

MicroRNAs in the Mouse Developing Retina

Abstract
1. Biogenesis of microRNAs
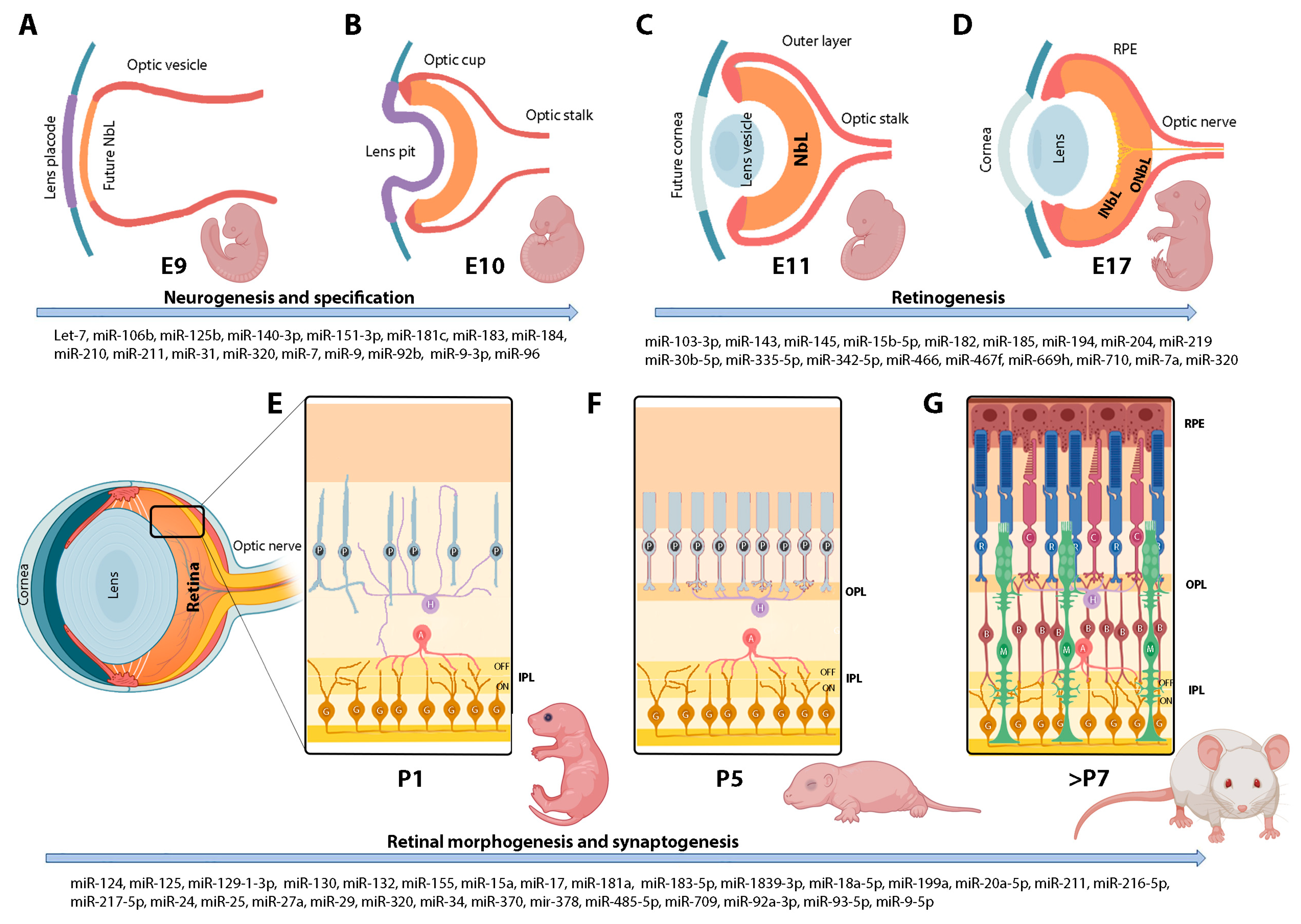
2. Brief Overview of the Retinal Development
2.1. Regulation of Retinal Neurogenesis and Specification
2.2. Regulation of the Morphology and Connectivity of Retinal Neurons
2.3. Regulation of Retinal Apoptosis
3. Retinal microRNAs in Development
3.1. Dissecting the Roles of microRNAs during Retinal Development
3.1.1. The miR-182/96/183 Cluster
3.1.2. The miR-204/211 Family
3.1.3. miR-124
3.1.4. miR-9
3.1.5. The Let-7 Family
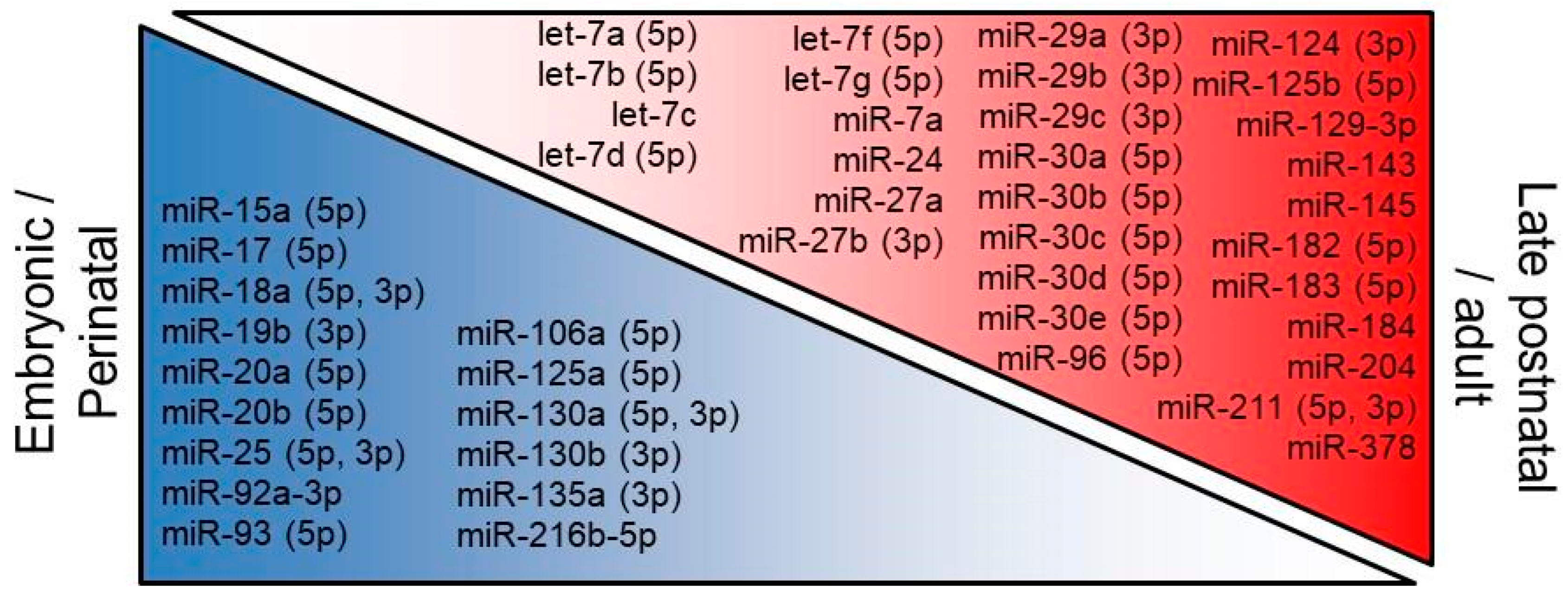
3.2. Developmental Regulation of Retinal microRNAs
4. Involvement of microRNAs in Retinal Pathologies
5. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hombach, S.; Kretz, M. Non-coding RNAs: Classification, Biology and Functioning. Adv. Exp. Med. Biol. 2016, 937, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Zuzic, M.; Rojo Arias, J.E.; Wohl, S.G.; Busskamp, V. Retinal miRNA Functions in Health and Disease. Genes 2019, 10, 377. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lee, Y.; Yeom, K.H.; Kim, Y.K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 complex in primary microRNA processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef] [PubMed]
- Westholm, J.O.; Lai, E.C. Mirtrons: microRNA biogenesis via splicing. Biochimie 2011, 93, 1897–1904. [Google Scholar] [CrossRef]
- Yi, R.; Qin, Y.; Macara, I.G.; Cullen, B.R. Exportin-5 mediates the nuclear export of pre-microRNAs and short hairpin RNAs. Genes Dev. 2003, 17, 3011–3016. [Google Scholar] [CrossRef] [PubMed]
- Hutvagner, G.; McLachlan, J.; Pasquinelli, A.E.; Balint, E.; Tuschl, T.; Zamore, P.D. A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNA. Science 2001, 293, 834–838. [Google Scholar] [CrossRef]
- Liu, J.; Carmell, M.A.; Rivas, F.V.; Marsden, C.G.; Thomson, J.M.; Song, J.J.; Hammond, S.M.; Joshua-Tor, L.; Hannon, G.J. Argonaute2 is the catalytic engine of mammalian RNAi. Science 2004, 305, 1437–1441. [Google Scholar] [CrossRef]
- Stenkamp, D.L. Development of the Vertebrate Eye and Retina. Prog. Mol. Biol. Transl. Sci. 2015, 134, 397–414. [Google Scholar] [CrossRef]
- Jeon, C.J.; Strettoi, E.; Masland, R.H. The major cell populations of the mouse retina. J. Neurosci. 1998, 18, 8936–8946. [Google Scholar] [CrossRef]
- Nadal-Nicolas, F.M.; Kunze, V.P.; Ball, J.M.; Peng, B.T.; Krishnan, A.; Zhou, G.; Dong, L.; Li, W. True S-cones are concentrated in the ventral mouse retina and wired for color detection in the upper visual field. Elife 2020, 9, e56840. [Google Scholar] [CrossRef] [PubMed]
- Byerly, M.S.; Blackshaw, S. Vertebrate retina and hypothalamus development. Wiley Interdiscip. Rev. Syst. Biol. Med. 2009, 1, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Bassett, E.A.; Wallace, V.A. Cell fate determination in the vertebrate retina. Trends Neurosci. 2012, 35, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Hoon, M.; Okawa, H.; Della Santina, L.; Wong, R.O. Functional architecture of the retina: Development and disease. Prog. Retin. Eye Res. 2014, 42, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Fuerst, P.G.; Bruce, F.; Tian, M.; Wei, W.; Elstrott, J.; Feller, M.B.; Erskine, L.; Singer, J.H.; Burgess, R.W. DSCAM and DSCAML1 function in self-avoidance in multiple cell types in the developing mouse retina. Neuron 2009, 64, 484–497. [Google Scholar] [CrossRef]
- Sun, L.O.; Jiang, Z.; Rivlin-Etzion, M.; Hand, R.; Brady, C.M.; Matsuoka, R.L.; Yau, K.W.; Feller, M.B.; Kolodkin, A.L. On and off retinal circuit assembly by divergent molecular mechanisms. Science 2013, 342, 1241974. [Google Scholar] [CrossRef]
- Lefebvre, J.L.; Kostadinov, D.; Chen, W.V.; Maniatis, T.; Sanes, J.R. Protocadherins mediate dendritic self-avoidance in the mammalian nervous system. Nature 2012, 488, 517–521. [Google Scholar] [CrossRef]
- Raven, M.A.; Oh, E.C.; Swaroop, A.; Reese, B.E. Afferent control of horizontal cell morphology revealed by genetic respecification of rods and cones. J. Neurosci. 2007, 27, 3540–3547. [Google Scholar] [CrossRef]
- Behrens, C.; Schubert, T.; Haverkamp, S.; Euler, T.; Berens, P. Connectivity map of bipolar cells and photoreceptors in the mouse retina. Elife 2016, 5, e20041. [Google Scholar] [CrossRef]
- Ribic, A.; Liu, X.; Crair, M.C.; Biederer, T. Structural organization and function of mouse photoreceptor ribbon synapses involve the immunoglobulin protein synaptic cell adhesion molecule 1. J. Comp. Neurol. 2014, 522, 900–920. [Google Scholar] [CrossRef]
- Omori, Y.; Araki, F.; Chaya, T.; Kajimura, N.; Irie, S.; Terada, K.; Muranishi, Y.; Tsujii, T.; Ueno, S.; Koyasu, T.; et al. Presynaptic dystroglycan-pikachurin complex regulates the proper synaptic connection between retinal photoreceptor and bipolar cells. J. Neurosci. 2012, 32, 6126–6137. [Google Scholar] [CrossRef] [PubMed]
- Soto, F.; Watkins, K.L.; Johnson, R.E.; Schottler, F.; Kerschensteiner, D. NGL-2 regulates pathway-specific neurite growth and lamination, synapse formation, and signal transmission in the retina. J. Neurosci. 2013, 33, 11949–11959. [Google Scholar] [CrossRef]
- Brandstätter, J.H.; Fletcher, E.L.; Garner, C.C.; Gundelfinger, E.D.; Wässle, H. Differential expression of the presynaptic cytomatrix protein bassoon among ribbon synapses in the mammalian retina. Eur. J. Neurosci. 1999, 11, 3683–3693. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.L.; Schubert, T.; Wong, R.O. Developmental patterning of glutamatergic synapses onto retinal ganglion cells. Neural Dev. 2008, 3, 8. [Google Scholar] [CrossRef]
- Fisher, L.J. Development of synaptic arrays in the inner plexiform layer of neonatal mouse retina. J. Comp. Neurol. 1979, 187, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Murcia-Belmonte, V.; Erskine, L. Wiring the Binocular Visual Pathways. Int. J. Mol. Sci. 2019, 20, 3282. [Google Scholar] [CrossRef]
- Herrera, E.; Erskine, L.; Morenilla-Palao, C. Guidance of retinal axons in mammals. Semin. Cell Dev. Biol. 2019, 85, 48–59. [Google Scholar] [CrossRef]
- Prieur, D.S.; Rebsam, A. Retinal axon guidance at the midline: Chiasmatic misrouting and consequences. Dev. Neurobiol. 2017, 77, 844–860. [Google Scholar] [CrossRef]
- Cang, J.; Rentería, R.C.; Kaneko, M.; Liu, X.; Stryker, M.P. Development of precise maps in visual cortex requires patterned spontaneous activity in the retina. Neuron 2005, 48, 797–809. [Google Scholar] [CrossRef]
- Davies, A.M. Regulation of neuronal survival and death by extracellular signals during development. EMBO J. 2003, 22, 2537–2545. [Google Scholar] [CrossRef]
- Donovan, M.; Doonan, F.; Cotter, T.G. Decreased expression of pro-apoptotic Bcl-2 family members during retinal development and differential sensitivity to cell death. Dev. Biol. 2006, 291, 154–169. [Google Scholar] [CrossRef] [PubMed]
- O’Driscoll, C.; Donovan, M.; Cotter, T.G. Analysis of apoptotic and survival mediators in the early post-natal and mature retina. Exp. Eye Res. 2006, 83, 1482–1492. [Google Scholar] [CrossRef] [PubMed]
- Wahlin, K.J.; Enke, R.A.; Fuller, J.A.; Kalesnykas, G.; Zack, D.J.; Merbs, S.L. Epigenetics and cell death: DNA hypermethylation in programmed retinal cell death. PLoS ONE 2013, 8, e79140. [Google Scholar] [CrossRef]
- Wallace, D.M.; Donovan, M.; Cotter, T.G. Histone deacetylase activity regulates apaf-1 and caspase 3 expression in the developing mouse retina. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2765–2772. [Google Scholar] [CrossRef] [PubMed]
- Sundermeier, T.R.; Palczewski, K. The impact of microRNA gene regulation on the survival and function of mature cell types in the eye. FASEB J. 2016, 30, 23–33. [Google Scholar] [CrossRef]
- Karali, M.; Persico, M.; Mutarelli, M.; Carissimo, A.; Pizzo, M.; Singh Marwah, V.; Ambrosio, C.; Pinelli, M.; Carrella, D.; Ferrari, S.; et al. High-resolution analysis of the human retina miRNome reveals isomiR variations and novel microRNAs. Nucleic Acids Res. 2016, 44, 1525–1540. [Google Scholar] [CrossRef]
- Pawlick, J.S.; Zuzic, M.; Pasquini, G.; Swiersy, A.; Busskamp, V. MiRNA Regulatory Functions in Photoreceptors. Front. Cell Dev. Biol. 2020, 8, 620249. [Google Scholar] [CrossRef]
- Georgi, S.A.; Reh, T.A. Dicer is required for the transition from early to late progenitor state in the developing mouse retina. J. Neurosci. 2010, 30, 4048–4061. [Google Scholar] [CrossRef]
- La Torre, A.; Georgi, S.; Reh, T.A. Conserved microRNA pathway regulates developmental timing of retinal neurogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, E2362–E2370. [Google Scholar] [CrossRef]
- Wohl, S.G.; Hooper, M.J.; Reh, T.A. MicroRNAs miR-25, let-7 and miR-124 regulate the neurogenic potential of Muller glia in mice. Development 2019, 146, dev179556. [Google Scholar] [CrossRef]
- Baba, Y.; Aihara, Y.; Watanabe, S. MicroRNA-7a regulates Muller glia differentiation by attenuating Notch3 expression. Exp. Eye Res. 2015, 138, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Witmer, P.D.; Lumayag, S.; Kovacs, B.; Valle, D. MicroRNA (miRNA) transcriptome of mouse retina and identification of a sensory organ-specific miRNA cluster. J. Biol. Chem. 2007, 282, 25053–25066. [Google Scholar] [CrossRef]
- Arora, A.; Guduric-Fuchs, J.; Harwood, L.; Dellett, M.; Cogliati, T.; Simpson, D.A. Prediction of microRNAs affecting mRNA expression during retinal development. BMC Dev. Biol. 2010, 10, 1. [Google Scholar] [CrossRef]
- Wohl, S.G.; Reh, T.A. The microRNA expression profile of mouse Muller glia in vivo and in vitro. Sci. Rep. 2016, 6, 35423. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, F.; Okuno, M.; Tanaka, T.; Sanuki, R. Overexpression of neural miRNAs miR-9/9* and miR-124 suppresses differentiation to Muller glia and promotes differentiation to neurons in mouse retina in vivo. Genes Cells 2020, 25, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; Jiang, Y.; Liu, R.; Cao, D.; Pan, J.; Luo, Y. Identification of key miRNAs and genes for mouse retinal development using a linear model. Mol. Med. Rep. 2020, 22, 494–506. [Google Scholar] [CrossRef]
- Guan, J.T.; Li, X.X.; Peng, D.W.; Zhang, W.M.; Qu, J.; Lu, F.; D’Amato, R.J.; Chi, Z.L. MicroRNA-18a-5p Administration Suppresses Retinal Neovascularization by Targeting FGF1 and HIF1A. Front. Pharmacol. 2020, 11, 276. [Google Scholar] [CrossRef]
- Morris, D.R.; Bounds, S.E.; Liu, H.; Ding, W.Q.; Chen, Y.; Liu, Y.; Cai, J. Exosomal MiRNA Transfer between Retinal Microglia and RPE. Int. J. Mol. Sci. 2020, 21, 3541. [Google Scholar] [CrossRef] [PubMed]
- Hackler, L., Jr.; Wan, J.; Swaroop, A.; Qian, J.; Zack, D.J. MicroRNA profile of the developing mouse retina. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1823–1831. [Google Scholar] [CrossRef]
- Zhang, Y.; Shen, B.; Zhang, D.; Wang, Y.; Tang, Z.; Ni, N.; Jin, X.; Luo, M.; Sun, H.; Gu, P. miR-29a regulates the proliferation and differentiation of retinal progenitors by targeting Rbm8a. Oncotarget 2017, 8, 31993–32008. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, R.; Hong, Y.; Wang, Y.; Zhu, Y.; Wen, T.; Lu, J.; Mao, S.; Wang, X.; Pan, J.; et al. MicroRNA-92a-3p Regulates Retinal Angiogenesis by Targeting SGK3 in Vascular Endothelial Cells. Investig. Ophthalmol. Vis. Sci. 2022, 63, 19. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Zhang, J.; Rao, F.Q.; Yang, Q.L.; Zeng, H.Y.; Huang, S.H.; Xie, Z.X.; Lv, J.N.; Lin, D.; Chen, X.J.; et al. Depletion of miR-96 Delays, but Does Not Arrest, Photoreceptor Development in Mice. Investig. Ophthalmol. Vis. Sci. 2022, 63, 24. [Google Scholar] [CrossRef] [PubMed]
- Sanuki, R.; Onishi, A.; Koike, C.; Muramatsu, R.; Watanabe, S.; Muranishi, Y.; Irie, S.; Uneo, S.; Koyasu, T.; Matsui, R.; et al. miR-124a is required for hippocampal axogenesis and retinal cone survival through Lhx2 suppression. Nat. Neurosci. 2011, 14, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Marler, K.J.; Suetterlin, P.; Dopplapudi, A.; Rubikaite, A.; Adnan, J.; Maiorano, N.A.; Lowe, A.S.; Thompson, I.D.; Pathania, M.; Bordey, A.; et al. BDNF promotes axon branching of retinal ganglion cells via miRNA-132 and p250GAP. J. Neurosci. 2014, 34, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Sreekanth, S.; Rasheed, V.A.; Soundararajan, L.; Antony, J.; Saikia, M.; Sivakumar, K.C.; Das, A.V. miR Cluster 143/145 Directly Targets Nrl and Regulates Rod Photoreceptor Development. Mol. Neurobiol. 2017, 54, 8033–8049. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.H.; Wang, Z.; Huang, S.; Sun, Y.; Chen, J. MicroRNA-145 Regulates Pathological Retinal Angiogenesis by Suppression of TMOD3. Mol. Ther. Nucleic Acids 2019, 16, 335–347. [Google Scholar] [CrossRef]
- Dong, Y.; Alonso, F.; Jahjah, T.; Fremaux, I.; Grosset, C.F.; Genot, E. miR-155 regulates physiological angiogenesis but an miR-155-rich microenvironment disrupts the process by promoting unproductive endothelial sprouting. Cell. Mol. Life Sci. 2022, 79, 208. [Google Scholar] [CrossRef]
- Wu, K.C.; Chen, X.J.; Jin, G.H.; Wang, X.Y.; Yang, D.D.; Li, Y.P.; Xiang, L.; Zhang, B.W.; Zhou, G.H.; Zhang, C.J.; et al. Deletion of miR-182 Leads to Retinal Dysfunction in Mice. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1265–1274. [Google Scholar] [CrossRef]
- Zhang, C.J.; Xiang, L.; Chen, X.J.; Wang, X.Y.; Wu, K.C.; Zhang, B.W.; Chen, D.F.; Jin, G.H.; Zhang, H.; Chen, Y.C.; et al. Ablation of Mature miR-183 Leads to Retinal Dysfunction in Mice. Investig. Ophthalmol. Vis. Sci. 2020, 61, 12. [Google Scholar] [CrossRef]
- Barbato, S.; Marrocco, E.; Intartaglia, D.; Pizzo, M.; Asteriti, S.; Naso, F.; Falanga, D.; Bhat, R.S.; Meola, N.; Carissimo, A.; et al. MiR-211 is essential for adult cone photoreceptor maintenance and visual function. Sci. Rep. 2017, 7, 17004. [Google Scholar] [CrossRef]
- Zhang, H.; Zhuang, P.; Welchko, R.M.; Dai, M.; Meng, F.; Turner, D.L. Regulation of retinal amacrine cell generation by miR-216b and Foxn3. Development 2022, 149, dev199484. [Google Scholar] [CrossRef]
- Chen, G.; Qian, H.M.; Chen, J.; Wang, J.; Guan, J.T.; Chi, Z.L. Whole transcriptome sequencing identifies key circRNAs, lncRNAs, and miRNAs regulating neurogenesis in developing mouse retina. BMC Genom. 2021, 22, 779. [Google Scholar] [CrossRef]
- Fishman, E.S.; Louie, M.; Miltner, A.M.; Cheema, S.K.; Wong, J.; Schlaeger, N.M.; Moshiri, A.; Simo, S.; Tarantal, A.F.; La Torre, A. MicroRNA Signatures of the Developing Primate Fovea. Front. Cell Dev. Biol. 2021, 9, 654385. [Google Scholar] [CrossRef] [PubMed]
- Kabekkodu, S.P.; Shukla, V.; Varghese, V.K.; D’Souza, J.; Chakrabarty, S.; Satyamoorthy, K. Clustered miRNAs and their role in biological functions and diseases. Biol. Rev. Camb. Philos. Soc. 2018, 93, 1955–1986. [Google Scholar] [CrossRef] [PubMed]
- Dambal, S.; Shah, M.; Mihelich, B.; Nonn, L. The microRNA-183 cluster: The family that plays together stays together. Nucleic Acids Res. 2015, 43, 7173–7188. [Google Scholar] [CrossRef] [PubMed]
- Jalvy-Delvaille, S.; Maurel, M.; Majo, V.; Pierre, N.; Chabas, S.; Combe, C.; Rosenbaum, J.; Sagliocco, F.; Grosset, C.F. Molecular basis of differential target regulation by miR-96 and miR-182: The Glypican-3 as a model. Nucleic Acids Res. 2012, 40, 1356–1365. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Du, X.; Tai, S.; Zhong, X.; Wang, Z.; Hu, Z.; Zhang, L.; Kang, P.; Ji, D.; Jiang, X.; et al. GPC1 regulated by miR-96-5p, rather than miR-182-5p, in inhibition of pancreatic carcinoma cell proliferation. Int. J. Mol. Sci. 2014, 15, 6314–6327. [Google Scholar] [CrossRef]
- Krol, J.; Krol, I.; Alvarez, C.P.; Fiscella, M.; Hierlemann, A.; Roska, B.; Filipowicz, W. A network comprising short and long noncoding RNAs and RNA helicase controls mouse retina architecture. Nat. Commun. 2015, 6, 7305. [Google Scholar] [CrossRef]
- Lumayag, S.; Haldin, C.E.; Corbett, N.J.; Wahlin, K.J.; Cowan, C.; Turturro, S.; Larsen, P.E.; Kovacs, B.; Witmer, P.D.; Valle, D.; et al. Inactivation of the microRNA-183/96/182 cluster results in syndromic retinal degeneration. Proc. Natl. Acad. Sci. USA 2013, 110, E507–E516. [Google Scholar] [CrossRef]
- Jin, Z.B.; Hirokawa, G.; Gui, L.; Takahashi, R.; Osakada, F.; Hiura, Y.; Takahashi, M.; Yasuhara, O.; Iwai, N. Targeted deletion of miR-182, an abundant retinal microRNA. Mol. Vis. 2009, 15, 523–533. [Google Scholar]
- Peskova, L.; Jurcikova, D.; Vanova, T.; Krivanek, J.; Capandova, M.; Sramkova, Z.; Sebestikova, J.; Kolouskova, M.; Kotasova, H.; Streit, L.; et al. miR-183/96/182 cluster is an important morphogenetic factor targeting PAX6 expression in differentiating human retinal organoids. Stem Cells 2020, 38, 1557–1567. [Google Scholar] [CrossRef]
- Shaham, O.; Gueta, K.; Mor, E.; Oren-Giladi, P.; Grinberg, D.; Xie, Q.; Cvekl, A.; Shomron, N.; Davis, N.; Keydar-Prizant, M.; et al. Pax6 regulates gene expression in the vertebrate lens through miR-204. PLoS Genet. 2013, 9, e1003357. [Google Scholar] [CrossRef] [PubMed]
- Bereimipour, A.; Najafi, H.; Mirsane, E.S.; Moradi, S.; Satarian, L. Roles of miR-204 in retinal development and maintenance. Exp. Cell Res. 2021, 406, 112737. [Google Scholar] [CrossRef] [PubMed]
- Lagos-Quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of tissue-specific microRNAs from mouse. Curr. Biol. 2002, 12, 735–739. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Luo, Z.M.; Guo, X.M.; Su, D.F.; Liu, X. An updated role of microRNA-124 in central nervous system disorders: A review. Front. Cell. Neurosci. 2015, 9, 193. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.B.; Murakami, Y.; Miller, J.W.; Vavvas, D.G. Neuroprotection for Age-Related Macular Degeneration. Ophthalmol. Sci. 2022, 2, 100192. [Google Scholar] [CrossRef]
- Alwin Prem Anand, A.; Huber, C.; Asnet Mary, J.; Gallus, N.; Leucht, C.; Klafke, R.; Hirt, B.; Wizenmann, A. Expression and function of microRNA-9 in the mid-hindbrain area of embryonic chick. BMC Dev. Biol. 2018, 18, 3. [Google Scholar] [CrossRef]
- Zhao, C.; Sun, G.; Li, S.; Shi, Y. A feedback regulatory loop involving microRNA-9 and nuclear receptor TLX in neural stem cell fate determination. Nat. Struct. Mol. Biol. 2009, 16, 365–371. [Google Scholar] [CrossRef]
- Hu, Y.; Luo, M.; Ni, N.; Den, Y.; Xia, J.; Chen, J.; Ji, J.; Zhou, X.; Fan, X.; Gu, P. Reciprocal actions of microRNA-9 and TLX in the proliferation and differentiation of retinal progenitor cells. Stem Cells Dev. 2014, 23, 2771–2781. [Google Scholar] [CrossRef]
- Chen, M.L.; Hong, C.G.; Yue, T.; Li, H.M.; Duan, R.; Hu, W.B.; Cao, J.; Wang, Z.X.; Chen, C.Y.; Hu, X.K.; et al. Inhibition of miR-331-3p and miR-9-5p ameliorates Alzheimer’s disease by enhancing autophagy. Theranostics 2021, 11, 2395–2409. [Google Scholar] [CrossRef]
- Hawley, Z.C.E.; Campos-Melo, D.; Strong, M.J. MiR-105 and miR-9 regulate the mRNA stability of neuronal intermediate filaments. Implications for the pathogenesis of amyotrophic lateral sclerosis (ALS). Brain Res. 2019, 1706, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Packer, A.N.; Xing, Y.; Harper, S.Q.; Jones, L.; Davidson, B.L. The bifunctional microRNA miR-9/miR-9* regulates REST and CoREST and is downregulated in Huntington’s disease. J. Neurosci. 2008, 28, 14341–14346. [Google Scholar] [CrossRef]
- Pena, J.T.; Sohn-Lee, C.; Rouhanifard, S.H.; Ludwig, J.; Hafner, M.; Mihailovic, A.; Lim, C.; Holoch, D.; Berninger, P.; Zavolan, M.; et al. miRNA in situ hybridization in formaldehyde and EDC-fixed tissues. Nat. Methods 2009, 6, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Roush, S.; Slack, F.J. The let-7 family of microRNAs. Trends Cell Biol. 2008, 18, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, H.; Song, H.; Qin, Y.; Wang, Y.; Xu, M.; Liu, C.; Gao, J.; Sun, S. Let-7d microRNA Attenuates 6-OHDA-Induced Injury by Targeting Caspase-3 in MN9D Cells. J. Mol. Neurosci. 2017, 63, 403–411. [Google Scholar] [CrossRef]
- Jiang, S.; Yan, W.; Wang, S.E.; Baltimore, D. Let-7 Suppresses B Cell Activation through Restricting the Availability of Necessary Nutrients. Cell Metab. 2018, 27, 393–403.e394. [Google Scholar] [CrossRef]
- Wells, A.C.; Daniels, K.A.; Angelou, C.C.; Fagerberg, E.; Burnside, A.S.; Markstein, M.; Alfandari, D.; Welsh, R.M.; Pobezinskaya, E.L.; Pobezinsky, L.A. Modulation of let-7 miRNAs controls the differentiation of effector CD8 T cells. Elife 2017, 6, e26398. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Q.; Yi, S.; Liu, Q.; Zhang, R.; Wang, P.; Qian, T.; Li, S. The microRNAs let-7 and miR-9 down-regulate the axon-guidance genes Ntn1 and Dcc during peripheral nerve regeneration. J. Biol. Chem. 2019, 294, 3489–3500. [Google Scholar] [CrossRef]
- Wang, X.W.; Li, Q.; Liu, C.M.; Hall, P.A.; Jiang, J.J.; Katchis, C.D.; Kang, S.; Dong, B.C.; Li, S.; Zhou, F.Q. Lin28 Signaling Supports Mammalian PNS and CNS Axon Regeneration. Cell Rep. 2018, 24, 2540–2552.e2546. [Google Scholar] [CrossRef]
- Sinigaglia, K.; Wiatrek, D.; Khan, A.; Michalik, D.; Sambrani, N.; Sedmik, J.; Vukic, D.; O’Connell, M.A.; Keegan, L.P. ADAR RNA editing in innate immune response phasing, in circadian clocks and in sleep. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 356–369. [Google Scholar] [CrossRef]
- Rom, S.; Dykstra, H.; Zuluaga-Ramirez, V.; Reichenbach, N.L.; Persidsky, Y. miR-98 and let-7g* protect the blood-brain barrier under neuroinflammatory conditions. J. Cereb. Blood Flow Metab. 2015, 35, 1957–1965. [Google Scholar] [CrossRef] [PubMed]
- Benhamed, M.; Herbig, U.; Ye, T.; Dejean, A.; Bischof, O. Senescence is an endogenous trigger for microRNA-directed transcriptional gene silencing in human cells. Nat. Cell Biol. 2012, 14, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Schwamborn, J.C.; Berezikov, E.; Knoblich, J.A. The TRIM-NHL protein TRIM32 activates microRNAs and prevents self-renewal in mouse neural progenitors. Cell 2009, 136, 913–925. [Google Scholar] [CrossRef] [PubMed]
- Fairchild, C.L.A.; Cheema, S.K.; Wong, J.; Hino, K.; Simo, S.; La Torre, A. Let-7 regulates cell cycle dynamics in the developing cerebral cortex and retina. Sci. Rep. 2019, 9, 15336. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, E.; Kim, S.Y.; Carmell, M.A.; Murchison, E.P.; Alcorn, H.; Li, M.Z.; Mills, A.A.; Elledge, S.J.; Anderson, K.V.; Hannon, G.J. Dicer is essential for mouse development. Nat. Genet. 2003, 35, 215–217. [Google Scholar] [CrossRef]
- Fiorenza, A.; Barco, A. Role of Dicer and the miRNA system in neuronal plasticity and brain function. Neurobiol. Learn. Mem. 2016, 135, 3–12. [Google Scholar] [CrossRef]
- Herrera-Carrillo, E.; Berkhout, B. Dicer-independent processing of small RNA duplexes: Mechanistic insights and applications. Nucleic Acids Res. 2017, 45, 10369–10379. [Google Scholar] [CrossRef]
- Damiani, D.; Alexander, J.J.; O’Rourke, J.R.; McManus, M.; Jadhav, A.P.; Cepko, C.L.; Hauswirth, W.W.; Harfe, B.D.; Strettoi, E. Dicer inactivation leads to progressive functional and structural degeneration of the mouse retina. J. Neurosci. 2008, 28, 4878–4887. [Google Scholar] [CrossRef]
- Sundermeier, T.R.; Zhang, N.; Vinberg, F.; Mustafi, D.; Kohno, H.; Golczak, M.; Bai, X.; Maeda, A.; Kefalov, V.J.; Palczewski, K. DICER1 is essential for survival of postmitotic rod photoreceptor cells in mice. FASEB J. 2014, 28, 3780–3791. [Google Scholar] [CrossRef]
- Aldunate, E.Z.; Di Foggia, V.; Di Marco, F.; Hervas, L.A.; Ribeiro, J.C.; Holder, D.L.; Patel, A.; Jannini, T.B.; Thompson, D.A.; Martinez-Barbera, J.P.; et al. Conditional Dicer1 depletion using Chrnb4-Cre leads to cone cell death and impaired photopic vision. Sci. Rep. 2019, 9, 2314. [Google Scholar] [CrossRef]
- Pinter, R.; Hindges, R. Perturbations of microRNA function in mouse dicer mutants produce retinal defects and lead to aberrant axon pathfinding at the optic chiasm. PLoS ONE 2010, 5, e10021. [Google Scholar] [CrossRef] [PubMed]
- Haider, N.B.; Mollema, N.; Gaule, M.; Yuan, Y.; Sachs, A.J.; Nystuen, A.M.; Naggert, J.K.; Nishina, P.M. Nr2e3-directed transcriptional regulation of genes involved in photoreceptor development and cell-type specific phototransduction. Exp. Eye Res. 2009, 89, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Haider, N.B.; Demarco, P.; Nystuen, A.M.; Huang, X.; Smith, R.S.; McCall, M.A.; Naggert, J.K.; Nishina, P.M. The transcription factor Nr2e3 functions in retinal progenitors to suppress cone cell generation. Vis. Neurosci. 2006, 23, 917–929. [Google Scholar] [CrossRef] [PubMed]
- Corbo, J.C.; Cepko, C.L. A hybrid photoreceptor expressing both rod and cone genes in a mouse model of enhanced S-cone syndrome. PLoS Genet. 2005, 1, e11. [Google Scholar] [CrossRef]
- Fujieda, H.; Bremner, R.; Mears, A.J.; Sasaki, H. Retinoic acid receptor-related orphan receptor alpha regulates a subset of cone genes during mouse retinal development. J. Neurochem. 2009, 108, 91–101. [Google Scholar] [CrossRef]
- Olivares, A.M.; Jelcick, A.S.; Reinecke, J.; Leehy, B.; Haider, A.; Morrison, M.A.; Cheng, L.; Chen, D.F.; DeAngelis, M.M.; Haider, N.B. Multimodal Regulation Orchestrates Normal and Complex Disease States in the Retina. Sci. Rep. 2017, 7, 690. [Google Scholar] [CrossRef]
- Dai, X.; Rao, C.; Li, H.; Chen, Y.; Fan, L.; Geng, H.; Li, S.; Qu, J.; Hou, L. Regulation of pigmentation by microRNAs: MITF-dependent microRNA-211 targets TGF-beta receptor 2. Pigment Cell Melanoma Res. 2015, 28, 217–222. [Google Scholar] [CrossRef]
- Moore, S.M.; Skowronska-Krawczyk, D.; Chao, D.L. Targeting of the NRL Pathway as a Therapeutic Strategy to Treat Retinitis Pigmentosa. J. Clin. Med. 2020, 9, 2224. [Google Scholar] [CrossRef]
- Fujitani, Y.; Fujitani, S.; Luo, H.; Qiu, F.; Burlison, J.; Long, Q.; Kawaguchi, Y.; Edlund, H.; MacDonald, R.J.; Furukawa, T.; et al. Ptf1a determines horizontal and amacrine cell fates during mouse retinal development. Development 2006, 133, 4439–4450. [Google Scholar] [CrossRef]
- Conte, I.; Hadfield, K.D.; Barbato, S.; Carrella, S.; Pizzo, M.; Bhat, R.S.; Carissimo, A.; Karali, M.; Porter, L.F.; Urquhart, J.; et al. MiR-204 is responsible for inherited retinal dystrophy associated with ocular coloboma. Proc. Natl. Acad. Sci. USA 2015, 112, E3236–E3245. [Google Scholar] [CrossRef]
- Kim, H.; Kim, J.; Ryu, J. Noncoding RNAs as a novel approach to target retinopathy of prematurity. Front. Pharmacol. 2022, 13, 1033341. [Google Scholar] [CrossRef] [PubMed]
- Ko, G.Y.; Yu, F.; Bayless, K.J.; Ko, M.L. MicroRNA-150 (miR-150) and Diabetic Retinopathy: Is miR-150 Only a Biomarker or Does It Contribute to Disease Progression? Int. J. Mol. Sci. 2022, 23, 12099. [Google Scholar] [CrossRef] [PubMed]
- Lande, K.; Gupta, J.; Ranjan, R.; Kiran, M.; Torres Solis, L.F.; Solis Herrera, A.; Aliev, G.; Karnati, R. Exosomes: Insights from Retinoblastoma and Other Eye Cancers. Int. J. Mol. Sci. 2020, 21, 7055. [Google Scholar] [CrossRef] [PubMed]
- Moazzeni, H.; Khani, M.; Elahi, E. Insights into the regulatory molecules involved in glaucoma pathogenesis. Am. J. Med. Genet. Part C Semin. Med. Genet. 2020, 184, 782–827. [Google Scholar] [CrossRef]
- Jun, S.; Datta, S.; Wang, L.; Pegany, R.; Cano, M.; Handa, J.T. The impact of lipids, lipid oxidation, and inflammation on AMD, and the potential role of miRNAs on lipid metabolism in the RPE. Exp. Eye Res. 2019, 181, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Gallenga, C.E.; Lonardi, M.; Pacetti, S.; Violanti, S.S.; Tassinari, P.; Di Virgilio, F.; Tognon, M.; Perri, P. Molecular Mechanisms Related to Oxidative Stress in Retinitis Pigmentosa. Antioxidants 2021, 10, 848. [Google Scholar] [CrossRef]
- Wei, Y.; Li, N.; Zhao, L.; Yang, C.; Ma, B.; Li, X.; Wei, R.; Nian, H. MicroRNAs and Autoimmune-Mediated Eye Diseases. Front. Cell Dev. Biol. 2020, 8, 818. [Google Scholar] [CrossRef]
- Kehl, T.; Backes, C.; Kern, F.; Fehlmann, T.; Ludwig, N.; Meese, E.; Lenhof, H.P.; Keller, A. About miRNAs, miRNA seeds, target genes and target pathways. Oncotarget 2017, 8, 107167–107175. [Google Scholar] [CrossRef]
- Kern, F.; Aparicio-Puerta, E.; Li, Y.; Fehlmann, T.; Kehl, T.; Wagner, V.; Ray, K.; Ludwig, N.; Lenhof, H.P.; Meese, E.; et al. miRTargetLink 2.0-interactive miRNA target gene and target pathway networks. Nucleic Acids Res. 2021, 49, W409–W416. [Google Scholar] [CrossRef]
- Narayanan, A.; Hill-Teran, G.; Moro, A.; Ristori, E.; Kasper, D.M.; Roden, C.A.; Lu, J.; Nicoli, S. In vivo mutagenesis of miRNA gene families using a scalable multiplexed CRISPR/Cas9 nuclease system. Sci. Rep. 2016, 6, 32386. [Google Scholar] [CrossRef]
- Ribeiro, A.O.; de Oliveira, A.C.; Costa, J.M.; Nachtigall, P.G.; Herkenhoff, M.E.; Campos, V.F.; Delella, F.K.; Pinhal, D. MicroRNA roles in regeneration: Multiple lessons from zebrafish. Dev. Dyn. 2022, 251, 556–576. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.M.; Giuffre, E.; Moseley, P.; Hitchcock, P.F. The MicroRNA, miR-18a, Regulates NeuroD and Photoreceptor Differentiation in the Retina of Zebrafish. Dev. Neurobiol. 2019, 79, 202–219. [Google Scholar] [CrossRef] [PubMed]
- Madelaine, R.; Sloan, S.A.; Huber, N.; Notwell, J.H.; Leung, L.C.; Skariah, G.; Halluin, C.; Pasca, S.P.; Bejerano, G.; Krasnow, M.A.; et al. MicroRNA-9 Couples Brain Neurogenesis and Angiogenesis. Cell Rep. 2017, 20, 1533–1542. [Google Scholar] [CrossRef]
- Sun, L.; Li, H.; Xu, X.; Xiao, G.; Luo, C. MicroRNA-20a is essential for normal embryogenesis by targeting vsx1 mRNA in fish. RNA Biol. 2015, 12, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Olena, A.F.; Rao, M.B.; Thatcher, E.J.; Wu, S.Y.; Patton, J.G. miR-216a regulates snx5, a novel notch signaling pathway component, during zebrafish retinal development. Dev. Biol. 2015, 400, 72–81. [Google Scholar] [CrossRef]
- Yamagata, M.; Yan, W.; Sanes, J.R. A cell atlas of the chick retina based on single-cell transcriptomics. Elife 2021, 10, e63907. [Google Scholar] [CrossRef]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly Parallel Genome-wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Dharmat, R.; Owen, L.; Shakoor, A.; Li, Y.; Kim, S.; Vitale, A.; Kim, I.; Morgan, D.; Liang, S.; et al. Single-nuclei RNA-seq on human retinal tissue provides improved transcriptome profiling. Nat. Commun. 2019, 10, 5743. [Google Scholar] [CrossRef]
- Pandey, R.S.; Krebs, M.P.; Bolisetty, M.T.; Charette, J.R.; Naggert, J.K.; Robson, P.; Nishina, P.M.; Carter, G.W. Single-Cell RNA Sequencing Reveals Molecular Features of Heterogeneity in the Murine Retinal Pigment Epithelium. Int. J. Mol. Sci. 2022, 23, 10419. [Google Scholar] [CrossRef]
- Youkilis, J.C.; Bassnett, S. Single-cell RNA-sequencing analysis of the ciliary epithelium and contiguous tissues in the mouse eye. Exp. Eye Res. 2021, 213, 108811. [Google Scholar] [CrossRef]
- Lyu, P.; Hoang, T.; Santiago, C.P.; Thomas, E.D.; Timms, A.E.; Appel, H.; Gimmen, M.; Le, N.; Jiang, L.; Kim, D.W.; et al. Gene regulatory networks controlling temporal patterning, neurogenesis, and cell-fate specification in mammalian retina. Cell Rep. 2021, 37, 109994. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| miRNA | Evidence | Experimental Approach | Role | Validated Targets | Experimental Target Validation | Ref. |
|---|---|---|---|---|---|---|
| let-7a | Decreased expression in Dicer1 cKO (αPax6cre; R26EYFP; Dicerfl/fl) | RT-qPCR | Dicer-dependent miRNA, part of the transition from early to late progenitors (E12-E16) | [38] | ||
| Increased expression during development (E0–P3) in C57BL/6, enriched in progenitor Hes5+ cells; use of mimics rescued Dicer1 cKO phenotype | Microarray, RT-qPCR, plasmid transfection | Prtg, Lin28b | Antiparallel expression, upregulated in Dicer1 cKO, luciferase assays, OE maintains early progenitor competence | [39] | ||
| let-7c | Increased expression in adult MG (FACS-purified from Rlbp1-CreER: tdTomatoflSTOP/flSTOP mice) compared to P2 RPC (FACS-purified from Sox2-CreER: tdTomatoflSTOP/flSTOP mice) | nCounter | Antagonistic role to miR-25 and miR-124 in MG differentiation | Ascl1 (and other mature neuronal markers?) | Increase in the number of Ascl1:tdTomato+ cells, increase in Ascl-reporter activity, defective differentiation after miR-25 mimic/let7a antagomiR (scRNA-seq) | [40] |
| let-7f | Increased expression during development (E10–P3) in C57BL/6, enriched in progenitors Hes5+ cells; use of mimics rescued Dicer1 cKO phenotype | Microarray, RT-qPCR | Prtg, Lin28b | Antiparallel expression, upregulated in Dicer1 cKO, luciferase assays, maintenance of early progenitor competence in target OE assays | [39] | |
| miR-7a | Expression pattern of increase (E14–P0), decrease (P0–P2), and increase (P2–adult) in ICR mice. Suppressed RPC differentiation into MG without perturbing proliferation | RT-qPCR, transfection | Negative MG differentiation | Notch3 | Luciferase and IHC assays after miR-7 or Notch3 manipulation levels | [41] |
| miR-9 | Preferentially expressed in retina compared to brain and heart. Increased expression during development (peak at P10) in SVJ129 mice | Microarray, RT-qPCR | [42] | |||
| Decreased expression during development (P4–adult) in C57BL/6J mice | RT-qPCR | ACCN2, ETS1, KLF13, LIN28B, SH2B3 | Pool of miR-124, miR-125, and miR-9 in HEK293 cells | [43] | ||
| Decreased expression in Dicer1 cKO (αPax6cre; R26EYFP; Dicerfl/fl) | RT-qPCR | Dicer-dependent miRNA, part of the transition from early to late progenitors (E12-E16) | [38] | |||
| Increased expression during development (E10–P3) in C57BL/6, enriched in progenitors Hes5+ cells; use of mimics rescued Dicer1 cKO phenotype | Microarray, RT-qPCR, transfection | Prtg, Lin28b | Antiparallel expression, upregulated in Dicer1 cKO, luciferase assays, OE maintains early progenitor competence | [39] | ||
| Decreased expression from DIV8 to DIV14 in cultured MG | nCounter | [44] | ||||
| OE of miR-9/9* and miR-124 suppresses RPC differentiation in glial cells and promotes their differentiation into neurons (P3, P14) in Slc:ICR mice | Plasmid electroporation | Regulation of cell fate in RPC | [45] | |||
| Decreased expression of miR-9-5p during development (P0–P21) in C57BL/6J mice | Microarray, RT-qPCR | [46] | ||||
| Preferentially expressed in retina compared to brain and heart. Increased expression of miR-9-3p during development (E10–adult) in SVJ129 mice | Microarray, RT-qPCR | [42] | ||||
| Increased expression of miR-9-3p during development (P0–P21) in C57BL/6 mice | Microarray, RT-qPCR | [46] | ||||
| miR-18a-5p | Decreased expression during development (P1–8w) in C57Bl/6 mice, suppression of endothelial function of HRMEC using agomiR | Small RNA-seq, RT-qPCR | Negative regulator of angiogenesis | FGF1, HIF1A | Luciferase reporter and Western blotting assays after manipulation of miR-18a-5p levels | [47] |
| miR-21 | Increased expression in human-cultured RPE (1 to 4 weeks) and mouse RPE/choroid/sclera explants (4 to 22 months) | RNA-seq of exosomes, RT-qPCR, ISH | Activation of p53 pathway in retinal microglia | Cdkn1a, Cdc25a, Daxx | RT-qPCR in transfected cells with miR-21 mimics | [48] |
| miR-24 | Decreased expression in Dicer1 cKO (αPax6cre; R26EYFP; Dicerfl/fl) | RT-qPCR | Dicer-dependent miRNA, part of the transition from early to late progenitors (E12-E16) | [38] | ||
| miR-25 | Preferentially expressed in retina compared to brain and heart. Increased expression during development (peak at P10) in SVJ129 mice | Microarray, RT-qPCR | [42] | |||
| Decreased expression during development (P4–adult) in C57BL/6J mice | [43] | |||||
| Decreased expression in adult MG (FACS-purified from Rlbp1-CreER: tdTomatoflSTOP/flSTOP mice) compared to P2 RPC (FACS-purified from Sox2-CreER: tdTomatoflSTOP/flSTOP mice) | nCounter | Neurogenic role: adult MG reprogramming to neuronal/RPC phenotype | Ascl1 (and other mature neuronal markers?) | Increase in the number of Ascl1:tdTomato+ cells, increase in Ascl-reporter activity, defective differentiation after miR-25 mimic/let7a antagomiR (scRNA-seq) | [40] | |
| miR-29a | Increased expression in Nrl KO mice | Microarray, RT-qPCR | [49] | |||
| Increased expression during in vitro RPC differentiation | RT-qPCR | Inhibition of RPC proliferation and induction of differentiation | RBM8A | Decreased mRNA and protein expression in RPC differentiation, luciferase assay | [50] | |
| Decreased expression of miR- 29a-3p during development (P0–P21) in C57BL/6 mice | Microarray, RT-qPCR | Inhibition of retinal angiogenesis. Potential antiangiogenic factor in oxygen-induced retinopathy | SGK3 | Protein and mRNA change following manipulation of miR-96a-3p levels | [51] | |
| miR-96 | Preferentially expressed in the retina compared to brain and heart. Increased expression during development (E10–adult) in SVJ129 mice | Microarray, RT-qPCR | Regulation of circadian-dependent expression | Adcy6 | Antiparallel circadian expression and decrease luciferase activity | [42] |
| 64 bp deletion of miR-96-5p/3p sequences results in developmental delay of photoreceptor cells | Characterization of KO mice: IHC, ERG | Indispensable for the maturation of photoreceptors, especially cones | Several (potentially > 400 genes) | Differential expression mutant vs. wt (P120) in RNA-seq | [52] | |
| miR-124 | Increased expression during development (P4–adult) in INL and ONL of C57BL/6 mice | RT-qPCR, ISH | ACCN2, ETS1, KLF13, LIN28B, SH2B3 | Pool of miR-124, miR-125, and miR-9 in HEK293 cells | [43] | |
| Decreased expression in Dicer1 cKO (αPax6cre; R26EYFP; Dicerfl/fl) | [38] | |||||
| Increased expression of miR-124-3p during development (P0–P21) in C57BL/6J mice | Microarray, RT-qPCR | [46] | ||||
| OE of miR-9/9* and miR-124 suppresses RPC differentiation in glial cells and promotes their differentiation into neurons (P3, P14) in Slc:ICR mice | Plasmid electroporation | Regulation of cell fate in RPC | [45] | |||
| KO mice for the precursor of miR-124a, Rncr3/LINC00599, shows abnormal brain including retinal cone cell death | RT-qPCR | Survival of cone photoreceptors | LHX2 | Luciferase and IHC assays | [53] | |
| miR-125a | Decreased expression during development (P4–adult) in C57BL/6J mice, enriched at ONL and INL | RT-qPCR, ISH | ACCN2, ETS1, KLF13, LIN28B, SH2B3 | Pool of miR-124, miR-125, and miR-9 in HEK293 cells | [43] | |
| Decreased expression of miR-125a-5p during development (P0–P21) in C57BL/6 mice | Microarray, RT-qPCR | [46] | ||||
| miR-125b | Increased expression during development (P4–adult) in C57BL/6J mice, enriched at ONL and INL | RT-qPCR, ISH | ACCN2, ETS1, KLF13, LIN28B, SH2B3 | Pool of miR-124, miR-125, and miR-9 in HEK293 cells | [43] | |
| Increased expression during development (E0–P3) in C57BL/6; use of mimics rescued Dicer1 cKO phenotype | Microarray, RT-qPCR | Prtg, Lin28b | Antiparallel expression, upregulated in Dicer1 cKO, luciferase assays, maintenance of early progenitor competence in target OE assays | [39] | ||
| Prominent increased expression of miR-125-5p from P11 to adult MG (FACS-purified from Rlbp1-CreER: tdTomatoflSTOP/flSTOP mice) | nCounter | [44] | ||||
| miR-132 | Uniformly expressed in the mouse RGC layer between P0 and P12 and regulated by BDNF in CD-1 mice | IHS | Axonal branching and terminal maturation of RGC | p250GAP (Arhgap32) | Axonal branching effects in OE assays | [54] |
| miR-143 | Decreased expression in Nrl KO mice | Microarray, RT-qPCR | [49] | |||
| Increased expression at E16, decreased at P1, and increased at adult C57BL/6 mice | RT-qPCR | Photoreceptor differentiation by Nrl inhibition | Nrl, Neurod1 | Luciferase and FACS (GFP) assays and expression analysis after manipulation of miR-143 levels | [55] | |
| miR-145 | Increased expression at E16, decreased at P1, and increased at adult C57BL/6 mice | RT-qPCR | Photoreceptor differentiation by Nrl inhibition | Nrl, Crx | Luciferase and FACS (GFP) analysis and expression analysis after manipulation of miR-145 levels | [55] |
| Increased expression during development (P7–P17) in C57BL/6 mice | RT-qPCR | Pathological vascularization (OIR model) but little impact on normal capillary development | Tmod3 | Luciferase assay and decreased levels in EC of OIR model | [56] | |
| miR-155 | Manipulation of miR155 levels disrupts physiological angiogenesis (P0–P8) in Lifeact-EGFP wt | Characterization of in vitro matrigel-based assays of microvascular EC | Regulation of neonatal retinal vasculature | SMAD1, SMAD5 | Protein expression and phosphorylation change upon manipulation of miR-155 levels | [57] |
| miR-182 | Preferentially expressed in retina compared to brain and heart. Increased expression during development (E10–adult) in SVJ129 mice | Microarray, RT-qPCR | Regulation of circadian-dependent expression | Adcy6 | Antiparallel circadian expression and decrease luciferase activity | [42] |
| Increased expression during development (E13.5–adult) in wt. KO mice had subtle morphological changes, reduced light response, and retinal transcriptional dysregulation | RT-qPCR, characterization of KO mice: OCT, ERG | Several (>100 genes, including Rho, Prph2, Pde6b, Opn1mw, Opn1sw, Gnat2) | Differential expression mutant vs. wt (P7, P42) in RNA-seq and RT-qPCR | [58] | ||
| miR-183 | Preferentially expressed in adult retina compared to brain and heart. Increased expression during development (E10–adult) in SVJ129 mice | Microarray, RT-qPCR | [42] | |||
| Decreased expression in Dicer1 cKO (αPax6cre; R26EYFP; Dicerfl/fl) | RT-qPCR | Dicer-dependent miRNA, part of the transition from early to late progenitors (E12-E16) | [38] | |||
| Increased expression during development (E10–P3) in C57BL/6J mice | RT-qPCR | [39] | ||||
| miR-183 KO leads to altered retinal light responses | ERG | Rnf217 | Decreased expression in retina development and after AAV-miR-183 injection; altered ERG after AAV-Rnf217 injection | [59] | ||
| miR-204 | Increased expression during development (E15–P12 and adult) in C57BL6 mice; decreased expression in Nrl KO adult mice | Microarray, RT-qPCR | [49] | |||
| Increased expression during development (P4–adult) in C57BL/6J mice | RT-qPCR | [43] | ||||
| Predominant increase from P11 to adult MG (FACS-purified from Rlbp1-CreER:tdTomatoflSTOP/flSTOP mice) and in cultured MG | nCounter | [44] | ||||
| miR-211 | Preferentially expressed in retina compared to brain and heart. Increased expression during development (E10–adult) in SVJ129 mice | Microarray, RT-qPCR | [42] | |||
| Decreased expression in Nrl KO adult mice | Microarray, RT-qPCR | [49] | ||||
| Increased expression during development (E15–P12 and adult) in C57BL6 mice | RT-qPCR | [49] | ||||
| miR-211 KO mice lead to a progressive cone dystrophy phenotype with specific cone receptor cell affectation | Immunostaining of cone markers, ERG | Regulation of cone photoreceptor survival and function | Several (>60 genes including Plin, Fbp4, Cidec, and Pck1) | Differential expression mutant vs. wt (P7, P42) in RNA-seq | [60] | |
| miR-216a/b-5p | Most downregulated miRNA in PTF1A-defective mice. Decreased expression during development (P0–P12) in CD-1 mice | Small RNA-seq, RT-qPCR, ISH | Amacrine cell formation | Foxn3 | PAR-CLIP for miR-216a/b sites and luciferase assays after manipulation of miR-216b levels | [61] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-Calvo, J.; Esquiva, G.; Gómez-Vicente, V.; Valor, L.M. MicroRNAs in the Mouse Developing Retina. Int. J. Mol. Sci. 2023, 24, 2992. https://doi.org/10.3390/ijms24032992
Navarro-Calvo J, Esquiva G, Gómez-Vicente V, Valor LM. MicroRNAs in the Mouse Developing Retina. International Journal of Molecular Sciences. 2023; 24(3):2992. https://doi.org/10.3390/ijms24032992
Chicago/Turabian StyleNavarro-Calvo, Jorge, Gema Esquiva, Violeta Gómez-Vicente, and Luis M. Valor. 2023. "MicroRNAs in the Mouse Developing Retina" International Journal of Molecular Sciences 24, no. 3: 2992. https://doi.org/10.3390/ijms24032992
APA StyleNavarro-Calvo, J., Esquiva, G., Gómez-Vicente, V., & Valor, L. M. (2023). MicroRNAs in the Mouse Developing Retina. International Journal of Molecular Sciences, 24(3), 2992. https://doi.org/10.3390/ijms24032992