gastric mucosa. Oncology 2003, 65, 275–282. [Google Scholar] [CrossRef]
- Tahara, H.; Kuniyasu, H.; Yokozaki, H.; Yasui, W.; Shay, J.W.; Ide, T.; Tahara, E. Telomerase activity in preneoplastic and neoplastic gastric and colorectal lesions. Clin. Cancer Res. 1995, 1, 1245–1251. [Google Scholar]
- Kuniyasu, H.; Domen, T.; Hamamoto, T.; Yokozaki, H.; Yasui, W.; Tahara, H.; Tahara, E. Expression of human telomerase RNA is an early event of stomach carcinogenesis. Jpn J. Cancer Res. 1997, 88, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Harley, C.B. Telomerase and cancer therapeutics. Nat. Rev. Cancer 2008, 8, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Zvereva, M.I.; Shcherbakova, D.M.; Dontsova, O.A. Telomerase: Structure, functions, and activity regulation. Biochemistry 2010, 75, 1563–1583. [Google Scholar] [CrossRef]
- Yuan, X.; Larsson, C.; Xu, D. Mechanisms underlying the activation of TERT transcription and telomerase activity in human cancer: Old actors and new players. Oncogene 2019, 38, 6172–6183. [Google Scholar] [CrossRef]
- Tahara, E.; Kuniyasu, H.; Yasui, W.; Yokozaki, H. Gene alterations in intestinal metaplasia and gastric cancer. Eur. J. Gastroenterol. Hepatol. 1994, 6 (Suppl. 1), S97–S102. [Google Scholar] [PubMed]
- Kuniyasu, H.; Yasui, W.; Yokozaki, H.; Tahara, E. Helicobacter pylori infection and carcinogenesis of the stomach. Langenbecks Arch. Surg. 2000, 385, 69–74. [Google Scholar] [CrossRef]
- Li, K.; Wu, H.; Wang, A.; Charron, J.; Mishina, Y.; Habib, S.L.; Liu, H.; Li, B. mTOR signaling regulates gastric epithelial progenitor homeostasis and gastric tumorigenesis via MEK1-ERKs and BMP-Smad1 pathways. Cell Rep. 2021, 35, 109069. [Google Scholar] [CrossRef]
- Sasaki, T.; Kuniyasu, H.; Luo, Y.; Kitayoshi, M.; Tanabe, E.; Kato, D.; Shinya, S.; Fujii, K.; Ohmori, H.; Yamashita, Y. Increased phosphorylation of AKT in high-risk gastric mucosa. Anticancer Res. 2013, 33, 3295–3300. [Google Scholar] [PubMed]
- Sasaki, T.; Yamashita, Y.; Kuniyasu, H. AKT plays a crucial role in gastric cancer. Oncol. Lett. 2015, 10, 607–611. [Google Scholar] [CrossRef]
- Japanese Gastric Cancer Association. Japanese classification of gastric carcinoma: 3rd English edition. Gastric. Cancer 2011, 14, 101–112. [Google Scholar] [CrossRef]
- Kotelevets, S.M.; Chekh, S.A.; Chukov, S.Z. Updated Kimura-Takemoto classification of atrophic gastritis. World J. Clin. Cases 2021, 9, 3014–3023. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, X.; Ding, X.; Wang, F.; Geng, X. Telomere and its role in the aging pathways: Telomere shortening, cell senescence and mitochondria dysfunction. Biogerontology 2019, 20, 1–16. [Google Scholar] [CrossRef]
- Shay, J.W. Role of Telomeres and Telomerase in Aging and Cancer. Cancer Discov. 2016, 6, 584–593. [Google Scholar] [CrossRef]
- Thorsteinsdottir, S.; Gudjonsson, T.; Nielsen, O.H.; Vainer, B.; Seidelin, J.B. Pathogenesis and biomarkers of carcinogenesis in ulcerative colitis. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 395–404. [Google Scholar] [CrossRef]
- Drummond, M.W.; Balabanov, S.; Holyoake, T.L.; Brummendorf, T.H. Concise review: Telomere biology in normal and leukemic hematopoietic stem cells. Stem Cells 2007, 25, 1853–1861. [Google Scholar] [CrossRef]
- Londoño-Vallejo, J.A. Telomere instability and cancer. Biochimie 2008, 90, 73–82. [Google Scholar] [CrossRef]
- Armanios, M.; Greider, C.W. Telomerase and cancer stem cells. Cold Spring Harb. Symp. Quant. Biol. 2005, 70, 205–208. [Google Scholar] [CrossRef]
- Bell, R.J.; Rube, H.T.; Xavier-Magalhães, A.; Costa, B.M.; Mancini, A.; Song, J.S.; Costello, J.F. Understanding TERT Promoter Mutations: A Common Path to Immortality. Mol. Cancer Res. 2016, 14, 315–323. [Google Scholar] [CrossRef]
- Huang, D.S.; Wang, Z.; He, X.J.; Diplas, B.H.; Yang, R.; Killela, P.J.; Meng, Q.; Ye, Z.Y.; Wang, W.; Jiang, X.T.; et al. Recurrent TERT promoter mutations identified in a large-scale study of multiple tumour types are associated with increased TERT expression and telomerase activation. Eur. J. Cancer 2015, 51, 969–976. [Google Scholar] [CrossRef]
- Liu, T.; Liang, X.; Björkholm, M.; Jia, J.; Xu, D. The absence of TERT promoter mutations in primary gastric cancer. Gene 2014, 540, 266–267. [Google Scholar] [CrossRef]
- Hannen, R.; Bartsch, J.W. Essential roles of telomerase reverse transcriptase hTERT in cancer stemness and metastasis. FEBS Lett. 2018, 592, 2023–2031. [Google Scholar] [CrossRef]
- Kong, C.M.; Lee, X.W.; Wang, X. Telomere shortening in human diseases. FEBS J. 2013, 280, 3180–3193. [Google Scholar] [CrossRef]
- Sperka, T.; Song, Z.; Morita, Y.; Nalapareddy, K.; Guachalla, L.M.; Lechel, A.; Begus-Nahrmann, Y.; Burkhalter, M.D.; Mach, M.; Schlaudraff, F.; et al. Puma and p21 represent cooperating checkpoints limiting self-renewal and chromosomal instability of somatic stem cells in response to telomere dysfunction. Nat. Cell Biol. 2011, 14, 73–79. [Google Scholar] [CrossRef]
- Gunes, C.; Avila, A.I.; Rudolph, K.L. Telomeres in cancer. Differentiation 2018, 99, 41–50. [Google Scholar] [CrossRef]
- Correa, P. Helicobacter pylori and gastric carcinogenesis. Am. J. Surg. Pathol. 1995, 19 (Suppl. 1), S37–S43. [Google Scholar]
- Nagtegaal, I.; Odze, R.D.; Klismstra, D.; Paradis, V.; Rugge, M.; Schirmacher, P.; Washington, K.M.; Carnerio, F.; Cree, I.A. WHO Classification of Tumours of the Digestive System, 5th ed.; International Agency for Research on Cancer: Lyon, France, 2019. [Google Scholar]
- Estifan, E.; Cavanagh, Y.; Grossman, M.A. Hybrid Argon Plasma Coagulation for Treatment of Gastric Intestinal Metaplasia. Cureus 2020, 12, e7427. [Google Scholar] [CrossRef]
- Rugge, M.; Cassaro, M.; Di Mario, F.; Leo, G.; Leandro, G.; Russo, V.M.; Pennelli, G.; Farinati, F. The long term outcome of gastric non-invasive neoplasia. Gut 2003, 52, 1111–1116. [Google Scholar] [CrossRef]
- Sung, J.K. Diagnosis and management of gastric dysplasia. Korean J. Intern. Med. 2016, 31, 201–209. [Google Scholar] [CrossRef]
- Lauwers, G.Y.; Shimizu, M.; Correa, P.; Riddell, R.H.; Kato, Y.; Lewin, K.J.; Yamabe, H.; Sheahan, D.G.; Lewin, D.; Sipponen, P.; et al. Evaluation of gastric biopsies for neoplasia: Differences between Japanese and Western pathologists. Am. J. Surg. Pathol. 1999, 23, 511–518. [Google Scholar] [CrossRef]
- Li, Y.; Chang, X.; Zhou, W.; Xiao, Y.; Nakatsuka, L.N.; Chen, J.; Lauwers, G.Y. Gastric intestinal metaplasia with basal gland atypia: A morphological and biologic evaluation in a large Chinese cohort. Hum. Pathol. 2013, 44, 578–590. [Google Scholar] [CrossRef]
- Shichijo, S.; Hirata, Y. Characteristics and predictors of gastric cancer after Helicobacter pylori eradication. World J. Gastroenterol. 2018, 24, 2163–2172. [Google Scholar] [CrossRef]
- Pan, H.; Fang, C.; He, L.; Li, H.; Liu, L.; Wang, C.; Chen, J. Risk of multiple early gastric cancers in a patient with precursor lesions and endoscopic surveillance for 7 years: A case report. Medicine 2019, 98, e15686. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research#Network, Comprehensive molecular characterization of gastric adenocarcinoma. Nature 2014, 513, 202–209.
- Fujiwara-Tani, R.; Okamoto, A.; Katsuragawa, H.; Ohmori, H.; Fujii, K.; Mori, S.; Kishi, S.; Sasaki, T.; Nakashima, C.; Kawahara, I.; et al. BRAF Mutation Is Associated with Hyperplastic Polyp-Associated Gastric Cancer. Int. J. Mol. Sci. 2021, 22, 12724. [Google Scholar] [CrossRef]
- Hacia, J.G.; Novotny, E.A.; Mayer, R.A.; Woski, S.A.; Ashlock, M.A.; Collins, F.S. Design of modified oligodeoxyribonucleotide probes to detect telomere repeat sequences in FISH assays. Nucleic Acids Res. 1999, 27, 4034–4039. [Google Scholar] [CrossRef]
- Kuniyasu, H.; Luo, Y.; Fujii, K.; Sasahira, T.; Moriwaka, Y.; Tatsumoto, N.; Sasaki, T.; Yamashita, Y.; Ohmori, H. CD10 enhances metastasis of colorectal cancer by abrogating the anti-tumoural effect of methionine-enkephalin in the liver. Gut 2010, 59, 348–356. [Google Scholar] [CrossRef]
- Southern, E.M. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 1975, 98, 503–517. [Google Scholar] [CrossRef]
- Kuniyasu, H.; Yasui, W.; Kitadai, Y.; Yokozaki, H.; Ito, H.; Tahara, E. Frequent amplification of the c-met gene in scirrhous type stomach cancer. Biochem. Biophys. Res. Commun. 1992, 189, 227–232. [Google Scholar] [CrossRef]
- Kuniyasu, H.; Yasui, W.; Shimamoto, F.; Fujii, K.; Nakahara, M.; Asahara, T.; Dohi, K.; Tahara, E. Hepatoblastoma in an adult associated with c-met proto-oncogene imbalance. Pathol. Int. 1996, 46, 1005–1010. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Cancer | Short Telomere Lesion | ||||
|---|---|---|---|---|---|---|
| Histology (1) | TV (%) | Histology | Grade | TV (%) (2) | TERT | |
| 1 | tub2, pT1 (lesion 1, area I) | 43 | IM (around cancer) | Low | 65 | 5 |
| tub2 + por2, pT3 (lesion 2, area IV) | 32 | IM (around cancer) | High | 55 | 10 | |
| IM (area II) | High | 58 | 8 | |||
| IM (area III) | Low | 66 | 3 | |||
| 2 | tub1, pT1 | 45 | IM (around cancer) | High | 58 | 8 |
| IM | Low | 75 | 2 | |||
| 3 | tub1, pT1 (area I) | 47 | IM (area II) | Low | 72 | 1 |
| 4 | tub1, pT1 | 51 | IM (around cancer) | Low | 75 | 3 |
| IM | Low | 72 | 2 | |||
| 5 | tub2, pT1 | 42 | IM (around cancer) | Low | 68 | 2 |
| IM | High | 61 | 6 |
| Histological Feature | Low-Grade DM | High-Grade DM |
|---|---|---|
| Nuclear enlargement | Less than twice the size of lymphocyte nucleus | More than twice the size of lymphocyte nucleus |
| Area of nuclear enlargement | Less than 1/3 of gland base | More than 1/3 of gland base |
| Structural atypia | None | None |
| Mucus production | Reduced: mild to moderate | Reduced: moderate to marked |
| Goblet cell | Reduced: mild to moderate | Reduced: moderate to marked |
| Case | Grade | TV (%) | Ki-67 | (%) * | TERT † | NS | p53 | Months to Cancer | |
|---|---|---|---|---|---|---|---|---|---|
| Lower | Middle | Upper | (%) | (%) | diagnosis | ||||
| 1 | Low | 88 | 89 | 90 | 24 | 0 | 8 | 0 | - |
| 2 | Low | 99 | 92 | 90 | 15 | 0 | 5 | 0 | - |
| 3 | Low | 82 | 95 | 96 | 20 | 1 | 6 | 0 | - |
| 4 | High | 62 | 91 | 92 | 88 | 3 | 15 | 0 | - |
| 5 | Low | 74 | 96 | 95 | 25 | 2 | 12 | 0 | - |
| 6 | Low | 68 | 88 | 90 | 33 | 2 | 13 | 0 | - |
| 7 | High | 66 | 97 | 95 | 82 | 4 | 18 | 0 | - |
| 8 | High | 68 | 82 | 80 | 83 | 2 | 10 | 0 | - |
| 9 | Low | 70 | 99 | 95 | 22 | 1 | 11 | 0 | - |
| 10 | High | 52 | 98 | 92 | 95 | 5 | 45 | 5 | 58 |
| 11 | High | 65 | 92 | 90 | 88 | 6 | 22 | 0 | - |
| 12 | High | 50 | 85 | 80 | 75 | 8 | 41 | 8 | 75 |
| 13 | Low | 96 | 86 | 88 | 10 | 1 | 5 | 0 | - |
| 14 | Low | 94 | 98 | 92 | 8 | 2 | 7 | 0 | - |
| 15 | Low | 97 | 95 | 96 | 15 | 0 | 3 | 0 | - |
| 16 | Low | 99 | 99 | 94 | 10 | 2 | 7 | 0 | - |
| 17 | High | 64 | 98 | 95 | 90 | 6 | 19 | 0 | - |
| 18 | High | 54 | 90 | 88 | 92 | 6 | 52 | 0 | 98 |
| 19 | Low | 82 | 95 | 92 | 16 | 0 | 10 | 0 | - |
| 20 | High | 58 | 89 | 90 | 87 | 7 | 38 | 0 | 105 |
| 21 | High | 58 | 97 | 94 | 90 | 8 | 28 | 0 | - |
| 22 | High | 63 | 92 | 95 | 94 | 7 | 30 | 0 | 82 |
| 23 | Low | 82 | 98 | 92 | 9 | 2 | 10 | 0 | - |
| 24 | High | 52 | 92 | 90 | 95 | 2 | 42 | 0 | 48 |
| 25 | Low | 78 | 88 | 82 | 18 | 2 | 15 | 0 | - |
| 26 | Low | 82 | 98 | 90 | 15 | 1 | 10 | 0 | - |
| 27 | Low | 84 | 99 | 91 | 10 | 0 | 8 | 0 | - |
| 28 | Low | 80 | 93 | 88 | 8 | 1 | 6 | 0 | - |
| 29 | High | 54 | 92 | 90 | 95 | 10 | 33 | 0 | 118 |
| 30 | Low | 65 | 95 | 91 | 22 | 3 | 20 | 0 | - |
| 31 | Low | 70 | 97 | 89 | 15 | 2 | 9 | 0 | - |
| 32 | Low | 74 | 89 | 85 | 10 | 1 | 12 | 0 | - |
| Endoscopical Finding | Incidence | (%) | p Value |
|---|---|---|---|
| Grade | High grade | Low grade | |
| Age (yo) | 76 ± 12 | 70 ± 13 | NS |
| Sex (male:female) | 9:4 | 12:7 | NS |
| Location | NS | ||
| Antrum | 7 | 9 | |
| Corpus | 6 | 10 | |
| Cardia | 0 | 0 | |
| Size | <0.0001 | ||
| <1cm | 1 | 15 | |
| >1cm | 12 | 4 | |
| Atrophy * | 0.0007 | ||
| C-3 or O-1 | 0 | 11 | |
| O-2 or O-3 | 13 | 8 | |
| Shape | NS | ||
| Depression | 10 | 14 | |
| Elevation | 3 | 3 | |
| Flat | 0 | 2 | |
| Polyp | 0 | 0 | |
| Erosion | 0 | 0 | |
| Ulcer | 0 | 0 | |
| Color | NS | ||
| Normal | 3 | 10 | |
| Redness | 10 | 9 |
| Target * | Manufacturer † | Clone | Code | Working Concentration (μg/mL) |
|---|---|---|---|---|
| Ki-67 | Dako | MIB-1 | M7240 | 0.5 |
| TERT | Santa-Cruz | A-6 | sc-393013 | 1 |
| CDX2 | Abcam | CDX2-88 | ab157524 | 0.2 |
| p53 | Abcam | PAb240 | ab26 | 1 |
| NS | Abcam | - | ab70346 | 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujiwara-Tani, R.; Takagi, T.; Mori, S.; Kishi, S.; Nishiguchi, Y.; Sasaki, T.; Ikeda, M.; Nagai, K.; Bhawal, U.K.; Ohmori, H.; et al. Short Telomere Lesions with Dysplastic Metaplasia Histology May Represent Precancerous Lesions of Helicobacter pylori-Positive Gastric Mucosa. Int. J. Mol. Sci. 2023, 24, 3182. https://doi.org/10.3390/ijms24043182
Fujiwara-Tani R, Takagi T, Mori S, Kishi S, Nishiguchi Y, Sasaki T, Ikeda M, Nagai K, Bhawal UK, Ohmori H, et al. Short Telomere Lesions with Dysplastic Metaplasia Histology May Represent Precancerous Lesions of Helicobacter pylori-Positive Gastric Mucosa. International Journal of Molecular Sciences. 2023; 24(4):3182. https://doi.org/10.3390/ijms24043182
Chicago/Turabian StyleFujiwara-Tani, Rina, Tadataka Takagi, Shiori Mori, Shingo Kishi, Yukiko Nishiguchi, Takamitsu Sasaki, Masayuki Ikeda, Kenta Nagai, Ujjal Kumar Bhawal, Hitoshi Ohmori, and et al. 2023. "Short Telomere Lesions with Dysplastic Metaplasia Histology May Represent Precancerous Lesions of Helicobacter pylori-Positive Gastric Mucosa" International Journal of Molecular Sciences 24, no. 4: 3182. https://doi.org/10.3390/ijms24043182
APA StyleFujiwara-Tani, R., Takagi, T., Mori, S., Kishi, S., Nishiguchi, Y., Sasaki, T., Ikeda, M., Nagai, K., Bhawal, U. K., Ohmori, H., Fujii, K., & Kuniyasu, H. (2023). Short Telomere Lesions with Dysplastic Metaplasia Histology May Represent Precancerous Lesions of Helicobacter pylori-Positive Gastric Mucosa. International Journal of Molecular Sciences, 24(4), 3182. https://doi.org/10.3390/ijms24043182