


A Picrocrocin-Enriched Fraction from a Saffron Extract Affects Lipid Homeostasis in HepG2 Cells through a Non-Statin-like Mode
 ,
,  , , ,
, , ,  ,
,  , , , , and
, , , , and
Abstract
1. Introduction
2. Results and Discussion
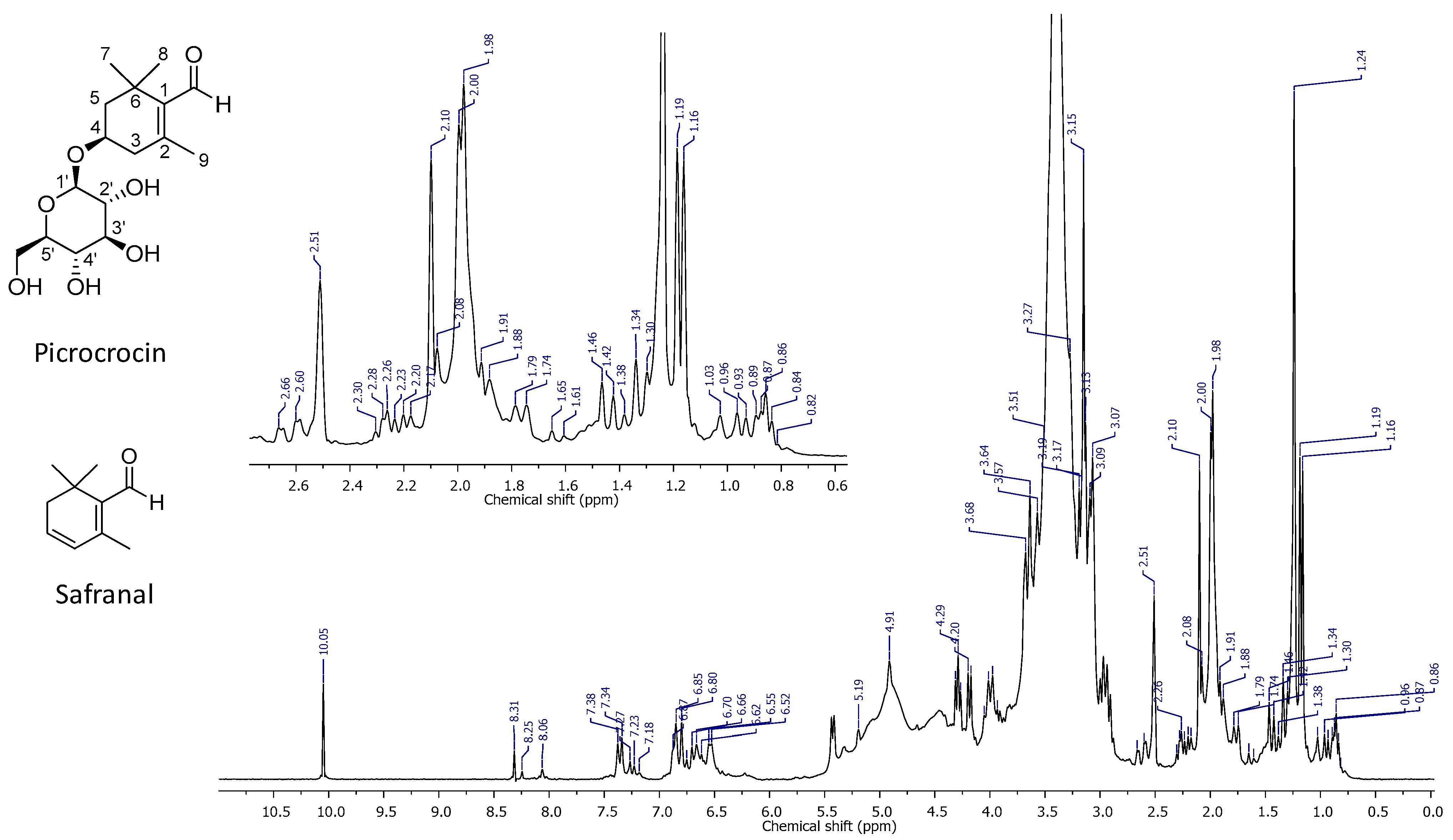
2.1. High-Resolution 1H NMR Spectrum of PEF
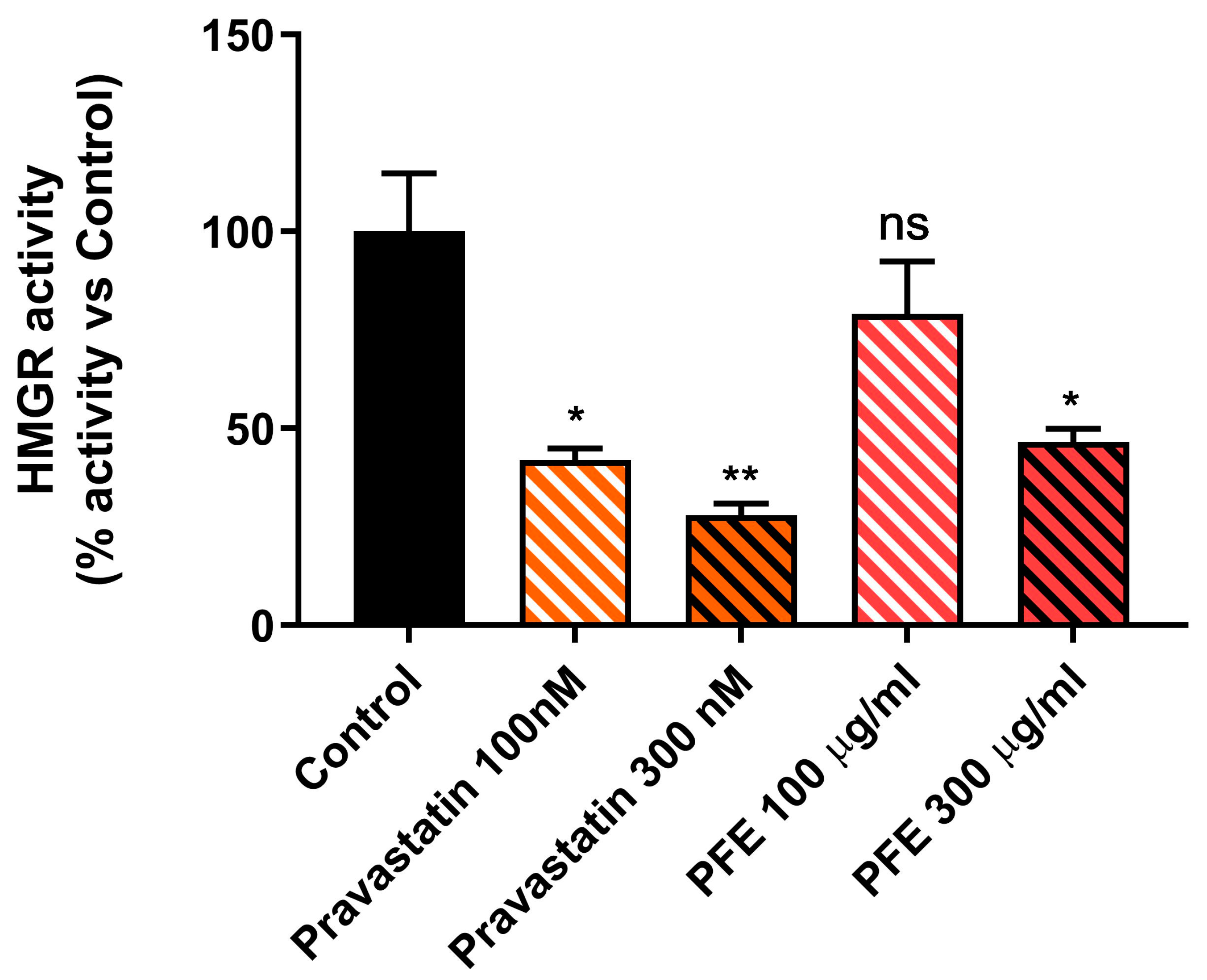
2.2. Picrocrocin Inhibits the Activity of the Purified Human Catalytic Fragment (cf-HMGR)
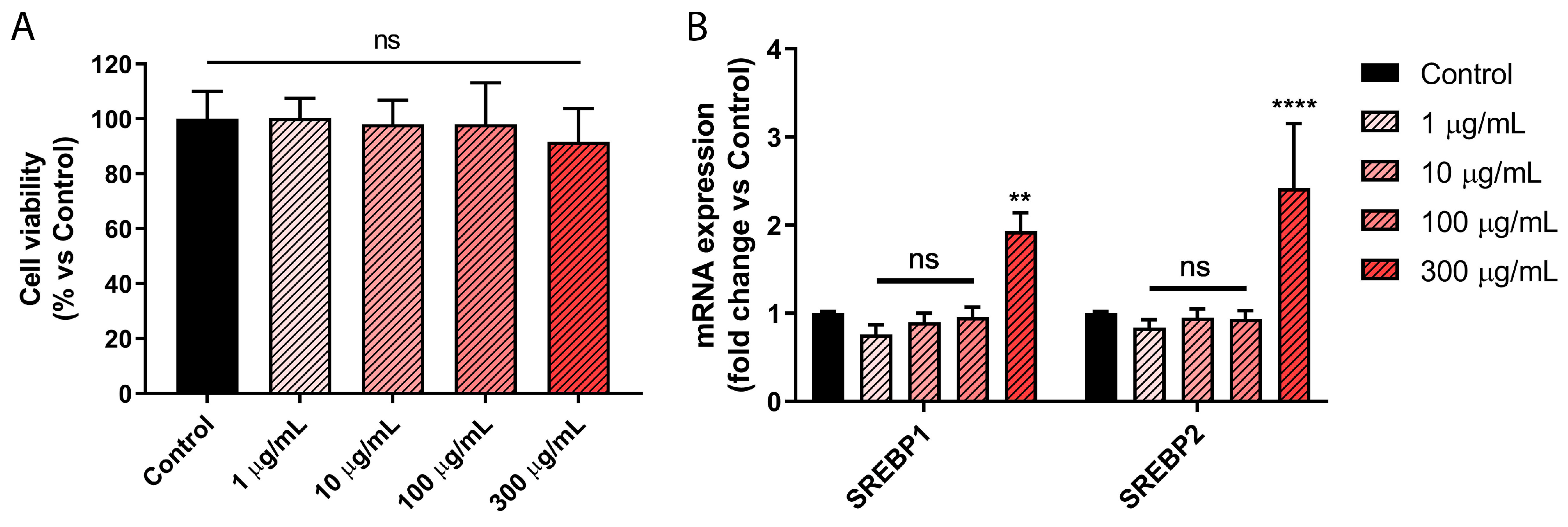
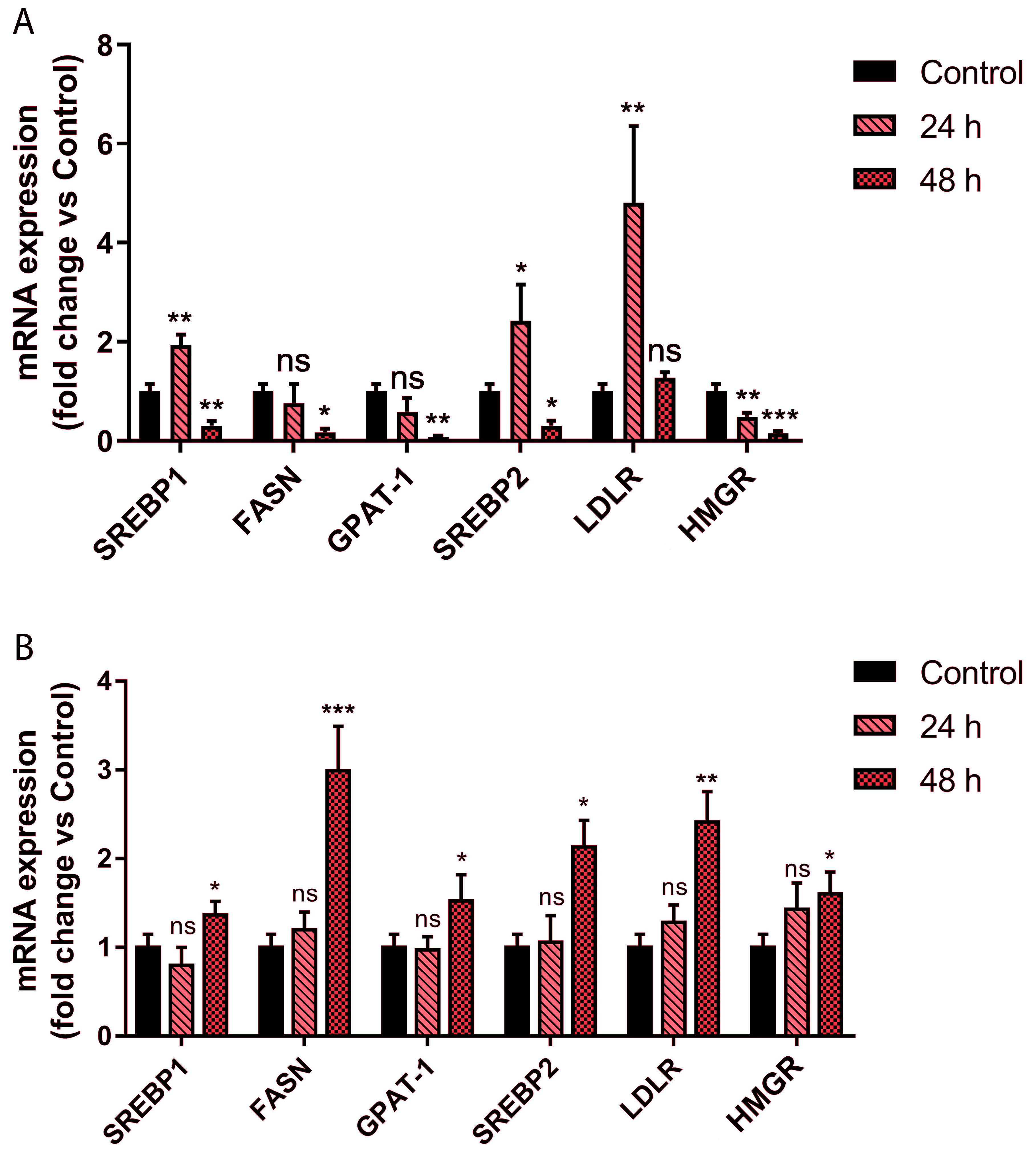
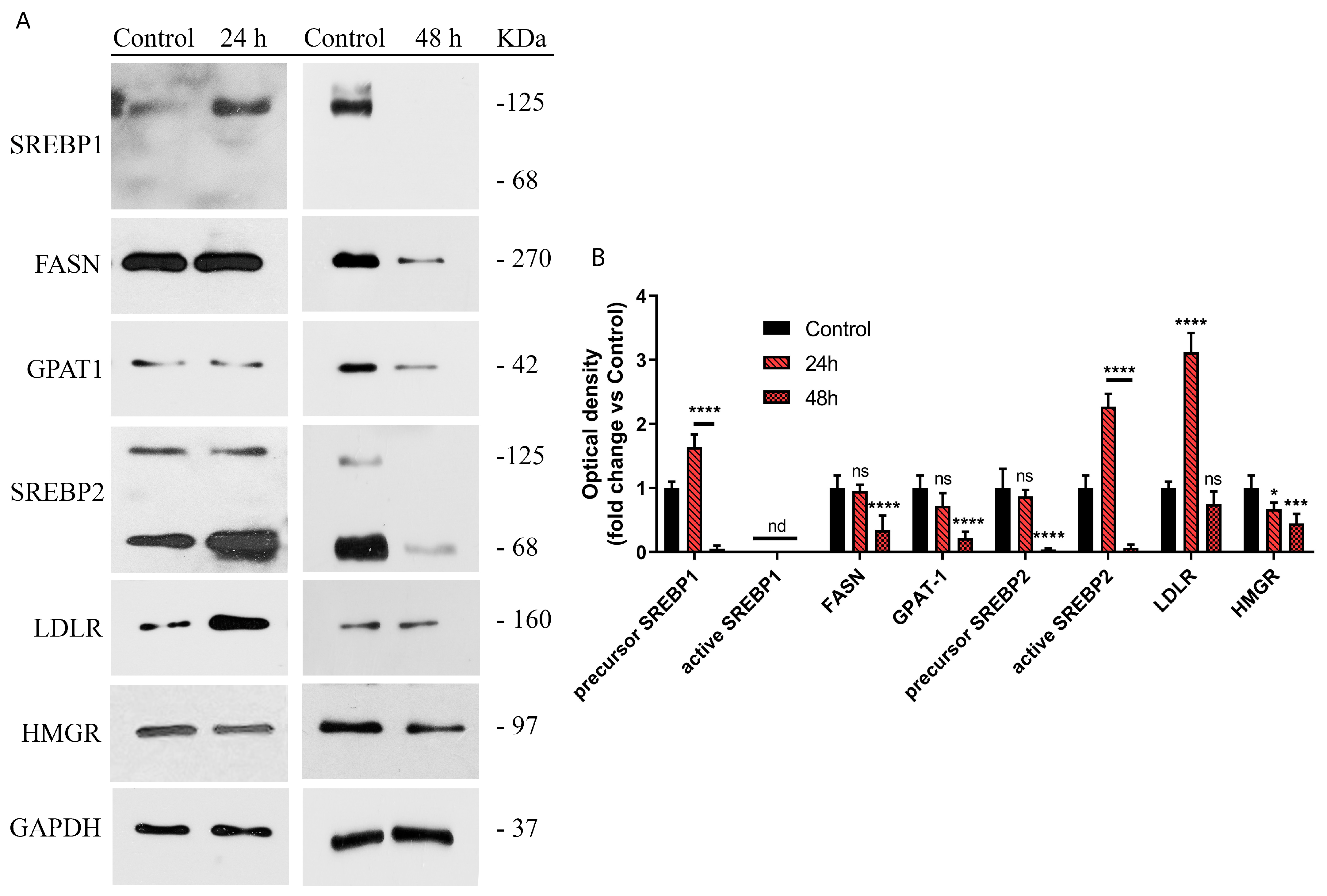
2.3. Picrocrocin Affects Lipid Homeostasis in HepG2 Cells with a Non-Statin-like Mode of Action
3. Materials and Methods
3.1. Chemicals
3.2. NMR Spectroscopy. Experimental Details
3.3. Sample Preparation
3.4. Enzymatic Activity of the Purified Human Catalytic Fragment (cf-HMGR)
3.5. Cell Cultures
3.6. Cell Viability Assay
3.7. Quantitative PCR with Reverse Transcription (qRT–PCR)
3.8. Immunoblotting Analysis
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Hannon, B.A.; Khan, N.A.; Teran-Garcia, M. Nutrigenetic Contributions to Dyslipidemia: A Focus on Physiologically Relevant Pathways of Lipid and Lipoprotein Metabolism. Nutrients 2018, 10, 1404. [Google Scholar] [CrossRef]
- Abdela, O.A.; Ayalew, M.B.; Yesuf, J.S.; Getnet, S.A.; Biyazin, A.A.; Lemma, M.Y.; Yimer, B.T. Ethiopian university students’ knowledge and perception towards cardiovascular disease risk factors: A cross sectional study. Am. J. Cardiovasc. Dis. 2019, 9, 1–7. [Google Scholar] [PubMed]
- Mayneris-Perxachs, J.; Puig, J.; Burcelin, R.; Dumas, M.-E.; Barton, R.H.; Hoyles, L.; Federici, M.; Fernández-Real, J.-M. The APOA1bp-SREBF-NOTCH axis is associated with reduced atherosclerosis risk in morbidly obese patients. Clin. Nutr. 2020, 39, 3408–3418. [Google Scholar] [CrossRef] [PubMed]
- Fiorillo, M.; Peiris-Pages, M.; Sanchez-Alvarez, R.; Bartella, L.; Di Donna, L.; Dolce, V.; Sindona, G.; Sotgia, F.; Cappello, A.R.; Lisanti, M.P. Bergamot natural products eradicate cancer stem cells (CSCs) by targeting mevalonate, Rho-GDI-signalling and mitochondrial metabolism. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 984–996. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.; Guido, C.; De Amicis, F.; Sisci, D.; Cione, E.; Vincenza, D.; Dona, A.; Panno, M.L.; Aquila, S. Bergapten induces metabolic reprogramming in breast cancer cells. Oncol. Rep. 2016, 35, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Frattaruolo, L.; Carullo, G.; Brindisi, M.; Mazzotta, S.; Bellissimo, L.; Rago, V.; Curcio, R.; Dolce, V.; Aiello, F.; Cappello, A.R. Antioxidant and Anti-Inflammatory Activities of Flavanones from Glycyrrhiza glabra L. (licorice) Leaf Phytocomplexes: Identification of Licoflavanone as a Modulator of NF-kB/MAPK Pathway. Antioxidants 2019, 8, 186. [Google Scholar] [CrossRef]
- Cappello, A.R.; Dolce, V.; Iacopetta, D.; Martello, M.; Fiorillo, M.; Curcio, R.; Muto, L.; Dhanyalayam, D. Bergamot (Citrus bergamia Risso) Flavonoids and Their Potential Benefits in Human Hyperlipidemia and Atherosclerosis: An Overview. Mini-Rev. Med. Chem. 2016, 16, 619–629. [Google Scholar] [CrossRef]
- Frattaruolo, L.; Brindisi, M.; Curcio, R.; Marra, F.; Dolce, V.; Cappello, A.R. Targeting the Mitochondrial Metabolic Network: A Promising Strategy in Cancer Treatment. Int. J. Mol. Sci. 2020, 21, 6014. [Google Scholar] [CrossRef]
- Brindisi, M.; Frattaruolo, L.; Fiorillo, M.; Dolce, V.; Sotgia, F.; Lisanti, M.P.; Cappello, A.R. New insights into cholesterol-mediated ERRalpha activation in breast cancer progression and pro-tumoral microenvironment orchestration. FEBS J. 2022. [Google Scholar] [CrossRef]
- Schumacher, M.M.; DeBose-Boyd, R.A. Posttranslational Regulation of HMG CoA Reductase, the Rate-Limiting Enzyme in Synthesis of Cholesterol. Annu. Rev. Biochem. 2021, 90, 659–679. [Google Scholar] [CrossRef]
- Lim, S.; Park, Y.M.; Sakuma, I.; Koh, K.K. How to control residual cardiovascular risk despite statin treatment: Focusing on HDL–cholesterol. Int. J. Cardiol. 2013, 166, 8–14. [Google Scholar] [CrossRef]
- Crismaru, I.; Pantea Stoian, A.; Bratu, O.G.; Gaman, M.A.; Stanescu, A.M.A.; Bacalbasa, N.; Diaconu, C.C. Low-density lipoprotein cholesterol lowering treatment: The current approach. Lipids Health Dis. 2020, 19, 85. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, C.; Alegria, E.; Bonadonna, R.C.; Catapano, A.L.; Cosentino, F.; Elisaf, M.; Farnier, M.; Ferrieres, J.; Filardi, P.P.; Hancu, N.; et al. A review of the evidence on reducing macrovascular risk in patients with atherogenic dyslipidaemia: A report from an expert consensus meeting on the role of fenofibrate-statin combination therapy. Atheroscler. Suppl. 2015, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Parisi, O.I.; Fiorillo, M.; Scrivano, L.; Sinicropi, M.S.; Dolce, V.; Iacopetta, D.; Puoci, F.; Cappello, A.R. Sericin/Poly(ethylcyanoacrylate) Nanospheres by Interfacial Polymerization for Enhanced Bioefficacy of Fenofibrate: In Vitro and In Vivo Studies. Biomacromolecules 2015, 16, 3126–3133. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.K.; Sehgal, V.S.; Kashfi, K. Molecular targets of statins and their potential side effects: Not all the glitter is gold. Eur. J. Pharm. 2022, 922, 174906. [Google Scholar] [CrossRef] [PubMed]
- Malachowski, S.J.; Quattlebaum, A.M.; Miladinovic, B. Adverse Effects of Statins. Jama 2017, 317, 1079–1080. [Google Scholar] [CrossRef] [PubMed]
- du Souich, P.; Roederer, G.; Dufour, R. Myotoxicity of statins: Mechanism of action. Pharmacol. Ther. 2017, 175, 1–16. [Google Scholar] [CrossRef]
- Mach, F.; Ray, K.K.; Wiklund, O.; Corsini, A.; Catapano, A.L.; Bruckert, E.; De Backer, G.; Hegele, R.A.; Hovingh, G.K.; Jacobson, T.A.; et al. Adverse effects of statin therapy: Perception vs. the evidence—Focus on glucose homeostasis, cognitive, renal and hepatic function, haemorrhagic stroke and cataract. Eur. Heart J. 2018, 39, 2526–2539. [Google Scholar] [CrossRef]
- Jones, P.H. Expert perspective: Reducing cardiovascular risk in metabolic syndrome and type 2 diabetes mellitus beyond low-density lipoprotein cholesterol lowering. Am. J. Cardiol. 2008, 102, 41L–47L. [Google Scholar] [CrossRef] [PubMed]
- Mollace, V.; Sacco, I.; Janda, E.; Malara, C.; Ventrice, D.; Colica, C.; Visalli, V.; Muscoli, S.; Ragusa, S.; Muscoli, C.; et al. Hypolipemic and hypoglycaemic activity of bergamot polyphenols: From animal models to human studies. Fitoterapia 2011, 82, 309–316. [Google Scholar] [CrossRef]
- Parizad, S.; Dizadji, A.; Habibi, M.K.; Winter, S.; Kalantari, S.; Movi, S.; Lorenzo Tendero, C.; Alonso, G.L.; Moratalla-Lopez, N. The effects of geographical origin and virus infection on the saffron (Crocus sativus L.) quality. Food Chem. 2019, 295, 387–394. [Google Scholar] [CrossRef] [PubMed]
- El Midaoui, A.; Ghzaiel, I.; Vervandier-Fasseur, D.; Ksila, M.; Zarrouk, A.; Nury, T.; Khallouki, F.; El Hessni, A.; Ibrahimi, S.O.; Latruffe, N.; et al. Saffron (Crocus sativus L.): A Source of Nutrients for Health and for the Treatment of Neuropsychiatric and Age-Related Diseases. Nutrients 2022, 14, 597. [Google Scholar] [CrossRef] [PubMed]
- Xing, B.; Li, S.; Yang, J.; Lin, D.; Feng, Y.; Lu, J.; Shao, Q. Phytochemistry, pharmacology, and potential clinical applications of saffron: A review. J. Ethnopharmacol. 2021, 281, 114555. [Google Scholar] [CrossRef] [PubMed]
- Korani, S.; Korani, M.; Sathyapalan, T.; Sahebkar, A. Therapeutic effects of Crocin in autoimmune diseases: A review. Biofactors 2019, 45, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Shao, Q.; Lu, Z.; Duan, C.; Yi, H.; Su, L. Rapid determination of crocins in saffron by near-infrared spectroscopy combined with chemometric techniques. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2018, 190, 283–289. [Google Scholar] [CrossRef]
- Nanda, S.; Madan, K. The role of Safranal and saffron stigma extracts in oxidative stress, diseases and photoaging: A systematic review. Heliyon 2021, 7, e06117. [Google Scholar] [CrossRef] [PubMed]
- Bukhari, S.I.; Manzoor, M.; Dhar, M.K. A comprehensive review of the pharmacological potential of Crocus sativus and its bioactive apocarotenoids. Biomed. Pharmacother. 2018, 98, 733–745. [Google Scholar] [CrossRef]
- Mir, M.A.; Ganai, S.A.; Mansoor, S.; Jan, S.; Mani, P.; Masoodi, K.Z.; Amin, H.; Rehman, M.U.; Ahmad, P. Isolation, purification and characterization of naturally derived Crocetin beta-d-glucosyl ester from Crocus sativus L. against breast cancer and its binding chemistry with ER-alpha/HDAC2. Saudi J. Biol. Sci. 2020, 27, 975–984. [Google Scholar] [CrossRef]
- Colapietro, A.; Mancini, A.; D’Alessandro, A.M.; Festuccia, C. Crocetin and Crocin from Saffron in Cancer Chemotherapy and Chemoprevention. Anticancer Agents Med. Chem. 2019, 19, 38–47. [Google Scholar] [CrossRef]
- Shakeri, M.; Hashemi Tayer, A.; Shakeri, H.; Sotoodeh Jahromi, A.; Moradzadeh, M.; Hojjat-Farsangi, M. Toxicity of Saffron Extracts on Cancer and Normal Cells: A Review Article. Asian Pac. J. Cancer Prev. 2020, 21, 1867–1875. [Google Scholar] [CrossRef]
- Erfanparast, A.; Tamaddonfard, E.; Henareh-Chareh, F. Central H(2) histaminergic and alpha-2 adrenergic receptors involvement in crocetin-induced antinociception in orofacial formalin pain in rats. Vet. Res. Forum 2020, 11, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Li Puma, S.; Landini, L.; Macedo, S.J., Jr.; Seravalli, V.; Marone, I.M.; Coppi, E.; Patacchini, R.; Geppetti, P.; Materazzi, S.; Nassini, R.; et al. TRPA1 mediates the antinociceptive properties of the constituent of Crocus sativus L., safranal. J. Cell Mol. Med. 2019, 23, 1976–1986. [Google Scholar] [CrossRef] [PubMed]
- Boskabady, M.-H.; Gholamnezhad, Z.; Khazdair, M.-R.; Tavakol-Afshari, J. Chapter 25—Antiinflammatory and immunomodulatory effects of saffron and its derivatives. In Saffron; Koocheki, A., Khajeh-Hosseini, M., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 405–421. [Google Scholar] [CrossRef]
- Asbaghi, O.; Sadeghian, M.; Sadeghi, O.; Rigi, S.; Tan, S.C.; Shokri, A.; Mousavi, S.M. Effects of saffron (Crocus sativus L.) supplementation on inflammatory biomarkers: A systematic review and meta-analysis. Phytother. Res. 2021, 35, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Arasteh, A.; Aliyev, A.; Khamnei, S.; Delazar, A.; Mesgari Abbasi, M.; Mehmannavaz, Y. Effects of hydromethanolic extract of saffron (Crocus sativus) on serum glucose, insulin and cholesterol levels in healthy male rats. J. Med. Plants 2010, 4, 397–402. [Google Scholar]
- Kianbakht, S.; Mozaffari, K. Effects of Saffron and its Active Constituents, Crocin and Safranal, on Prevention of Indomethacin Induced Gastric Ulcers in Diabetic and Nondiabetic Rats. J. Med. Plants 2009, 8, 30–38. [Google Scholar]
- Kianbakht, S.; Hajiaghaee, R. Anti-hyperglycemic Effects of Saffron and its Active Constituents, Crocin and Safranal, in Alloxan-Induced Diabetic Rats. J. Med. Plants 2011, 10, 82–89. [Google Scholar]
- Bian, Y.; Zhao, C.; Lee, S.M. Neuroprotective Potency of Saffron Against Neuropsychiatric Diseases, Neurodegenerative Diseases, and Other Brain Disorders: From Bench to Bedside. Front. Pharm. 2020, 11, 579052. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, H.; Karimi, G.; Niapoor, M. Antidepressant effects of Crocus sativus stigma extracts and its constituents, crocin and safranal, in mice. J. Med. Plants 2004, 3, 48–58. [Google Scholar]
- Sheng, L.; Qian, Z.; Zheng, S.; Xi, L. Mechanism of hypolipidemic effect of crocin in rats: Crocin inhibits pancreatic lipase. Eur. J. Pharm. 2006, 543, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Kamalipour, M.; Akhondzadeh, S. Cardiovascular effects of saffron: An evidence-based review. J. Tehran Heart Cent. 2011, 6, 59–61. [Google Scholar]
- He, S.Y.; Qian, Z.Y.; Tang, F.T.; Wen, N.; Xu, G.L.; Sheng, L. Effect of crocin on experimental atherosclerosis in quails and its mechanisms. Life Sci. 2005, 77, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Avila-Sosa, R.; Nevárez-Moorillón, G.V.; Ochoa-Velasco, C.E.; Navarro-Cruz, A.R.; Hernández-Carranza, P.; Cid-Pérez, T.S. Detection of Saffron’s Main Bioactive Compounds and Their Relationship with Commercial Quality. Foods 2022, 11, 3245. [Google Scholar] [CrossRef]
- Garavand, F.; Rahaee, S.; Vahedikia, N.; Jafari, S.M. Different techniques for extraction and micro/nanoencapsulation of saffron bioactive ingredients. Trends Food Sci. Technol. 2019, 89, 26–44. [Google Scholar] [CrossRef]
- Leoni, V.; Giupponi, L.; Pedrali, D.; Zuccolo, M.; Borgonovo, G.; Bassoli, A.; Giorgi, A. How harvest, cleaning and conservation good practices affect the quality of saffron: Results of a research conducted in Italy. J. Appl. Bot. Food Qual. 2022, 95, 9. [Google Scholar]
- Sobolev, A.P.; Carradori, S.; Capitani, D.; Vista, S.; Trella, A.; Marini, F.; Mannina, L. Saffron Samples of Different Origin: An NMR Study of Microwave-Assisted Extracts. Foods 2014, 3, 403–419. [Google Scholar] [CrossRef]
- Yilmaz, A.; Nyberg, N.T.; Mølgaard, P.; Asili, J.; Jaroszewski, J.W. 1H NMR metabolic fingerprinting of saffron extracts. Metabolomics 2010, 6, 511–517. [Google Scholar] [CrossRef]
- Curcio, R.; Aiello, D.; Vozza, A.; Muto, L.; Martello, E.; Cappello, A.R.; Capobianco, L.; Fiermonte, G.; Siciliano, C.; Napoli, A.; et al. Cloning, Purification, and Characterization of the Catalytic C-Terminal Domain of the Human 3-Hydroxy-3-methyl glutaryl-CoA Reductase: An Effective, Fast, and Easy Method for Testing Hypocholesterolemic Compounds. Mol. Biotechnol. 2020, 62, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Bonofiglio, D.; Santoro, A.; Martello, E.; Vizza, D.; Rovito, D.; Cappello, A.R.; Barone, I.; Giordano, C.; Panza, S.; Catalano, S.; et al. Mechanisms of divergent effects of activated peroxisome proliferator-activated receptor-gamma on mitochondrial citrate carrier expression in 3T3-L1 fibroblasts and mature adipocytes. Biochim. Biophys. Acta 2013, 1831, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Dolce, V.; Cappello, A.R.; Capobianco, L. Mitochondrial tricarboxylate and dicarboxylate–Tricarboxylate carriers: From animals to plants. IUBMB Life 2014, 66, 462–471. [Google Scholar] [CrossRef]
- Zingg, J.M.; Hasan, S.T.; Meydani, M. Molecular mechanisms of hypolipidemic effects of curcumin. Biofactors 2013, 39, 101–121. [Google Scholar] [CrossRef]
- Li, Z.; Votava, J.A.; Zajac, G.J.M.; Nguyen, J.N.; Leyva Jaimes, F.B.; Ly, S.M.; Brinkman, J.A.; De Giorgi, M.; Kaul, S.; Green, C.L.; et al. Integrating Mouse and Human Genetic Data to Move beyond GWAS and Identify Causal Genes in Cholesterol Metabolism. Cell Metab. 2020, 31, 741–754.e5. [Google Scholar] [CrossRef]
- Yang, H.X.; Zhang, M.; Long, S.Y.; Tuo, Q.H.; Tian, Y.; Chen, J.X.; Zhang, C.P.; Liao, D.F. Cholesterol in LDL receptor recycling and degradation. Clin. Chim. Acta 2020, 500, 81–86. [Google Scholar] [CrossRef]
- Cohen, L.H.; Van Vliet, A.; Roodenburg, L.; Jansen, L.M.C.; Griffigen, M. Pravastatin inhibited the cholesterol synthesis in human hepatoma cell line Hep G2 less than simvastatin and lovastatin, which is reflected in the upregulation of 3-hydroxy-3-methylglutaryl coenzyme a reductase and squalene synthase. Biochem. Pharmacol. 1993, 45, 2203–2208. [Google Scholar] [CrossRef]
- Kawata, S.; Nagase, T.; Yamasaki, E.; Ishiguro, H.; Matsuzawa, Y. Modulation of the mevalonate pathway and cell growth by pravastatin and d-limonene in a human hepatoma cell line (Hep G2). Br. J. Cancer 1994, 69, 1015–1020. [Google Scholar] [CrossRef]
- Bartolomei, M.; Bollati, C.; Li, J.; Arnoldi, A.; Lammi, C. Assessment of the Cholesterol-Lowering Effect of MOMAST®: Biochemical and Cellular Studies. Nutrients 2022, 14, 493. [Google Scholar] [PubMed]
- Panda, T.; Devi, V.A. Regulation and degradation of HMGCo-A reductase. Appl. Microbiol. Biotechnol. 2004, 66, 143–152. [Google Scholar] [CrossRef]
- Aguirre, L.; Hijona, E.; Macarulla, M.T.; Gracia, A.; Larrechi, I.; Bujanda, L.; Hijona, L.; Portillo, M.P. Several statins increase body and liver fat accumulation in a model of metabolic syndrome. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2013, 64, 281–288. [Google Scholar]
- Amemiya-Kudo, M.; Shimano, H.; Hasty, A.H.; Yahagi, N.; Yoshikawa, T.; Matsuzaka, T.; Okazaki, H.; Tamura, Y.; Iizuka, Y.; Ohashi, K.; et al. Transcriptional activities of nuclear SREBP-1a, -1c, and -2 to different target promoters of lipogenic and cholesterogenic genes. J. Lipid Res. 2002, 43, 1220–1235. [Google Scholar] [CrossRef]
- Lico, D.; Vuono, D.; Siciliano, C.; Nagy, J.B.; De Luca, P. Removal of unleaded gasoline from water by multi-walled carbon nanotubes. J. Environ. Manag. 2019, 237, 636–643. [Google Scholar] [CrossRef]
- Aiello, D.; Furia, E.; Siciliano, C.; Bongiorno, D.; Napoli, A. Study of the coordination of ortho-tyrosine and trans-4-hydroxyproline with aluminum(III) and iron(III). J. Mol. Liq. 2018, 269, 387–397. [Google Scholar] [CrossRef]
- Siciliano, C.; Belsito, E.; De Marco, R.; Di Gioia, M.L.; Leggio, A.; Liguori, A. Quantitative determination of fatty acid chain composition in pork meat products by high resolution 1H NMR spectroscopy. Food Chem. 2013, 136, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Aiello, D.; Siciliano, C.; Mazzotti, F.; Di Donna, L.; Athanassopoulos, C.M.; Napoli, A. Molecular species fingerprinting and quantitative analysis of saffron (Crocus sativus L.) for quality control by MALDI mass spectrometry. RSC Adv. 2018, 8, 36104–36113. [Google Scholar] [CrossRef]
- Aiello, D.; Siciliano, C.; Mazzotti, F.; Di Donna, L.; Athanassopoulos, C.M.; Napoli, A. A rapid MALDI MS/MS based method for assessing saffron (Crocus sativus L.) adulteration. Food Chem. 2020, 307, 125527. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, A.M.; Carmona, M.; del Campo, C.P.; Alonso, G.L. Solid-phase extraction for picrocrocin determination in the quality control of saffron spice (Crocus sativus L.). Food Chem. 2009, 116, 792–798. [Google Scholar] [CrossRef]
- Lappano, R.; Rosano, C.; Pisano, A.; Santolla, M.F.; De Francesco, E.M.; De Marco, P.; Dolce, V.; Ponassi, M.; Felli, L.; Cafeo, G.; et al. A calixpyrrole derivative acts as an antagonist to GPER, a G-protein coupled receptor: Mechanisms and models. Dis. Model. Mech. 2015, 8, 1237–1246. [Google Scholar] [CrossRef]
- Frattaruolo, L.; Fiorillo, M.; Brindisi, M.; Curcio, R.; Dolce, V.; Lacret, R.; Truman, A.W.; Sotgia, F.; Lisanti, M.P.; Cappello, A.R. Thioalbamide, A Thioamidated Peptide from Amycolatopsis alba, Affects Tumor Growth and Stemness by Inducing Metabolic Dysfunction and Oxidative Stress. Cells 2019, 8, 1408. [Google Scholar] [CrossRef] [PubMed]
- Carrisi, C.; Madeo, M.; Morciano, P.; Dolce, V.; Cenci, G.; Cappello, A.R.; Mazzeo, G.; Iacopetta, D.; Capobianco, L. Identification of the Drosophila melanogaster mitochondrial citrate carrier: Bacterial expression, reconstitution, functional characterization and developmental distribution. J. Biochem. 2008, 144, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Raho, S.; Capobianco, L.; Malivindi, R.; Vozza, A.; Piazzolla, C.; De Leonardis, F.; Gorgoglione, R.; Scarcia, P.; Pezzuto, F.; Agrimi, G.; et al. KRAS-regulated glutamine metabolism requires UCP2-mediated aspartate transport to support pancreatic cancer growth. Nat. Metab. 2020, 2, 1373–1381. [Google Scholar] [CrossRef]
- Mazzotta, S.; Frattaruolo, L.; Brindisi, M.; Ulivieri, C.; Vanni, F.; Brizzi, A.; Carullo, G.; Cappello, A.R.; Aiello, F. 3-Amino-alkylated indoles: Unexplored green products acting as anti-inflammatory agents. Future Med. Chem. 2020, 12, 5–17. [Google Scholar] [CrossRef]
- Scarcia, P.; Gorgoglione, R.; Messina, E.; Fiermonte, G.; Blank, L.M.; Wierckx, N.; Palmieri, L.; Agrimi, G. Mitochondrial carriers of Ustilago maydis and Aspergillus terreus involved in itaconate production: Same physiological role but different biochemical features. FEBS Lett. 2020, 594, 728–739. [Google Scholar] [CrossRef]
- Lunetti, P.; Gorgoglione, R.; Curcio, R.; Marra, F.; Pignataro, A.; Vozza, A.; Riley, C.L.; Capobianco, L.; Palmieri, L.; Dolce, V.; et al. Drosophila melanogaster Uncoupling Protein-4A (UCP4A) Catalyzes a Unidirectional Transport of Aspartate. Int. J. Mol. Sci. 2022, 23, 1020. [Google Scholar] [CrossRef] [PubMed]
- Lunetti, P.; Cappello, A.R.; Marsano, R.M.; Pierri, C.L.; Carrisi, C.; Martello, E.; Caggese, C.; Dolce, V.; Capobianco, L. Mitochondrial glutamate carriers from Drosophila melanogaster: Biochemical, evolutionary and modeling studies. Biochim. Biophys. Acta 2013, 1827, 1245–1255. [Google Scholar] [CrossRef]
- Lunetti, P.; Marsano, R.M.; Curcio, R.; Dolce, V.; Fiermonte, G.; Cappello, A.R.; Marra, F.; Moschetti, R.; Li, Y.; Aiello, D.; et al. The mitochondrial aspartate/glutamate carrier (AGC or Aralar1) isoforms in D. melanogaster: Biochemical characterization, gene structure, and evolutionary analysis. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129854. [Google Scholar] [CrossRef]
- Armentano, B.; Curcio, R.; Brindisi, M.; Mancuso, R.; Rago, V.; Ziccarelli, I.; Frattaruolo, L.; Fiorillo, M.; Dolce, V.; Gabriele, B.; et al. 5-(Carbamoylmethylene)-oxazolidin-2-ones as a Promising Class of Heterocycles Inducing Apoptosis Triggered by Increased ROS Levels and Mitochondrial Dysfunction in Breast and Cervical Cancer. Biomedicines 2020, 8, 35. [Google Scholar] [CrossRef]
- Profilo, E.; Pena-Altamira, L.E.; Corricelli, M.; Castegna, A.; Danese, A.; Agrimi, G.; Petralla, S.; Giannuzzi, G.; Porcelli, V.; Sbano, L.; et al. Down-regulation of the mitochondrial aspartate-glutamate carrier isoform 1 AGC1 inhibits proliferation and N-acetylaspartate synthesis in Neuro2A cells. Biochim. Biophys. Acta 2017, 1863, 1422–1435. [Google Scholar] [CrossRef]
- Curcio, R.; Muto, L.; Pierri, C.L.; Montalto, A.; Lauria, G.; Onofrio, A.; Fiorillo, M.; Fiermonte, G.; Lunetti, P.; Vozza, A.; et al. New insights about the structural rearrangements required for substrate translocation in the bovine mitochondrial oxoglutarate carrier. Biochim. Biophys. Acta 2016, 1864, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cappello, A.R.; Muto, L.; Martello, E.; Madeo, M.; Curcio, R.; Lunetti, P.; Raho, S.; Zaffino, F.; Frattaruolo, L.; et al. Functional characterization of the partially purified Sac1p independent adenine nucleotide transport system (ANTS) from yeast endoplasmic reticulum. J. Biochem. 2018, 164, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Avino, S.; De Marco, P.; Cirillo, F.; Santolla, M.F.; De Francesco, E.M.; Perri, M.G.; Rigiracciolo, D.; Dolce, V.; Belfiore, A.; Maggiolini, M.; et al. Stimulatory actions of IGF-I are mediated by IGF-IR cross-talk with GPER and DDR1 in mesothelioma and lung cancer cells. Oncotarget 2016, 7, 52710–52728. [Google Scholar] [CrossRef] [PubMed]
- Farnier, M.; Zeller, M.; Masson, D.; Cottin, Y. Triglycerides and risk of atherosclerotic cardiovascular disease: An update. Arch. Cardiovasc. Dis. 2021, 114, 132–139. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) |
|---|---|
| SREBP1-Fw | GCGGAGCCATGGATTGCAC |
| SREBP1-Rv | TCTTCCTTGATACCAGGCCC |
| SREBP2-Fw | TGGCTTCTCTCCCTACTCCA |
| SREBP2-Rv | GCAGCTGCAAAATCTCCTCT |
| FASN-Fw | AGCTGCCAGAGTCGGAGAAC |
| FASN-Rv | TGTAGCCCACGAGTGTCTCG |
| GPAT1-Fw | GGCATCCTGAACTGGTGTGTG |
| GPAT1-Rv | GAGCTTGAGGAAGAGGATGGTG |
| HMGR-Fw | AGGTTCCAATGGCAACAACAGAAG |
| HMGR-Rv | ATGCTCCTTGAACACCTAGCATCT |
| LDLR-Fw | CAATGTCTCACCAAGCTCTG |
| LDLR-Rv | TCTGTCTCGAGGGGTAGCTG |
| PPIA-Fw | CATACGGGTCCTGGCATCTT |
| PPIA-Rv | TCCATGGCCTCCACAATATTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frattaruolo, L.; Marra, F.; Lauria, G.; Siciliano, C.; Curcio, R.; Muto, L.; Brindisi, M.; Aiello, D.; Napoli, A.; Fiermonte, G.; et al. A Picrocrocin-Enriched Fraction from a Saffron Extract Affects Lipid Homeostasis in HepG2 Cells through a Non-Statin-like Mode. Int. J. Mol. Sci. 2023, 24, 3060. https://doi.org/10.3390/ijms24043060
Frattaruolo L, Marra F, Lauria G, Siciliano C, Curcio R, Muto L, Brindisi M, Aiello D, Napoli A, Fiermonte G, et al. A Picrocrocin-Enriched Fraction from a Saffron Extract Affects Lipid Homeostasis in HepG2 Cells through a Non-Statin-like Mode. International Journal of Molecular Sciences. 2023; 24(4):3060. https://doi.org/10.3390/ijms24043060
Chicago/Turabian StyleFrattaruolo, Luca, Federica Marra, Graziantonio Lauria, Carlo Siciliano, Rosita Curcio, Luigina Muto, Matteo Brindisi, Donatella Aiello, Anna Napoli, Giuseppe Fiermonte, and et al. 2023. "A Picrocrocin-Enriched Fraction from a Saffron Extract Affects Lipid Homeostasis in HepG2 Cells through a Non-Statin-like Mode" International Journal of Molecular Sciences 24, no. 4: 3060. https://doi.org/10.3390/ijms24043060
APA StyleFrattaruolo, L., Marra, F., Lauria, G., Siciliano, C., Curcio, R., Muto, L., Brindisi, M., Aiello, D., Napoli, A., Fiermonte, G., Cappello, A. R., Fiorillo, M., Ahmed, A., & Dolce, V. (2023). A Picrocrocin-Enriched Fraction from a Saffron Extract Affects Lipid Homeostasis in HepG2 Cells through a Non-Statin-like Mode. International Journal of Molecular Sciences, 24(4), 3060. https://doi.org/10.3390/ijms24043060