
Blood–Brain Barrier Integrity Damage in Bacterial Meningitis: The Underlying Link, Mechanisms, and Therapeutic Targets

 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
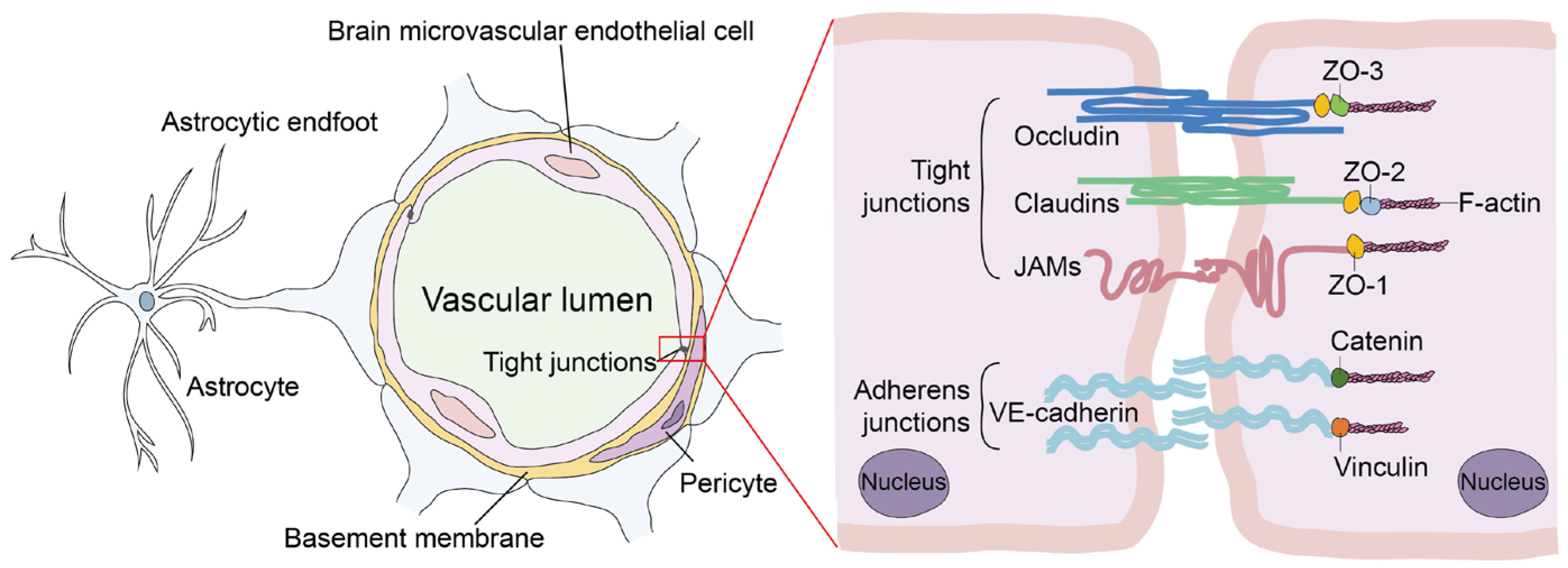
1. Introduction
2. Enhancement of BBB Permeability by Bacterial Virulence Factors
2.1. Bacterial Surface Structure
2.2. Hemolysins
2.3. Enzymes
3. Host Signaling Mediators That Regulate BBB Permeability
3.1. Cytokines
3.2. Vascularization Factors
3.3. Apoptosis Related Factors
3.4. Transcription Factors
3.5. Metalloproteinases
3.6. Non-Coding RNA
3.7. Pattern-Recognition Receptors
3.8. Others
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coureuil, M.; Lecuyer, H.; Bourdoulous, S.; Nassif, X. A journey into the brain: Insight into how bacterial pathogens cross blood-brain barriers. Nat. Rev. Microbiol. 2017, 15, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Reid, S.; Thompson, H.; Thakur, K.T. Nervous system infections and the global traveler. Semin. Neurol. 2018, 38, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S. Investigating bacterial penetration of the blood-brain barrier for the pathogenesis, prevention, and therapy of bacterial meningitis. ACS. Infect. Dis. 2020, 6, 34–42. [Google Scholar] [CrossRef]
- Lucas, M.J.; Brouwer, M.C.; van de Beek, D. Neurological sequelae of bacterial meningitis. J. Infect. 2016, 73, 18–27. [Google Scholar] [CrossRef]
- van Sorge, N.M.; Doran, K.S. Defense at the border: The blood-brain barrier versus bacterial foreigners. Future Microbiol. 2012, 7, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, R.J.; Rohlwink, U.; Misra, U.K.; van Crevel, R.; Mai, N.T.H.; Dooley, K.E.; Caws, M.; Figaji, A.; Savic, R.; Solomons, R.; et al. Tuberculous meningitis. Nat. Rev. Neurol. 2017, 13, 581–598. [Google Scholar] [CrossRef]
- Segura, M.; Fittipaldi, N.; Calzas, C.; Gottschalk, M. Critical streptococcus suis virulence factors: Are they all really critical? Trends Microbiol. 2017, 25, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Le Guennec, L.; Coureuil, M.; Nassif, X.; Bourdoulous, S. Strategies used by bacterial pathogens to cross the blood-brain barrier. Cell. Microbiol. 2020, 22, e13132. [Google Scholar] [CrossRef]
- Saunders, N.R.; Dziegielewska, K.M.; Mollgard, K.; Habgood, M.D. Physiology and molecular biology of barrier mechanisms in the fetal and neonatal brain. J. Physiol. 2018, 596, 5723–5756. [Google Scholar] [CrossRef]
- Kim, K.S. Acute bacterial meningitis in infants and children. Lancet Infect. Dis. 2010, 10, 32–42. [Google Scholar] [CrossRef]
- Bayir, E.; Sendemir, A. Role of intermediate filaments in blood-brain barrier in health and disease. Cells 2021, 10, 1400. [Google Scholar] [CrossRef] [PubMed]
- Anil, A.; Banerjee, A. Pneumococcal encounter with the blood-brain barrier endothelium. Front. Cell. Infect. Microbiol. 2020, 10, 590682. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.J.; Patabendige, A.A.K.; Dolman, D.E.M.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Cain, M.D.; Salimi, H.; Diamond, M.S.; Klein, R.S. Mechanisms of pathogen invasion into the central nervous system. Neuron 2019, 103, 771–783. [Google Scholar] [CrossRef]
- Yang, R.C.; Huang, F.; Fu, J.Y.; Dou, B.B.; Xu, B.J.; Miao, L.; Liu, W.T.; Yang, X.P.; Tan, C.; Chen, H.C.; et al. Differential transcription profiles of long non-coding RNAs in primary human brain microvascular endothelial cells in response to meningitic Escherichia coli. Sci. Rep. 2016, 6, 38903. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.C.; Huang, K.; Zhang, H.P.; Li, L.; Tan, C.; Chen, H.C.; Jin, M.L.; Wang, X.R. Transcriptional landscape of human neuroblastoma cells in response to SARS-CoV-2. BMC. Neurosci. 2022, 23, 43. [Google Scholar] [CrossRef]
- Yang, R.C.; Xu, B.J.; Yang, B.; Fu, J.Y.; Chen, H.C.; Wang, X.R. Non-coding RNAs: The extensive and interactive regulators of the blood-brain barrier permeability. RNA. Biol. 2021, 18, 108–116. [Google Scholar] [CrossRef]
- Stamatovic, S.M.; Johnson, A.M.; Keep, R.F.; Andjelkovic, A.V. Junctional proteins of the blood-brain barrier: New insights into function and dysfunction. Tissue Barriers 2016, 4, e1154641. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Campbell, M. Tight junction modulation at the blood-brain barrier: Current and future perspectives. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183298. [Google Scholar] [CrossRef]
- Yang, R.C.; Huang, K.; Zhang, H.P.; Li, L.; Zhang, Y.F.; Tan, C.; Chen, H.C.; Jin, M.L.; Wang, X.R. SARS-CoV-2 productively infects human brain microvascular endothelial cells. J. Neuroinflamm. 2022, 19, 149. [Google Scholar] [CrossRef]
- Castro Dias, M.; Coisne, C.; Lazarevic, I.; Baden, P.; Hata, M.; Iwamoto, N.; Francisco, D.M.F.; Vanlandewijck, M.; He, L.; Baier, F.A.; et al. Claudin-3-deficient C57BL/6J mice display intact brain barriers. Sci. Rep. 2019, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, D.V.; Tonry, J.H.; Kim, K.S.; Ramarao, N.; Popova, T.G.; Bailey, C.; Popov, S.; Chung, M.C. Bacillus anthracis protease InhA increases blood-brain barrier permeability and contributes to cerebral hemorrhages. PloS ONE 2011, 6, e17921. [Google Scholar] [CrossRef] [PubMed]
- Siddharthan, V.; Kim, Y.V.; Liu, S.; Kim, K.S. Human astrocytes/astrocyte-conditioned medium and shear stress enhance the barrier properties of human brain microvascular endothelial cells. Brain. Res. 2007, 1147, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S. Mechanisms of microbial traversal of the blood-brain barrier. Nat. Rev. Microbiol. 2008, 6, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S. Human meningitis-associated Escherichia coli. EcoSal Plus 2016, 7. [Google Scholar] [CrossRef]
- Doran, K.S.; Banerjee, A.; Disson, O.; Lecuit, M. Concepts and mechanisms: Crossing host barrier. Cold Spring Harb. Perspect. Med. 2013, 3, a010090. [Google Scholar] [CrossRef]
- Knirel, Y.A.; Anisimov, A.P.; Kislichkina, A.A.; Kondakova, A.N.; Bystrova, O.V.; Vagaiskaya, A.S.; Shatalin, K.Y.; Shashkov, A.S.; Dentovskaya, S.V. Lipopolysaccharide of the Yersinia pseudotuberculosis complex. Biomolecules 2021, 11, 1410. [Google Scholar] [CrossRef]
- Boitsova, E.B.; Morgun, A.V.; Osipova, E.D.; Pozhilenkova, E.A.; Martinova, G.P.; Frolova, O.V.; Olovannikova, R.Y.; Tohidpour, A.; Gorina, Y.V.; Panina, Y.A.; et al. The inhibitory effect of LPS on the expression of GPR81 lactate receptor in blood-brain barrier model in vitro. J. Neuroinflamm. 2018, 15, 196. [Google Scholar] [CrossRef]
- He, F.; Yin, F.; Omran, A.; Yang, L.F.; Xiang, Q.L.; Peng, J. PKC and RhoA signals cross-talk in Escherichia coli endotoxin induced alterations in brain endothelial permeability. Biochem. Biophys. Res. Commun. 2012, 425, 182–188. [Google Scholar] [CrossRef]
- Chang, S.; Cao, Y. Sulfasalazine maintains blood-brain barrier integrity and relieves lipopolysaccharide-induced inflammation in hCMEC/D3 cells. Neuroreport 2021, 32, 672–677. [Google Scholar] [CrossRef]
- Wispelwey, B.; Lesse, A.J.; Hansen, E.J.; Scheld, W.M. Haemophilus influenzae lipopolysaccharide-induced blood brain barrier permeability during experimental meningitis in the rat. J. Clin. Investig. 1988, 82, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Nieh, M.P.; Chen, W.; Lei, Y. Outer membrane vesicles (OMVs) enabled bio-applications: A critical review. Biotechnol. Bioeng. 2021, 119, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Wispelwey, B.; Hansen, E.J.; Scheld, W.M. Haemophilus influenzae outer membrane vesicle-induced blood-brain barrier permeability during experimental meningitis. Infect. Immun. 1989, 57, 2559–2562. [Google Scholar] [CrossRef] [PubMed]
- Lesse, A.J.; Moxon, E.R.; Zwahlen, A.; Scheld, W.M. Role of cerebrospinal fluid pleocytosis and Haemophilus influenzae type b capsule on blood brain barrier permeability during experimental meningitis in the rat. J. Clin. Investig. 1988, 82, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Roord, J.J.; Apicella, M.; Scheld, W.M. The induction of meningeal inflammation and blood-brain barrier permeability by Haemophilus influenzae type b peptidoglycan. J. Infect. Dis. 1994, 170, 254–256. [Google Scholar] [CrossRef] [PubMed]
- Popescu, C.P.; Zaharia, M.; Nica, M.; Stanciu, D.; Moroti, R.; Benea, S.; Melinte, V.; Vasile, T.; Ceausu, E.; Ruta, S.; et al. Anthrax meningoencephalitis complicated with brain abscess—A case report. Int. J. Infect. Dis. 2021, 108, 217–219. [Google Scholar] [CrossRef]
- Ebrahimi, C.M.; Kern, J.W.; Sheen, T.R.; Ebrahimi-Fardooee, M.A.; van Sorge, N.M.; Schneewind, O.; Doran, K.S. Penetration of the blood-brain barrier by Bacillus anthracis requires the pXO1-encoded BslA protein. J. Bacteriol. 2009, 191, 7165–7173. [Google Scholar] [CrossRef]
- Pian, Y.; Wang, P.; Liu, P.; Zheng, Y.; Zhu, L.; Wang, H.; Xu, B.; Yuan, Y.; Jiang, Y. Proteomics identification of novel fibrinogen-binding proteins of Streptococcus suis contributing to antiphagocytosis. Front. Cell. Infect. Microbiol. 2015, 5, 19. [Google Scholar] [CrossRef]
- Wang, J.; Kong, D.; Zhang, S.; Jiang, H.; Zheng, Y.; Zang, Y.; Hao, H.; Jiang, Y. Interaction of fibrinogen and muramidase-released protein promotes the development of Streptococcus suis meningitis. Front. Microbiol. 2015, 6, 1001. [Google Scholar] [CrossRef]
- Porubsky, S.; Federico, G.; Muthing, J.; Jennemann, R.; Gretz, N.; Buttner, S.; Obermuller, N.; Jung, O.; Hauser, I.A.; Grone, E.; et al. Direct acute tubular damage contributes to Shigatoxin-mediated kidney failure. J. Pathol. 2014, 234, 120–133. [Google Scholar] [CrossRef]
- Kong, D.C.; Chen, Z.; Wang, J.P.; Lv, Q.Y.; Jiang, H.; Zheng, Y.L.; Xu, M.K.; Zhou, X.Y.; Hao, H.J.; Jiang, Y.Q. Interaction of factor H-binding protein of Streptococcus suis with globotriaosylceramide promotes the development of meningitis. Virulence 2017, 8, 1290–1302. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Q.; Xia, J.; Tan, C.; Zhou, Y.; Wang, Y.; Zheng, C.K.; Chen, H.C.; Bei, W.C. Evaluation of the immunogenicity and the protective efficacy of a novel identified immunogenic protein, SsPepO, of Streptococcus suis serotype 2. Vaccine 2011, 29, 6514–6519. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, J.Q.; Yan, K.; Li, H.; Sun, C.F.; Zhang, S.; Yuan, F.Y.; Wang, X.R.; Tan, C.; Chen, H.C.; et al. Binding of fibronectin to SsPepO facilitates the development of Streptococcus suis meningitis. J. Infect. Dis. 2018, 217, 973–982. [Google Scholar] [CrossRef]
- Esgleas, M.; Li, Y.; Hancock, M.A.; Harel, J.; Dubreuil, J.D.; Gottschalk, M. Isolation and characterization of alpha-enolase, a novel fibronectin-binding protein from Streptococcus suis. Microbiology 2008, 154 Pt 9, 2668–2679. [Google Scholar] [CrossRef]
- Lopez-Alemany, R.; Longstaff, C.; Hawley, S.; Mirshahi, M.; Fabregas, P.; Jardi, M.; Merton, E.; Miles, L.A.; Felez, J. Inhibition of cell surface mediated plasminogen activation by a monoclonal antibody against alpha-Enolase. Am. J. Hematol. 2003, 72, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, N.; Zhang, J.; Liu, H.; Liu, J.; Xia, X.; Sun, C.; Feng, X.; Gu, J.; Du, C.; et al. Enolase of Streptococcus Suis serotype 2 enhances blood-brain barrier permeability by inducing IL-8 release. Inflammation 2016, 39, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Shang, X.; Chen, Y.; Zheng, Y.; Huang, W.; Jiang, H.; Lv, Q.; Kong, D.; Jiang, Y.; Liu, P. Bacteria elevate extracellular adenosine to exploit host signaling for blood-brain barrier disruption. Virulence 2020, 11, 980–994. [Google Scholar] [CrossRef]
- Ma, Z.; Peng, J.; Zhang, H.; Xu, B.; Pei, X.M.; Lin, H.X.; Lu, C.P.; Fan, H.J. SILAC and LC-MS/MS identification of Streptococcus equi ssp zooepidemicus proteins that contribute to mouse brain microvascular endothelial cell infection. Appl. Microbiol. Biotechnol. 2016, 100, 7125–7136. [Google Scholar]
- Ma, Z.; Peng, J.; Yu, D.D.; Park, J.S.; Lin, H.X.; Xu, B.; Lu, C.P.; Fan, H.J.; Waldor, M.K. A streptococcal Fic domain-containing protein disrupts blood-brain barrier integrity by activating moesin in endothelial cells. PLoS Pathog. 2019, 15, e1007737. [Google Scholar] [CrossRef]
- Price, K.E.; Greene, N.G.; Camilli, A. Export requirements of pneumolysin in Streptococcus pneumoniae. J. Bacteriol. 2012, 194, 3651–3660. [Google Scholar] [CrossRef]
- van Pee, K.; Mulvihill, E.; Muller, D.J.; Yildiz, O. Unraveling the pore-forming steps of pneumolysin from Streptococcus pneumoniae. Nano Lett. 2016, 16, 7915–7924. [Google Scholar] [CrossRef]
- Gentschev, I.; Dietrich, G.; Goebel, W. The E. coli alpha-hemolysin secretion system and its use in vaccine development. Trends Microbiol. 2002, 10, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.Y.; Li, L.; Huo, D.; Yang, R.C.; Yang, B.; Xu, B.J.; Yang, X.P.; Dai, M.H.; Tan, C.; Chen, H.C.; et al. Meningitic Escherichia coli alpha-hemolysin aggravates blood-brain barrier disruption via targeting TGFβ1-triggered hedgehog signaling. Mol. Brain 2021, 14, 116. [Google Scholar] [CrossRef] [PubMed]
- Vanier, G.; Segura, M.; Friedl, P.; Lacouture, S.; Gottschalk, M. Invasion of porcine brain microvascular endothelial cells by Streptococcus suis serotype 2. Infect. Immun. 2004, 72, 1441–1449. [Google Scholar] [CrossRef]
- Vanier, G.; Segura, M.; Lecours, M.P.; Grenier, D.; Gottschalk, M. Porcine brain microvascular endothelial cell-derived interleukin-8 is first induced and then degraded by Streptococcus suis. Microb. Pathog. 2009, 46, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Chen, Y.; Lv, Q.; Zheng, Y.; Kong, D.; Jiang, H.; Huang, W.; Ren, Y.; Liu, P.; Jiang, Y. Suilyin disrupts the blood-brain barrier by activating group III secretory phospholipase A2. Life 2022, 12, 919. [Google Scholar] [CrossRef]
- Cossart, P. Illuminating the landscape of host-pathogen interactions with the bacterium Listeria monocytogenes. Proc. Natl. Acad. Sci. USA 2011, 108, 19484–19491. [Google Scholar] [CrossRef]
- Zhang, T.; Bae, D.; Wang, C. Listeriolysin O mediates cytotoxicity against human brain microvascular endothelial cells. FEMS Microbiol. Lett. 2015, 362, fnv084. [Google Scholar] [CrossRef]
- Yau, B.; Hunt, N.H.; Mitchell, A.J.; Too, L.K. Blood-brain barrier pathology and CNS outcomes in Streptococcus pneumoniae meningitis. Int. J. Mol. Sci. 2018, 19, 3555. [Google Scholar] [CrossRef] [PubMed]
- Zysk, G.; Schneider-Wald, B.K.; Hwang, J.H.; Bejo, L.; Kim, K.S.; Mitchell, T.J.; Hakenbeck, R.; Heinz, H.P. Pneumolysin is the main inducer of cytotoxicity to brain microvascular endothelial cells caused by Streptococcus pneumoniae. Infect. Immun. 2001, 69, 845–852. [Google Scholar] [CrossRef]
- Chen, J.Q.; Li, N.N.; Wang, B.W.; Liu, X.F.; Liu, J.L.; Chang, Q. Upregulation of CBP by PLY can cause permeability of blood-brain barrier to increase meningitis. J. Biochem. Mol. Toxicol. 2019, 33, e22333. [Google Scholar] [CrossRef]
- Hupp, S.; Heimeroth, V.; Wippel, C.; Fortsch, C.; Ma, J.T.; Mitchell, T.J.; Iliev, A.I. Astrocytic tissue remodeling by the meningitis neurotoxin pneumolysin facilitates pathogen tissue penetration and produces interstitial brain edema. Glia 2012, 60, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Lembo, A.; Gurney, M.A.; Burnside, K.; Banerjee, A.; de los Reyes, M.; Connelly, J.E.; Lin, W.J.; Jewell, K.A.; Vo, A.; Renken, C.W.; et al. Regulation of CovR expression in Group B Streptococcus impacts blood-brain barrier penetration. Mol. Microbiol. 2010, 77, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Vornhagen, J.; Quach, P.; Boldenow, E.; Merillat, S.; Whidbey, C.; Ngo, L.Y.; Adams Waldorf, K.M.; Rajagopal, L. Bacterial hyaluronidase promotes ascending GBS infection and preterm birth. mBio 2016, 7, e00781-16. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Cao, Q.; Ma, K.; Wang, Z.; Liu, G.; Lu, C.; Liu, Y. Quantitative assessment of the blood-brain barrier opening caused by Streptococcus agalactiae hyaluronidase in a BALB/c mouse model. Sci. Rep. 2017, 7, 13529. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Cao, Q.; Luo, S.; Wang, Z.; Liu, G.; Lu, C.; Liu, Y. Cas9 enhances bacterial virulence by repressing the regR transcriptional regulator in Streptococcus agalactiae. Infect. Immun. 2018, 86, e00552-17. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.F.; Goss, L.; Dworkin, J. Eukaryote-like serine/threonine kinases and phosphatases in bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 192–212. [Google Scholar] [CrossRef] [PubMed]
- Rui, L.; Weiyi, L.; Yu, M.; Hong, Z.; Jiao, Y.; Zhe, M.; Hongjie, F. The serine/threonine protein kinase of Streptococcus suis serotype 2 affects the ability of the pathogen to penetrate the blood-brain barrier. Cell. Microbiol. 2018, 20, e12862. [Google Scholar]
- Barichello, T.; Pereira, J.S.; Savi, G.D.; Generoso, J.S.; Cipriano, A.L.; Silvestre, C.; Petronilho, F.; Dal-Pizzol, F.; Vilela, M.C.; Teixeira, A.L. A kinetic study of the cytokine/chemokines levels and disruption of blood-brain barrier in infant rats after pneumococcal meningitis. J. Neuroimmunol. 2011, 233, 12–17. [Google Scholar] [CrossRef]
- Azkur, A.K.; Akdis, M.; Azkur, D.; Sokolowska, M.; van de Veen, W.; Bruggen, M.C.; O’Mahony, L.; Gao, Y.D.; Nadeau, K.; Akdis, C.A. Immune response to SARS-CoV-2 and mechanisms of immunopathological changes in COVID-19. Allergy 2020, 75, 1564–1581. [Google Scholar] [CrossRef]
- de Vries, H.E.; Blom-Roosemalen, M.C.; van Oosten, M.; de Boer, A.G.; van Berkel, T.J.; Breimer, D.D.; Kuiper, J. The influence of cytokines on the integrity of the blood-brain barrier in vitro. J. Neuroimmunol. 1996, 64, 37–43. [Google Scholar] [CrossRef]
- Tanabe, S.; Gottschalk, M.; Grenier, D. Hemoglobin and Streptococcus suis cell wall act in synergy to potentiate the inflammatory response of monocyte-derived macrophages. Innate Immun. 2008, 14, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Barichello, T.; Lemos, J.C.; Generoso, J.S.; Cipriano, A.L.; Milioli, G.L.; Marcelino, D.M.; Vuolo, F.; Petronilho, F.; Dal-Pizzol, F.; Vilela, M.C.; et al. Oxidative stress, cytokine/chemokine and disruption of blood-brain barrier in neonate rats after meningitis by Streptococcus agalactiae. Neurochem. Res. 2011, 36, 1922–1930. [Google Scholar] [CrossRef] [PubMed]
- Yau, B.; Mitchell, A.J.; Too, L.K.; Ball, H.J.; Hunt, N.H. Interferon-γ-induced nitric oxide synthase-2 contributes to blood/brain barrier dysfunction and acute mortality in experimental Streptococcus pneumoniae meningitis. J. Interferon Cytokine Res. 2016, 36, 86–99. [Google Scholar] [CrossRef]
- Gilpin, T.E.; Walter, F.R.; Herbath, M.; Wigand, K.; Sandor, M.; Fabry, Z. Mycobacterium bovis bacillus calmette-guérin-infected dendritic cells induce TNF-α-dependent cell cluster formation that damage brain endothelial cells in an in vitro model of the blood brain barrier. J. Immunol. 2020, 204, 1065–1077. [Google Scholar] [CrossRef]
- Tsao, N.; Hsu, H.P.; Wu, C.M.; Liu, C.C.; Lei, H.Y. Tumour necrosis factor-alpha causes an increase in blood-brain barrier permeability during sepsis. J. Med. Microbiol. 2001, 50, 812–821. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.T.; Lv, Y.J.; Yang, R.C.; Fu, J.Y.; Liu, L.; Wang, H.; Cao, Q.; Tan, C.; Chen, H.C.; Wang, X.R. New insights into meningitic Escherichia coli infection of brain microvascular endothelial cells from quantitative proteomics analysis. J. Neuroinflamm. 2018, 15, 291. [Google Scholar] [CrossRef] [PubMed]
- Ireton, K.; Mortuza, R.; Gyanwali, G.C.; Gianfelice, A.; Hussain, M. Role of internalin proteins in the pathogenesis of Listeria monocytogenes. Mol. Microbiol. 2021, 116, 1407–1419. [Google Scholar] [CrossRef]
- Shahid, A.D.; Lu, Y.; Iqbal, M.A.; Lin, L.; Huang, S.; Jiang, X.G.; Chen, S.X. Listeria monocytogenes crosses blood brain barrier through Rho GTPases induced migration of macrophages and inflammatory interleukin expression. Microb. Pathog. 2021, 159, 105143. [Google Scholar] [CrossRef]
- Ma, Y.; Zechariah, A.; Qu, Y.; Hermann, D.M. Effects of vascular endothelial growth factor in ischemic stroke. J. Neurosci. Res. 2012, 90, 1873–1882. [Google Scholar] [CrossRef]
- Dzietko, M.; Derugin, N.; Wendland, M.F.; Vexler, Z.S.; Ferriero, D.M. Delayed VEGF treatment enhances angiogenesis and recovery after neonatal focal rodent stroke. Transl. Stroke Res. 2013, 4, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.C.; Liu, W.T.; Miao, L.; Yang, X.P.; Fu, J.Y.; Dou, B.B.; Cai, A.L.; Zong, X.; Tan, C.; Chen, H.C.; et al. Induction of VEGFA and Snail-1 by meningitic Escherichia coli mediates disruption of the blood-brain barrier. Oncotarget 2016, 7, 63839–63855. [Google Scholar] [CrossRef]
- Yang, R.C.; Lv, Y.J.; Miao, L.; Zhang, H.P.; Qu, X.Y.; Chen, J.Q.; Xu, B.J.; Yang, B.; Fu, J.Y.; Tan, C.; et al. Resveratrol attenuates meningitic Escherichia coli-mediated blood-brain barrier disruption. ACS. Infect. Dis. 2021, 7, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.N.; Pan, R.; Qin, X.J.; Yang, W.L.; Qi, Z.F.; Liu, W.L.; Liu, K.J. Ischemic neurons activate astrocytes to disrupt endothelial barrier via increasing VEGF expression. J. Neurochem. 2014, 129, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Caporarello, N.; Olivieri, M.; Cristaldi, M.; Scalia, M.; Toscano, M.A.; Genovese, C.; Addamo, A.; Salmeri, M.; Lupo, G.; Anfuso, C.D. Blood-brain barrier in a Haemophilus influenzae type a in vitro infection: Role of adenosine receptors A2A and A2B. Mol. Neurobiol. 2018, 55, 5321–5336. [Google Scholar] [CrossRef]
- Zucchi, F.C.R.; Tsanaclis, A.M.C.; Moura-Dias, Q.; Silva, C.L.; Pelegrini-da-Silva, A.; Neder, L.; Takayanagui, O.M. Modulation of angiogenic factor VEGF by DNA-hsp65 vaccination in a murine CNS tuberculosis model. Tuberculosis 2013, 93, 373–380. [Google Scholar] [CrossRef]
- van der Flier, M.; Hoppenreijs, S.; van Rensburg, A.J.; Nurs, D.; Ruyken, M.; Kolk, A.H.J.; Springer, P.; Hoepelman, A.I.M.; Geelen, S.P.M.; Kimpen, J.L.L.; et al. Vascular endothelial growth factor and blood-brain barrier disruption in tuberculous meningitis. Pediatr. Infect. Dis. J. 2004, 23, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, R.H.; Kantarjian, H.M.; Cortes, J.E. Biology of platelet-derived growth factor and its involvement in disease. Mayo Clin. Proc. 2006, 81, 1241–1257. [Google Scholar] [CrossRef]
- Kastin, A.J.; Akerstrom, V.; Hackler, L.; Pan, W.H. Different mechanisms influencing permeation of PDGF-AA and PDGF-BB across the blood-brain barrier. J. Neurochem. 2003, 87, 7–12. [Google Scholar] [CrossRef]
- Yao, H.H.; Duan, M.; Buch, S. Cocaine-mediated induction of platelet-derived growth factor: Implication for increased vascular permeability. Blood 2011, 117, 2538–2547. [Google Scholar] [CrossRef]
- Yang, R.C.; Qu, X.Y.; Xiao, S.Y.; Li, L.; Xu, B.J.; Fu, J.Y.; Lv, Y.J.; Amjad, N.; Tan, C.; Kim, K.S.; et al. Meningitic Escherichia coli-induced upregulation of PDGF-B and ICAM-1 aggravates blood-brain barrier disruption and neuroinflammatory response. J. Neuroinflamm. 2019, 16, 101. [Google Scholar] [CrossRef]
- Fernandez-Hernando, C.; Suarez, Y. ANGPTL4: A multifunctional protein involved in metabolism and vascular homeostasis. Curr. Opin. Hematol. 2020, 27, 206–213. [Google Scholar] [CrossRef]
- Garcia-Roman, J.; Zentella-Dehesa, A. Vascular permeability changes involved in tumor metastasis. Cancer Lett. 2013, 335, 259–269. [Google Scholar] [CrossRef]
- Liu, L.; Li, J.X.; Huo, D.; Peng, Z.; Yang, R.C.; Fu, J.Y.; Xu, B.J.; Yang, B.; Chen, H.C.; Wang, X.R. Meningitic Escherichia coli induction of ANGPTL4 in brain microvascular endothelial cells contributes to blood-brain barrier disruption via ARHGAP5/RhoA/MYL5 signaling cascade. Pathogens 2019, 8, 254. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.T.; Leaver, H.A. Brain endothelial cell death: Modes, signaling pathways, and relevance to neural development, homeostasis, and disease. Mol. Neurobiol. 2010, 42, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Basuroy, S.; Bhattacharya, S.; Leffler, C.W.; Parfenova, H. Nox4 NADPH oxidase mediates oxidative stress and apoptosis caused by TNF-alpha in cerebral vascular endothelial cells. Am. J. Physiol. Cell Physiol. 2009, 296, C422–C432. [Google Scholar] [CrossRef]
- Bermpohl, D.; Halle, A.; Freyer, D.; Dagand, E.; Braun, J.S.; Bechmann, I.; Schroder, N.W.J.; Weber, J.R. Bacterial programmed cell death of cerebral endothelial cells involves dual death pathways. J. Clin. Investig. 2005, 115, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, B.; Vanier, G.; Jacques, M.; Gottschalk, M. Interactions of Haemophilus parasuis and its LOS with porcine brain microvascular endothelial cells. Vet. Res. 2008, 39, 42. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lei, S.; Jia, L.; Xia, X.; Sun, Y.; Jiang, H.; Zhu, R.; Li, S.; Qu, G.; Gu, J.; et al. Streptococcus suis serotype 2 enolase interaction with host brain microvascular endothelial cells and RPSA-induced apoptosis lead to loss of BBB integrity. Vet. Res. 2021, 52, 30. [Google Scholar] [CrossRef] [PubMed]
- Jurenas, D.; Journet, L. Activity, delivery, and diversity of Type VI secretion effectors. Mol. Microbiol. 2021, 115, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Wang, X.; Shou, J.; Zong, B.; Zhang, Y.; Tan, J.; Chen, J.; Hu, L.; Zhu, Y.; Chen, H.; et al. Roles of Hcp family proteins in the pathogenesis of the porcine extraintestinal pathogenic Escherichia coli type VI secretion system. Sci. Rep. 2016, 6, 26816. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Tao, J.; Yu, H.; Ni, J.; Zeng, L.; Teng, Q.; Kim, K.S.; Zhao, G.P.; Guo, X.; Yao, Y. Hcp family proteins secreted via the type VI secretion system coordinately regulate Escherichia coli K1 interaction with human brain microvascular endothelial cells. Infect. Immun. 2012, 80, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Bruick, R.K.; McKnight, S.L. A conserved family of prolyl-4-hydroxylases that modify HIF. Science 2001, 294, 1337–1340. [Google Scholar] [CrossRef] [PubMed]
- Agani, F.H.; Pichiule, P.; Chavez, J.C.; LaManna, J.C. The role of mitochondria in the regulation of hypoxia-inducible factor 1 expression during hypoxia. J. Biol. Chem. 2000, 275, 35863–35867. [Google Scholar] [CrossRef] [PubMed]
- Devraj, G.; Guerit, S.; Seele, J.; Spitzer, D.; Macas, J.; Khel, M.I.; Heidemann, R.; Braczynskiz, A.K.; Ballhorn, W.; Gunther, S.; et al. HIF-1 alpha is involved in blood-brain barrier dysfunction and paracellular migration of bacteria in pneumococcal meningitis. Acta Neuropathol. 2020, 140, 183–208. [Google Scholar] [CrossRef] [PubMed]
- Arzumanyan, A.; Friedman, T.; Kotei, E.; Ng, I.O.L.; Lian, Z.; Feitelson, M.A. Epigenetic repression of E-cadherin expression by hepatitis B virus x antigen in liver cancer. Oncogene 2012, 31, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, E.L.; Liu, C.J.; Fearon, E.R.; Margolis, B. The transcription factor snail represses Crumbs3 expression and disrupts apico-basal polarity complexes. Oncogene 2008, 27, 3875–3879. [Google Scholar] [CrossRef]
- Carrozzino, F.; Soulie, P.; Huber, D.; Mensi, N.; Orci, L.; Cano, A.; Feraille, E.; Montesano, R. Inducible expression of Snail selectively increases paracellular ion permeability and differentially modulates tight junction proteins. Am. J. Physiol. Cell Physiol. 2005, 289, C1002–C1014. [Google Scholar] [CrossRef] [PubMed]
- Ohkubo, T.; Ozawa, M. The transcription factor Snail downregulates the tight junction components independently of E-cadherin downregulation. J. Cell Sci. 2004, 117, 1675–1685. [Google Scholar] [CrossRef]
- Kim, B.J.; Hancock, B.M.; Bermudez, A.; Del Cid, N.; Reyes, E.; van Sorge, N.M.; Lauth, X.; Smurthwaite, C.A.; Hilton, B.J.; Stotland, A.; et al. Bacterial induction of Snail1 contributes to blood-brain barrier disruption. J. Clin. Investig. 2015, 125, 2473–2483. [Google Scholar] [CrossRef]
- Rohlwink, U.K.; Walker, N.F.; Ordonez, A.A.; Li, Y.J.; Tucker, E.W.; Elkington, P.T.; Wilkinson, R.J.; Wilkinson, K.A. Matrix Metalloproteinases in Pulmonary and Central Nervous System Tuberculosis-A Review. Int. J. Mol. Sci. 2019, 20, 1350. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Hawkins, K.E.; Dore, S.; Candelario-Jalil, E. Neuroinflammatory mechanisms of blood-brain barrier damage in ischemic stroke. Am. J. Physiol. Cell Physiol. 2019, 316, C135–C153. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Estrada, E.Y.; Thompson, J.F.; Liu, W.L.; Rosenberg, G.A. Matrix metalloproteinase-mediated disruption of tight junction proteins in cerebral vessels is reversed by synthetic matrix metalloproteinase inhibitor in focal ischemia in rat. J. Cereb. Blood Flow Metab. 2007, 27, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Meli, D.N.; Christen, S.; Leib, S.L. Matrix metalloproteinase-9 in pneumococcal meningitis: Activation via an oxidative pathway. J. Infect. Dis. 2003, 187, 1411–1415. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.W.; Pabisiak, P.J.; Brilha, S.; Singh, P.; Roncaroli, F.; Elkington, P.T.; Friedland, J.S. Complex regulation of neutrophil-derived MMP-9 secretion in central nervous system tuberculosis. J. Neuroinflamm. 2017, 14, 31. [Google Scholar] [CrossRef]
- Mailankody, S.; Dangeti, G.V.; Soundravally, R.; Joseph, N.M.; Mandal, J.; Dutta, T.K.; Kadhiravan, T. Cerebrospinal fluid matrix metalloproteinase 9 levels, blood-brain barrier permeability, and treatment outcome in tuberculous meningitis. PLoS ONE 2017, 12, e0181262. [Google Scholar] [CrossRef]
- Brilha, S.; Ong, C.W.M.; Weksler, B.; Romero, N.; Couraud, P.O.; Friedland, J.S. Matrix metalloproteinase-9 activity and a downregulated Hedgehog pathway impair blood-brain barrier function in an in vitro model of CNS tuberculosis. Sci. Rep. 2017, 7, 16031. [Google Scholar] [CrossRef]
- Ricci, S.; Grandgirard, D.; Wenzel, M.; Braccini, T.; Salvatore, P.; Oggioni, M.R.; Leib, S.L.; Koedel, U. Inhibition of matrix metalloproteinases attenuates brain damage in experimental meningococcal meningitis. BMC. Infect. Dis. 2014, 14, 726. [Google Scholar] [CrossRef]
- Schubert-Unkmeir, A.; Konrad, C.; Slanina, H.; Czapek, F.; Hebling, S.; Frosch, M. Neisseria meningitidis induces brain microvascular endothelial cell detachment from the matrix and cleavage of occludin: A role for MMP-8. PLoS Pathog. 2010, 6, e1000874. [Google Scholar] [CrossRef]
- Tenenbaum, T.; Matalon, D.; Adam, R.; Seibt, A.; Wewer, C.; Schwerk, C.; Galla, H.J.; Schroten, H. Dexamethasone prevents alteration of tight junction-associated proteins and barrier function in porcine choroid plexus epithelial cells after infection with Streptococcus suis in vitro. Brain Res. 2008, 1229, 1–17. [Google Scholar] [CrossRef]
- Levi, M.; Scully, M.; Singer, M. The role of ADAMTS-13 in the coagulopathy of sepsis. J. Thromb. Haemost. 2018, 16, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Karayigit, M.O.; Dincel, G.C. Role of ADAMTS-13 and nNOS expression in neuropathogenesis of listeric encephalitis of small ruminants. Biotech. Histochem. 2020, 95, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.W.; Liu, Z.; Shi, Z.S. Non-coding RNA in acute ischemic stroke: Mechanisms, biomarkers and therapeutic targets. Cell Transplant. 2018, 27, 1763–1777. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Xu, B.; Yang, B.; Fu, J.; Liu, L.; Amjad, N.; Cai, A.; Tan, C.; Chen, H.; Wang, X. Circular RNA transcriptomic analysis of primary human brain microvascular endothelial cells infected with meningitic Escherichia coli. Mol. Ther. Nucleic Acids 2018, 13, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Chen, J.; Xu, B.; Yang, B.; Fu, J.; Xiao, S.; Tan, C.; Chen, H.; Wang, X. Circ_2858 helps blood-brain barrier disruption by increasing VEGFA via sponging miR-93-5p during Escherichia coli meningitis. Mol. Ther. Nucleic Acids 2020, 22, 708–721. [Google Scholar] [CrossRef]
- Xu, B.; Yang, R.; Fu, J.; Yang, B.; Chen, J.; Tan, C.; Chen, H.; Wang, X. LncRSPH9-4 facilitates meningitic Escherichia coli-caused blood-brain barrier disruption via miR-17-5p/MMP3 axis. Int. J. Mol. Sci. 2021, 22, 6364. [Google Scholar] [CrossRef]
- Zheng, K.; He, F.L.B.; Liu, H.S.; He, Q.S. Genetic variations of toll-like receptors: Impact on susceptibility, severity and prognosis of bacterial meningitis. Infect. Genet. Evol. 2021, 93, 104984. [Google Scholar] [CrossRef]
- Brightbill, H.D.; Libraty, D.H.; Krutzik, S.R.; Yang, R.B.; Belisle, J.T.; Bleharski, J.R.; Maitland, M.; Norgard, M.V.; Plevy, S.E.; Smale, S.T.; et al. Host defense mechanisms triggered by microbial lipoproteins through toll-like receptors. Science 1999, 285, 732–736. [Google Scholar] [CrossRef]
- Echchannaoui, H.; Frei, K.; Schnell, C.; Leib, S.L.; Zimmerli, W.; Landmann, R. Toll-like receptor 2-deficient mice are highly susceptible to Streptococcus pneumoniae meningitis because of reduced bacterial clearing and enhanced inflammation. J. Infect. Dis. 2002, 186, 798–806. [Google Scholar] [CrossRef]
- Kim, H.J. Role of nucleotide-binding and oligomerization domain 2 protein (NOD2) in the development of atherosclerosis. Korean J. Physiol. Pharmacol. 2015, 19, 479–484. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.J.; Liu, Q. NOD2 expression in Streptococcus pneumoniae meningitis and its influence on the blood-brain barrier. Can. J. Infect. Dis. Med. Microbiol. 2018, 2018, 7292084. [Google Scholar] [CrossRef] [PubMed]
- Dickson, K.B.; Zhou, J. Role of reactive oxygen species and iron in host defense against infection. Front. Biosci. 2020, 25, 1600–1616. [Google Scholar]
- Silwal, P.; Kim, J.K.; Kim, Y.J.; Jo, E.K. Mitochondrial reactive oxygen species: Double-edged weapon in host defense and pathological inflammation during infection. Front. Immunol. 2020, 11, 1649. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, A.; Rochfort, K.D.; McDonnell, C.J.; Kerrigan, S.W.; Cummins, P.M. Staphylococcus aureus-mediated blood-brain barrier injury: An in vitro human brain microvascular endothelial cell model. Cell. Microbiol. 2017, 19, e12664. [Google Scholar] [CrossRef] [PubMed]
- Aratani, Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Miric, D.; Katanic, R.; Kisic, B.; Zoric, L.; Miric, B.; Mitic, R.; Dragojevic, I. Oxidative stress and myeloperoxidase activity during bacterial meningitis: Effects of febrile episodes and the BBB permeability. Clin. Biochem. 2010, 43, 246–252. [Google Scholar] [CrossRef]
- Nisole, S.; Stoye, J.P.; Saib, A. TRIM family proteins: Retroviral restriction and antiviral defence. Nat. Rev. Microbiol. 2005, 3, 799–808. [Google Scholar] [CrossRef]
- Albor, A.; El-Hizawi, S.; Horn, E.J.; Laederich, M.; Frosk, P.; Wrogemann, K.; Kulesz-Martin, M. The interaction of Piasy with Trim32, an E3-ubiquitin ligase mutated in limb-girdle muscular dystrophy type 2H, promotes Piasy degradation and regulates UVB-induced keratinocyte apoptosis through NFkappaB. J. Biol. Chem. 2006, 281, 25850–25866. [Google Scholar] [CrossRef]
- OuYang, X.; Guo, J.; Lv, Q.; Jiang, H.; Zheng, Y.; Liu, P.; Zhao, T.; Kong, D.; Hao, H.; Jiang, Y. TRIM32 drives pathogenesis in Streptococcal toxic shock-like syndrome and Streptococcus suis meningitis by regulating innate immune responses. Infect. Immun. 2020, 88, e00957-19. [Google Scholar] [CrossRef]
- Chaudhuri, A.; Martinez-Martin, P.; Kennedy, P.G.; Andrew Seaton, R.; Portegies, P.; Bojar, M.; Steiner, I.; Force, E.T. EFNS guideline on the management of community-acquired bacterial meningitis: Report of an EFNS Task Force on acute bacterial meningitis in older children and adults. Eur. J. Neurol. 2008, 15, 649–659. [Google Scholar] [CrossRef]
- Xie, Y.; Kim, K.J.; Kim, K.S. Current concepts on Escherichia coli K1 translocation of the blood-brain barrier. FEMS. Immunol. Med. Microbiol. 2004, 42, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Woehrl, B.; Klein, M.; Grandgirard, D.; Koedel, U.; Leib, S. Bacterial meningitis: Current therapy and possible future treatment options. Expert Rev. Anti-Infect. Ther. 2011, 9, 1053–1065. [Google Scholar] [CrossRef] [PubMed]
- Segarra, M.; Aburto, M.R.; Acker-Palmer, A. Blood-brain barrier dynamics to maintain brain homeostasis. Trends Neurosci. 2021, 44, 393–405. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, R.; Wang, J.; Wang, F.; Zhang, H.; Tan, C.; Chen, H.; Wang, X. Blood–Brain Barrier Integrity Damage in Bacterial Meningitis: The Underlying Link, Mechanisms, and Therapeutic Targets. Int. J. Mol. Sci. 2023, 24, 2852. https://doi.org/10.3390/ijms24032852
Yang R, Wang J, Wang F, Zhang H, Tan C, Chen H, Wang X. Blood–Brain Barrier Integrity Damage in Bacterial Meningitis: The Underlying Link, Mechanisms, and Therapeutic Targets. International Journal of Molecular Sciences. 2023; 24(3):2852. https://doi.org/10.3390/ijms24032852
Chicago/Turabian StyleYang, Ruicheng, Jundan Wang, Fen Wang, Huipeng Zhang, Chen Tan, Huanchun Chen, and Xiangru Wang. 2023. "Blood–Brain Barrier Integrity Damage in Bacterial Meningitis: The Underlying Link, Mechanisms, and Therapeutic Targets" International Journal of Molecular Sciences 24, no. 3: 2852. https://doi.org/10.3390/ijms24032852
APA StyleYang, R., Wang, J., Wang, F., Zhang, H., Tan, C., Chen, H., & Wang, X. (2023). Blood–Brain Barrier Integrity Damage in Bacterial Meningitis: The Underlying Link, Mechanisms, and Therapeutic Targets. International Journal of Molecular Sciences, 24(3), 2852. https://doi.org/10.3390/ijms24032852