
A Genome-Wide Association Study Identified Novel Genetic Susceptibility Loci for Oral Cancer in Taiwan

 , , , and
, , , and
Abstract

1. Introduction
2. Results
2.1. Demographics of Study Population
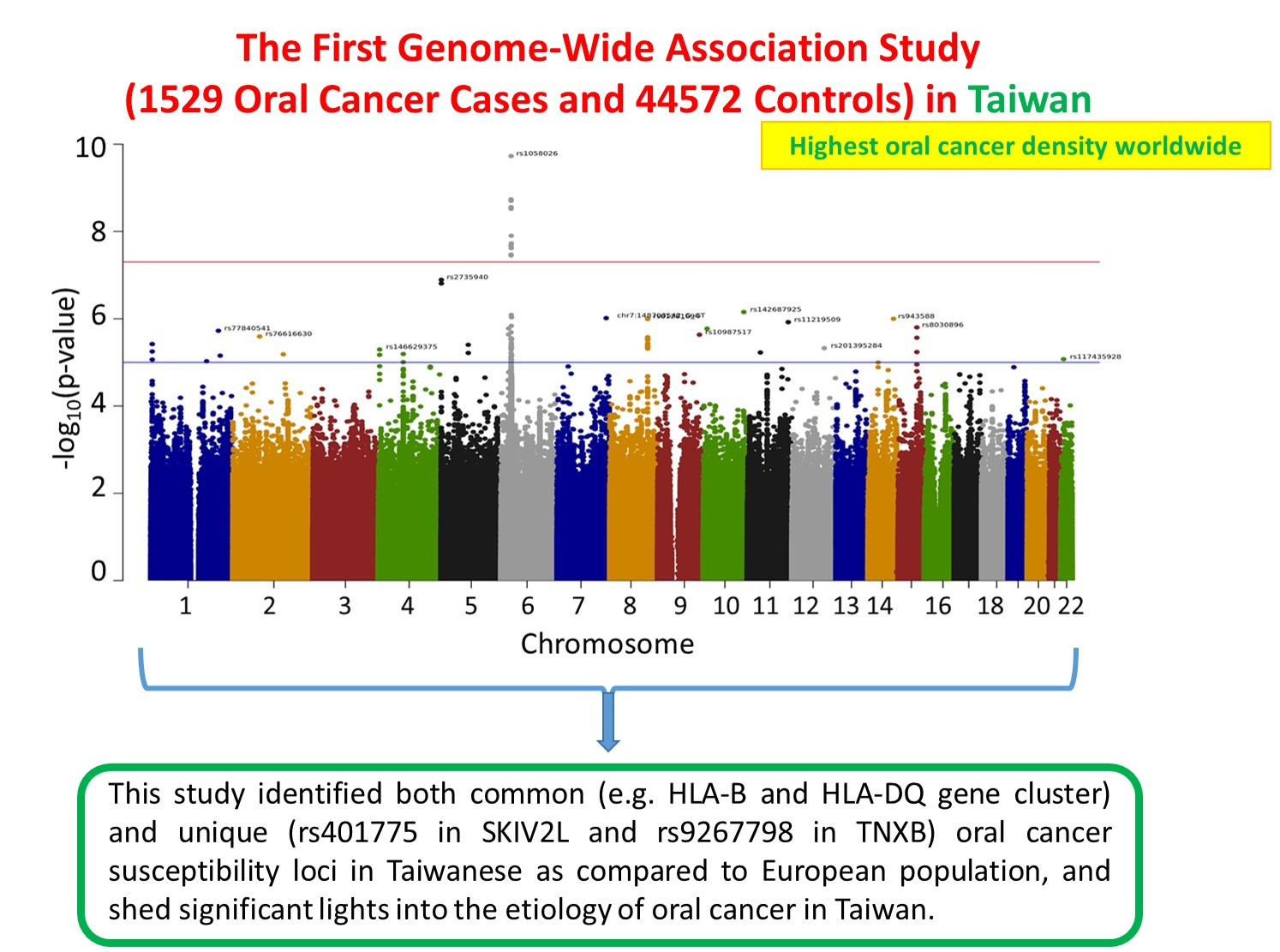
2.2. Novel Variants Associated with Oral Cancer in the Taiwanese
2.3. Common Variants Associated with Oral Cancer Validated among Various Populations
3. Discussion
4. Materials and Methods
4.1. Study Population and Data Collection
4.2. Genotyping and Imputation
4.3. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Jhuang, J.-R.; Su, S.-Y.; Chiang, C.-J.; Yang, Y.-W.; Lin, L.-J.; Hsu, T.-H.; Lee, W.-C. Forecast of peak attainment and imminent decline after 2017 of oral cancer incidence in men in Taiwan. Sci. Rep. 2022, 12, 5726. [Google Scholar] [CrossRef]
- Chuang, S.-L.; Su, W.W.-Y.; Chen, S.L.-S.; Yen, A.M.-F.; Wang, C.-P.; Fann, J.C.-Y.; Chiu, S.Y.-H.; Lee, Y.-C.; Chiu, H.-M.; Chang, D.-C.; et al. Population-based screening program for reducing oral cancer mortality in 2,334,299 Taiwanese cigarette smokers and/or betel quid chewers. Cancer 2017, 123, 1597–1609. [Google Scholar] [CrossRef]
- Huang, Y.-T.; Ho, P.-S.; Jou, Y.-Y.; Wu, C.-Y.; Wang, Y.-W.; Yang, Y.-H. Determining High Prevalence of Betel-Quid Chewing and Cigarette Smoking by Occupation Using the Taiwan National Health Interview Survey. Subst. Use Misuse 2020, 55, 1472–1482. [Google Scholar] [CrossRef]
- Wang, T.-H.; Hsia, S.-M.; Shih, Y.-H.; Shieh, T.-M. Association of Smoking, Alcohol Use, and Betel Quid Chewing with Epigenetic Aberrations in Cancers. Int. J. Mol. Sci. 2017, 18, 1210. [Google Scholar] [CrossRef]
- Petti, S.; Masood, M.; Scully, C. The Magnitude of Tobacco Smoking-Betel Quid Chewing-Alcohol Drinking Interaction Effect on Oral Cancer in South-East Asia. A Meta-Analysis of Observational Studies. PLoS ONE 2013, 8, e78999. [Google Scholar] [CrossRef]
- Lee, Y.A.; Li, S.; Chen, Y.; Li, Q.; Chen, C.; Hsu, W.; Lou, P.; Zhu, C.; Pan, J.; Shen, H.; et al. Tobacco smoking, alcohol drinking, betel quid chewing, and the risk of head and neck cancer in an East Asian population. Head Neck 2019, 41, 92–102. [Google Scholar] [CrossRef]
- Lin, W.-J.; Jiang, R.-S.; Wu, S.-H.; Chen, F.-J.; Liu, S.-A. Smoking, Alcohol, and Betel Quid and Oral Cancer: A Prospective Cohort Study. J. Oncol. 2011, 2011, 525976. [Google Scholar] [CrossRef]
- Toporcov, T.N.; Znaor, A.; Zhang, Z.-F.; Yu, G.-P.; Winn, D.M.; Wei, Q.; Vilensky, M.; Vaughan, T.; Thomson, P.; Talamini, R.; et al. Risk factors for head and neck cancer in young adults: A pooled analysis in the INHANCE consortium. Int. J. Epidemiol. 2015, 44, 169–185. [Google Scholar] [CrossRef]
- Garavello, W.; Foschi, R.; Talamini, R.; La Vecchia, C.; Rossi, M.; Maso, L.D.; Tavani, A.; Levi, F.; Barzan, L.; Ramazzotti, V.; et al. Family history and the risk of oral and pharyngeal cancer. Int. J. Cancer 2008, 122, 1827–1831. [Google Scholar] [CrossRef]
- Radoï, L.; Paget-Bailly, S.; Guida, F.; Cyr, D.; Menvielle, G.; Schmaus, A.; Carton, M.; Cenée, S.; Sanchez, M.; Guizard, A.-V.; et al. Family history of cancer, personal history of medical conditions and risk of oral cavity cancer in France: The ICARE study. BMC Cancer 2013, 13, 560. [Google Scholar] [CrossRef] [PubMed]
- Bau, D.-T.; Tsai, M.-H.; Huang, C.-Y.; Lee, C.-C.; Tseng, H.-C.; Lo, Y.-L.; Tsai, Y.; Tsai, F.-J. Relationship between polymorphisms of nucleotide excision repair genes and oral cancer risk in Taiwan: Evidence for modification of smoking habit. Chin. J. Physiol. 2007, 50, 294–300. [Google Scholar] [PubMed]
- Bau, D.T.; Tsai, M.H.; Lo, Y.L.; Hsu, C.M.; Tsai, Y.; Lee, C.C.; Tsai, F.J. Association of p53 and p21(CDKN1A/WAF1/CIP1) polymorphisms with oral cancer in Taiwan patients. Anticancer Res. 2007, 27, 1559–1564. [Google Scholar]
- Tsai, M.-H.; Tseng, H.-C.; Liu, C.-S.; Chang, C.-L.; Tsai, C.-W.; Tsou, Y.-A.; Wang, R.-F.; Lin, C.-C.; Wang, H.-C.; Chiu, C.-F.; et al. Interaction of Exo1 genotypes and smoking habit in oral cancer in Taiwan. Oral Oncol. 2009, 45, e90–e94. [Google Scholar] [CrossRef]
- Bau, D.-T.; Chang, C.-H.; Tsai, M.-H.; Chiu, C.-F.; Tsou, Y.-A.; Wang, R.-F.; Tsai, C.-W.; Tsai, R.-Y. Association between DNA repair gene ATM polymorphisms and oral cancer susceptibility. Laryngoscope 2010, 120, 2417–2422. [Google Scholar] [CrossRef]
- Tsai, M.-H.; Tsai, C.-W.; Tsou, Y.-A.; Hua, C.-H.; Hsu, C.-F.; Bau, D.-T. Significant association of cyclin D1 single nucleotide polymorphisms with oral cancer in taiwan. Anticancer Res. 2011, 31, 227–231. [Google Scholar]
- Tsai, C.-W.; Chang, W.-S.; Lin, K.-C.; Shih, L.-C.; Tsai, M.-H.; Hsiao, C.-L.; Yang, M.-D.; Lin, C.-C.; Bau, D.-T. Significant association of Interleukin-10 genotypes and oral cancer susceptibility in Taiwan. Anticancer Res. 2014, 34, 3731–3737. [Google Scholar]
- Tsai, C.-W.; Chang, W.-S.; Liu, J.-C.; Tsai, M.-H.; Lin, C.-C.; Bau, D.-T. Contribution of DNA double-strand break repair gene XRCC3 genotypes to oral cancer susceptibility in Taiwan. Anticancer Res. 2014, 34, 2951–2956. [Google Scholar]
- Hung, Y.-W.; Tsai, C.-W.; Wu, C.-N.; Shih, L.-C.; Chen, Y.-Y.; Liu, Y.-F.; Hung, H.-S.; Shen, M.-Y.; Chang, W.-S.; Bau, D.-T. The Contribution of Matrix Metalloproteinase-8 Promoter Polymorphism to Oral Cancer Susceptibility. Vivo 2017, 31, 585–590. [Google Scholar] [CrossRef]
- Hashibe, M.; McKay, J.D.; Curado, M.P.; Oliveira, J.C.; Koifman, S.; Koifman, R.; Zaridze, D.; Shangina, O.; Wünsch-Filho, V.; Eluf-Neto, J.; et al. Multiple ADH genes are associated with upper aerodigestive cancers. Nat. Genet. 2008, 40, 707–709. [Google Scholar] [CrossRef]
- Fan, J.; Liu, W.; Zhang, M.; Xing, C. A literature review and systematic meta-analysis on XRCC3 Thr241Met polymorphism associating with susceptibility of oral cancer. Oncol. Lett. 2019, 18, 3265–3273. [Google Scholar] [CrossRef]
- Cadoni, G.; Boccia, S.; Petrelli, L.; Di Giannantonio, P.; Arzani, D.; Giorgio, A.; De Feo, E.; Pandolfini, M.; Gallì, P.; Paludetti, G.; et al. A review of genetic epidemiology of head and neck cancer related to polymorphisms in metabolic genes, cell cycle control and alcohol metabolism. Acta Otorhinolaryngol. Ital. 2012, 32, 1–11. [Google Scholar]
- McKay, J.D.; Truong, T.; Gaborieau, V.; Chabrier, A.; Chuang, S.C.; Byrnes, G.; Zaridze, D.; Shangina, O.; Szeszenia-Dabrowska, N.; Lissowska, J.; et al. A genome-wide association study of upper aerodigestive tract cancers conducted within the INHANCE consortium. PLoS Genet. 2011, 7, e1001333. [Google Scholar] [CrossRef]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef] [PubMed]
- Visscher, P.M.; Wray, N.R.; Zhang, Q.; Sklar, P.; McCarthy, M.I.; Brown, M.A.; Yang, J. 10 Years of GWAS Discovery: Biology, Function, and Translation. Am. J. Hum. Genet. 2017, 101, 5–22. [Google Scholar] [CrossRef]
- Park, S.L.; Cheng, I.; Haiman, C.A. Genome-Wide Association Studies of Cancer in Diverse Populations. Cancer Epidemiol. Biomark. Prev. 2018, 27, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Lesseur, C.; Diergaarde, B.; Olshan, A.F.; Wünsch-Filho, V.; Ness, A.R.; Liu, G.; Lacko, M.; Eluf-Neto, J.; Franceschi, S.; Lagiou, P.; et al. Genome-wide association analyses identify new susceptibility loci for oral cavity and pharyngeal cancer. Nat. Genet. 2016, 48, 1544–1550. [Google Scholar] [CrossRef]
- Shete, S.; Liu, H.; Wang, J.; Yu, R.; Sturgis, E.M.; Li, G.; Dahlstrom, K.R.; Liu, Z.; Amos, C.I.; Wei, Q. A Genome-Wide Association Study Identifies Two Novel Susceptible Regions for Squamous Cell Carcinoma of the Head and Neck. Cancer Res 2020, 80, 2451–2460. [Google Scholar] [CrossRef]
- Mosaad, Y.M. Clinical Role of Human Leukocyte Antigen in Health and Disease. Scand. J. Immunol. 2015, 82, 283–306. [Google Scholar] [CrossRef]
- Traherne, J.A. Human MHC architecture and evolution: Implications for disease association studies. Int. J. Immunogenet. 2008, 35, 179–192. [Google Scholar] [CrossRef]
- Robinson, J.; Soormally, A.R.; Hayhurst, J.D.; Marsh, S.G. The IPD-IMGT/HLA Database—New developments in reporting HLA variation. Hum. Immunol. 2016, 77, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Broderick, P.; Webb, E.; Wu, X.; Vijayakrishnan, J.; Matakidou, A.; Qureshi, M.; Dong, Q.; Gu, X.; Chen, W.V.; et al. Common 5p15.33 and 6p21.33 variants influence lung cancer risk. Nat. Genet. 2008, 40, 1407–1409. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.-K.; Sun, J.; Cao, G.; Liu, Y.; Lin, D.; Gao, Y.-Z.; Ren, W.-H.; Long, X.-D.; Zhang, H.; Ma, X.-P.; et al. Genetic variants in STAT4 and HLA-DQ genes confer risk of hepatitis B virus–related hepatocellular carcinoma. Nat. Genet. 2013, 45, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Bowden, S.J.; Bodinier, B.; Kalliala, I.; Zuber, V.; Vuckovic, D.; Doulgeraki, T.; Whitaker, M.D.; Wielscher, M.; Cartwright, R.; Tsilidis, K.K.; et al. Genetic variation in cervical preinvasive and invasive disease: A genome-wide association study. Lancet Oncol. 2021, 22, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Kweon, S.-S.; Tanikawa, C.; Jia, W.-H.; Xiang, Y.-B.; Cai, Q.; Zeng, C.; Schmit, S.L.; Shin, A.; Matsuo, K.; et al. Large-Scale Genome-Wide Association Study of East Asians Identifies Loci Associated with Risk for Colorectal Cancer. Gastroenterology 2019, 156, 1455–1466. [Google Scholar] [CrossRef]
- Lin, W.-Y.; Fordham, S.E.; Hungate, E.; Sunter, N.J.; Elstob, C.; Xu, Y.; Park, C.; Quante, A.; Strauch, K.; Gieger, C.; et al. Genome-wide association study identifies susceptibility loci for acute myeloid leukemia. Nat. Commun. 2021, 12, 6233. [Google Scholar] [CrossRef]
- Lin, G.-W.; Xu, C.; Chen, K.; Huang, H.-Q.; Chen, J.; Song, B.; Chan, J.K.C.; Li, W.; Liu, W.; Shih, L.-Y.; et al. Genetic risk of extranodal natural killer T-cell lymphoma: A genome-wide association study in multiple populations. Lancet Oncol. 2016, 21, 306–316. [Google Scholar] [CrossRef]
- Sud, A.; Thomsen, H.; Law, P.J.; Försti, A.; Filho, M.I.D.S.; Holroyd, A.; Broderick, P.; Orlando, G.; Lenive, O.; Wright, L.; et al. Genome-wide association study of classical Hodgkin lymphoma identifies key regulators of disease susceptibility. Nat. Commun. 2017, 8, 1892. [Google Scholar] [CrossRef]
- Wei, Q.; Yu, D.; Liu, M.; Wang, M.; Zhao, M.; Liu, M.; Jia, W.; Ma, H.; Fang, J.; Xu, W.; et al. Genome-wide association study identifies three susceptibility loci for laryngeal squamous cell carcinoma in the Chinese population. Nat. Genet. 2014, 46, 1110–1114. [Google Scholar] [CrossRef]
- Ji, P.; Chang, J.; Wei, X.; Song, X.; Yuan, H.; Gong, L.; Li, Y.; Ding, D.; Zhang, E.; Yan, C.; et al. Genetic variants associated with expression of TCF19 contribute to the risk of head and neck cancer in Chinese population. J. Med. Genet. 2022, 59, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Rafnar, T.; Sulem, P.; Stacey, S.N.; Geller, F.; Gudmundsson, J.; Sigurdsson, A.; Jakobsdottir, M.; Helgadottir, H.; Thorlacius, S.; Aben, K.K.H.; et al. Sequence variants at the TERT-CLPTM1L locus associate with many cancer types. Nat. Genet. 2009, 41, 221–227. [Google Scholar] [CrossRef]
- Shete, S.; Hosking, F.J.; Robertson, L.B.; Dobbins, S.E.; Sanson, M.; Malmer, B.; Simon, M.; Marie, Y.; Boisselier, B.; Delattre, J.-Y.; et al. Genome-wide association study identifies five susceptibility loci for glioma. Nat. Genet. 2009, 41, 899–904. [Google Scholar] [CrossRef]
- Petersen, G.M.; Amundadottir, L.; Fuchs, C.S.; Kraft, P.; Stolzenberg-Solomon, R.Z.; Jacobs, K.B.; Arslan, A.A.; Bueno-De-Mesquita, H.B.; Gallinger, S.; Gross, M.; et al. A genome-wide association study identifies pancreatic cancer susceptibility loci on chromosomes 13q22.1, 1q32.1 and 5p15.33. Nat. Genet. 2010, 42, 224–228. [Google Scholar] [CrossRef]
- Haiman, C.; Chen, G.K.; Vachon, C.M.; Canzian, F.; Dunning, A.M.; Millikan, R.C.; Wang, X.; Ademuyiwa, F.; Ahmed, S. A common variant at the TERT-CLPTM1L locus is associated with estrogen receptor–negative breast cancer. Nat. Genet. 2011, 43, 1210–1214. [Google Scholar] [CrossRef]
- Chen, H.; Majumdar, A.; Wang, L.; Kar, S.; Brown, K.M.; Feng, H.; Turman, C.; Dennis, J.; Easton, D.; Michailidou, K.; et al. Large-scale cross-cancer fine-mapping of the 5p15.33 region reveals multiple independent signals. Hum. Genet. Genom. Adv. 2021, 2, 100041. [Google Scholar] [CrossRef]
- Tian, J.; Wang, Y.; Dong, Y.; Chang, J.; Wu, Y.; Chang, S.; Che, G. Cumulative Evidence for Relationships Between Multiple Variants in the TERT and CLPTM1L Region and Risk of Cancer and Non-Cancer Disease. Front. Oncol. 2022, 12, 946039. [Google Scholar] [CrossRef]
- Scherf, D.B.; Sarkisyan, N.; Jacobsson, H.; Claus, R.; Bermejo, J.L.; Peil, B.; Gu, L.; Muley, T.; Meister, M.; Dienemann, H.; et al. Epigenetic screen identifies genotype-specific promoter DNA methylation and oncogenic potential of CHRNB4. Oncogene 2013, 32, 3329–3338. [Google Scholar] [CrossRef]
- Helbig, S.; Wockner, L.; Bouendeu, A.; Hille-Betz, U.; McCue, K.; French, J.D.; Edwards, S.L.; Pickett, H.A.; Reddel, R.R.; Chenevix-Trench, G.; et al. Functional dissection of breast cancer risk-associated TERT promoter variants. Oncotarget 2017, 8, 67203–67217. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, N.; Miao, L.; Yuan, H.; Wang, R.; Jiang, H. Alcohol dehydrogenase-1B Arg47His polymorphism is associated with head and neck cancer risk in Asian: A meta-analysis. Tumor Biol. 2015, 36, 1023–1027. [Google Scholar] [CrossRef]
- Tian, J.; Zhu, L.; Wang, W.; Zhang, L.; Li, Z.; Zhao, Q.; Xing, K.; Feng, Z.; Peng, X. Genomic Analysis of Microbulbifer sp. Strain A4B-17 and the Characterization of Its Metabolic Pathways for 4-Hydroxybenzoic Acid Synthesis. Front. Microbiol. 2018, 9, 3115. [Google Scholar] [CrossRef]
- Witwicki, R.M.; Ekram, M.B.; Qiu, X.; Janiszewska, M.; Shu, S.; Kwon, M.; Trinh, A.; Frias, E.; Ramadan, N.; Hoffman, G.; et al. TRPS1 Is a Lineage-Specific Transcriptional Dependency in Breast Cancer. Cell Rep. 2018, 25, 1255–1267.e5. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, X.; Gong, X.; Wu, L.; Zhang, J.; Liu, W.; Li, J.; Chen, L. Atypical GATA transcription factor TRPS1 represses gene expression by recruiting CHD4/NuRD(MTA2) and suppresses cell migration and invasion by repressing TP63 expression. Oncogenesis 2018, 7, 96. [Google Scholar] [CrossRef]
- Yang, L.; Gong, X.; Wang, J.; Fan, Q.; Yuan, J.; Yang, X.; Sun, X.; Li, Y.; Wang, Y. Functional mechanisms of TRPS1 in disease progression and its potential role in personalized medicine. Pathol. Res. Pract. 2022, 237, 154022. [Google Scholar] [CrossRef]
- Yang, J.; Liu, X.; Huang, Y.; He, L.; Zhang, W.; Ren, J.; Wang, Y.; Wu, J.; Wu, X.; Shan, L.; et al. TRPS1 drives heterochromatic origin refiring and cancer genome evolution. Cell Rep. 2021, 34, 108814. [Google Scholar] [CrossRef]
- Wu, L.; Wang, Y.; Liu, Y.; Yu, S.; Xie, H.; Shi, X.; Qin, S.; Ma, F.; Tan, T.Z.; Thiery, J.P.; et al. A central role for TRPS1 in the control of cell cycle and cancer development. Oncotarget 2014, 5, 7677–7690. [Google Scholar] [CrossRef] [PubMed]
- Stinson, S.; Lackner, M.R.; Adai, A.T.; Yu, N.; Kim, H.J.; O’Brien, C.; Spoerke, J.; Jhunjhunwala, S.; Boyd, Z.; Januario, T.; et al. TRPS1 targeting by miR-221/222 promotes the epithelial-to-mesenchymal transition in breast cancer. Sci. Signal 2011, 4, ra41. [Google Scholar] [CrossRef]
- Hu, J.; Su, P.; Jia, M.; Wu, X.; Zhang, H.; Li, W.; Zhou, G. TRPS1 expression promotes angiogenesis and affects VEGFA expression in breast cancer. Exp. Biol. Med. 2014, 239, 423–429. [Google Scholar] [CrossRef]
- Liu, H.; Liao, Y.; Tang, M.; Wu, T.; Tan, D.; Zhang, S.; Wang, H. Trps1 is associated with the multidrug resistance of lung cancer cell by regulating MGMT gene expression. Cancer Med. 2018, 7, 1921–1932. [Google Scholar] [CrossRef]
- Serandour, A.A.; Mohammed, H.; Miremadi, A.; Mulder, K.W.; Carroll, J.S. TRPS1 regulates oestrogen receptor binding and histone acetylation at enhancers. Oncogene 2018, 37, 5281–5291. [Google Scholar] [CrossRef]
- Strating, J.R.; Hafmans, T.G.; Martens, G.J. Functional diversity among p24 subfamily members. Biol. Cell 2009, 101, 207–220. [Google Scholar] [CrossRef]
- Zheng, H.; Yang, Y.; Han, J.; Jiang, W.-H.; Chen, C.; Wang, M.-C.; Gao, R.; Li, S.; Tian, T.; Wang, J.; et al. TMED3 promotes hepatocellular carcinoma progression via IL-11/STAT3 signaling. Sci. Rep. 2016, 6, 37070. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Luo, Y.; Li, Q. TMED3 Promotes Proliferation and Migration in Breast Cancer Cells by Activating Wnt/beta-Catenin Signaling. Onco. Targets Ther. 2020, 13, 5819–5830. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Li, Y.; Ye, X.; Ji, Y.; Chen, Y.; Zhang, X.; Li, Z. TMED3/RPS15A Axis promotes the development and progression of osteosarcoma. Cancer Cell Int. 2021, 21, 630. [Google Scholar] [CrossRef]
- Zhang, D.; Sun, L.; Zhang, J. TMED3 exerts a protumor function in non-small cell lung cancer by enhancing the Wnt/beta-catenin pathway via regulation of AKT. Toxicol. Appl. Pharmacol. 2021, 433, 115793. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.; Zhang, M.; Wang, C. Analysis and Validation of TMED3 correlates with poor prognosis and tumor immune infiltration of glioma. J. Cancer Res. Clin. Oncol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Qi, Y. Depleting TMED3 alleviates the development of endometrial carcinoma. Cancer Cell Int. 2022, 22, 231. [Google Scholar] [CrossRef]
- Wang, R.-F.; Hong, Y.-G.; Hao, L.-Q.; Yu, H.-T. Expression of TMED3 is independently associated with colorectal cancer prognosis. Exp. Ther. Med. 2022, 23, 286. [Google Scholar] [CrossRef]
- Bhatnagar, R.; Dabholkar, J.; Saranath, D. Genome-wide disease association study in chewing tobacco associated oral cancers. Oral Oncol. 2012, 48, 831–835. [Google Scholar] [CrossRef]
- Liu, T.-Y.; Lin, C.-F.; Wu, H.-T.; Wu, Y.-L.; Chen, Y.-C.; Liao, C.-C.; Chou, Y.-P.; Chao, D.; Lu, H.-F.; Chang, Y.-S.; et al. Comparison of Multiple Imputation Algorithms and Verification Using Whole-Genome Sequencing in the CMUH Genetic Biobank. Biomedicine 2021, 11, 57–65. [Google Scholar] [CrossRef]
- Liao, W.-L.; Liu, T.-Y.; Cheng, C.-F.; Chou, Y.-P.; Wang, T.-Y.; Chang, Y.-W.; Chen, S.-Y.; Tsai, F.-J. Analysis of HLA Variants and Graves’ Disease and Its Comorbidities Using a High Resolution Imputation System to Examine Electronic Medical Health Records. Front. Endocrinol. 2022, 13, 842673. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.; Whitwham, A.; Keane, T.; McCarthy, S.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. GigaScience 2021, 10, giab008. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Cases, N = 1529 | Controls, N = 44,572 |
|---|---|---|
| Age, mean (SD) | 55 (11.5) | 54 (14.5) |
| Sex (N, %) | ||
| Male | 1328 (86.9) | 34,616 (77.7) |
| Female | 201 (13.1) | 9956 (22.3) |
| CHROM | POS | ID | Gene | REF | ALT | OR (95% CI) | p |
|---|---|---|---|---|---|---|---|
| 6 | 31263915 | rs2524113 | TCF19/HLA-C | T | G | 1.32 (1.2–1.46) | 3.40 × 10−8 |
| 6 | 31264003 | rs2524112 | TCF19/HLA-C | A | G | 1.32 (1.2–1.46) | 3.40 × 10−8 |
| 6 | 31264212 | rs2853961 | TCF19/HLA-C | G | A | 1.32 (1.2–1.46) | 2.12 × 10−8 |
| 6 | 31264470 | rs2844624 | TCF19/HLA-C | T | C | 1.32 (1.2–1.46) | 3.40 × 10−8 |
| 6 | 31264813 | rs2524107 | TCF19/HLA-C | C | T | 1.32 (1.2–1.46) | 3.40 × 10−8 |
| 6 | 31265454 | rs2246085 | TCF19/HLA-C | A | T | 1.32 (1.2–1.46) | 3.40 × 10−8 |
| 6 | 31265793 | rs2248885 | HLA-C/CLIC1 | C | T | 1.32 (1.2–1.46) | 3.54 × 10−8 |
| 6 | 31353774 | rs75456009 | HLA-C/CLIC1 | A | G | 1.32 (1.2–1.44) | 2.77 × 10−9 |
| 6 | 31353782 | rs74615740 | HLA-C/CLIC1 | C | A | 1.32 (1.21–1.45) | 1.98 × 10−9 |
| 6 | 31353786 | rs75256987 | HLA-C/CLIC1 | A | C | 1.32 (1.21–1.45) | 1.84 × 10−9 |
| 6 | 31353800 | rs114322933 | HLA-C/CLIC1 | C | T | 1.32 (1.2–1.45) | 3.01 × 10−9 |
| 6 | 31353908 | rs1058026 | HLA-B | A | C | 1.34 (1.23–1.47) | 1.87 × 10−10 |
| 6 | 31949780 | rs4151658 | CFB | C | CAT | 1.37 (1.23–1.53) | 2.41 × 10−8 |
| 6 | 31963360 | rs401775 | SKIV2L | T | C | 1.34 (1.21–1.49) | 1.25 × 10−8 |
| 6 | 32077057 | rs9267798 | TNXB | G | C | 0.76 (0.69–0.84) | 1.88 × 10−8 |
| Region | SNP | Chrom:Position | Gene | Allele | EAF Cases/Controls | OR | p | OR (95% CI) | p |
|---|---|---|---|---|---|---|---|---|---|
| Lesseur et al. Nature Genetics, 2016 | (Taiwanese) | (Taiwanese) | |||||||
| OC and OPC | |||||||||
| 4q23 | rs1229984 | 4:100239319 | ADH1B | G/A | 0.03/0.06 | 0.56 | 2.29 × 10−15 | 1.20 (1.06–1.35) | 0.00335 |
| 6p21.32 | rs3828805 | 6:32636120 | HLA-DQB1 | C/T | 0.75/0.72 | 1.28 | 3.35 × 10−13 | 0.75 (0.58–0.96) | 0.0249 |
| 10q26.13 | rs201982221 | 10:126157446 | LHPP | Ins/Del | 0.03/0.02 | 1.67 | 1.58 × 10−9 | NA | NA |
| 11p15.4 | rs1453414 | 11:5829084 | OR52N2–TRIM5 | A/C | 0.23/0.20 | 1.19 | 4.78 × 10−8 | NA | NA |
| OC | |||||||||
| 2p23.3 | rs6547741 | 2:27855924 | GPN1 | G/A | 0.50/0.54 | 0.83 | 3.97 × 10−8 | 1.00 (0.84–1.20) | 0.98 |
| 4q23 | rs1229984 | 4:100239319 | ADH1B | G/A | 0.03/0.06 | 0.57 | 1.09 × 10−9 | 1.20 (1.06–1.35) | 0.003 |
| 5p15.33 | rs10462706 | 5:1343794 | CLPTM1L | C/T | 0.12/0.15 | 0.74 | 5.54 × 10−10 | 0.93 (0.83–1.04) | 0.234 |
| 9p21.3 | rs8181047 | 9:22064465 | CDKN2B-AS1 | G/A | 0.29/0.24 | 1.24 | 3.80 × 10−9 | 1.17 (0.99–1.37) | 0.064 |
| 9q34.12 | rs928674 | 9:133952024 | LAMC3 | A/G | 0.14/0.12 | 1.33 | 2.09 × 10−8 | 0.75 (0.63–0.90) | 0.002 |
| Shete et al. Cancer Research, 2020 | |||||||||
| HNSCC | |||||||||
| 6p22.1 | rs259919 | 6:30025503 | ZNRD1-AS1 | A/G | 0.34/0.31 | 1.15 (1.06–1.25) | 2.96 × 10−9 | 0.94 (0.83–1.07) | 0.353 |
| 6p21.33 | rs1265081 | 6:31111675 | CCHCR1 | C/A | 0.47/0.5 | 0.85 (0.79–0.92) | 3.75 × 10−10 | 0.95 (0.85–1.07) | 0.414 |
| 6p21.32 | rs3135001 | 6:32670136 | HLA-DQB1 | T/C | 0.21/0.25 | 0.76 (0.69–0.83) | 1.44 × 10−16 | 0.76 (0.59–0.97) | 0.03 |
| OC | |||||||||
| 5p15.33 | rs10462706 | 5:1343794 | CLPTM1L | T/C | 0.12/0.15 | 0.72 (0.60–0.88) | 7.87 × 10−11 | 0.93 (0.83–1.05) | 0.234 |
| 6p21.32 | rs1049055 | 6:32634387 | HLA-DQB1 | C/T | 0.23/0.27 | 0.76 (0.66–0.89) | 2.96 × 10−9 | 0.71 (0.55–0.92) | 0.009 |
| Ji et al. J Med Genet, 2022 | |||||||||
| HNSCC | |||||||||
| 6p22.1 | rs2517611 | 6:30169327 | TRIM26 | A/G | 0.15/0.11 | 1.32 (1.18–1.47) | 7.45 × 10−7 | 1.22 (0.96–1.55) | 0.096 |
| 6p21.33 | rs2524182 | 6:31130593 | TRIM39-RPP21, HLA-E | A/T | 0.37/0.31 | 1.27 (1.17–1.37) | 1.76 × 10−9 | 1.04 (0.90–1.21) | 0.562 |
| 6p21.33 | rs3131018 | 6:31143582 | PSORS1C3-TCF19 | C/A | 0.22/0.30 | 0.77 (0.71–0.83) | 2.17 × 10−10 | 0.84 (0.75–0.94) | 0.004 |
| 9p21.3 | rs1063192 | 9:22003367 | CDKN2B | A/G | 0.24/0.19 | 1.33 (1.2–1.45) | 4.10 × 10−12 | 1.11 (0.96–1.28) | 0.166 |
| McKay et al. Plos Genetics, 2011 | |||||||||
| UADT Cancer | |||||||||
| 4q23 | rs1229984 | 4:99318162 | ADH1B | C/T | EAF: 0.05 | 0.64 (0.59–0.71) | 1.00 × 10−20 | 1.20 (1.06–1.35) | 0.00335 |
| 4q23 | rs971074 (or rs1573496) | 4:99420704 | ADH7 | G/C | EAF: 0.12 | 0.75 (0.70–0.80) | 9.00 × 10−19 | 1.22 (1.02–1.44) | 0.0264 |
| 4q23 | rs1789924 (or rs698) | 4:99353129 | ADH1C | T/C | EAF: 0.39 | 1.12 (1.07–1.17) | 3.00 × 10−7 | 1.13 (0.93–1.36) | 0.215 |
| 12q24 | rs4767364 | 12:112083644 | ALDH2 | G/A | EAF: 0.27 | 1.13 (1.08–1.18) | 2.00 × 10−8 | 0.89 (0.71–1.10) | 0.283 |
| 4q21 | rs1494961 | 4:83453327 | HEL308 | T/C | EAF: 0.49 | 1.12 (1.08–1.17) | 1.00 × 10−8 | 0.94 (0.83–1.06) | 0.33 |
| Region | SNP ID | Chr:Position | Gene | Effect Allele | Other Allele | Europeans | Taiwanese | ||
|---|---|---|---|---|---|---|---|---|---|
| OR | p | OR | p | ||||||
| 2p23.3 | rs4665986 | 2:27755168 | A | G | 0.84 | 1.75 × 10−7 | 1.114 | 0.030 | |
| 2p23.3 | rs4665381 | 2:27757150 | C | A | 0.84 | 1.30 × 10−7 | 1.101 | 0.054 | |
| 2p23.3 | rs1972669 | 2:27767107 | G | T | 0.84 | 1.50 × 10−7 | 1.101 | 0.054 | |
| 2p23.3 | rs4493210 | 2:27768072 | T | G | 0.84 | 1.08 × 10−7 | 1.101 | 0.053 | |
| 2p23.3 | rs1080060 | 2:27768808 | A | T | 0.84 | 1.08 × 10−7 | 1.101 | 0.054 | |
| 2p23.3 | rs2384626 | 2:27773648 | G | A | 0.84 | 2.93 × 10−7 | 1.101 | 0.054 | |
| 2p23.3 | rs1820297 | 2:27774924 | G | A | 0.84 | 6.23 × 10−8 | 1.096 | 0.065 | |
| 2p23.3 | rs6750943 | 2:27777904 | G | T | 0.84 | 9.17 × 10−8 | 1.096 | 0.065 | |
| 2p23.3 | rs6761095 | 2:27778010 | T | G | 0.84 | 1.78 × 10−7 | 1.096 | 0.065 | |
| 2p23.3 | rs6706610 | 2:27778091 | A | G | 0.84 | 6.41 × 10−8 | 1.096 | 0.065 | |
| 2p23.3 | rs6547709 | 2:27783418 | A | G | 0.84 | 8.93 × 10−8 | 1.094 | 0.068 | |
| 2p23.3 | rs12714204 | 2:27787540 | C | T | 0.84 | 1.66 × 10−7 | 1.094 | 0.068 | |
| 2p23.3 | rs4260197 | 2:27794150 | G | C | 0.84 | 1.70 × 10−7 | 1.095 | 0.068 | |
| 2p23.3 | rs10205364 | 2:27794233 | T | C | 0.84 | 1.20 × 10−7 | 1.095 | 0.068 | |
| 2p23.3 | rs10165098 | 2:27796526 | T | C | 0.84 | 8.30 × 10−8 | 1.095 | 0.068 | |
| 2p23.3 | rs1919125 | 2:27801403 | C2orf16 | G | C | 0.84 | 1.23 × 10−7 | 1.095 | 0.066 |
| 2p23.3 | rs1919126 | 2:27801418 | C2orf16 | C | A | 1.19 | 1.28 × 10−7 | 1.095 | 0.066 |
| 2p23.3 | rs13026621 | 2:27807624 | ZNF512 | A | G | 0.84 | 8.99 × 10−8 | 1.091 | 0.079 |
| 2p23.3 | rs4665995 | 2:27813827 | ZNF512 | G | T | 0.84 | 9.66 × 10−8 | 1.091 | 0.079 |
| 2p23.3 | rs4665996 | 2:27814343 | ZNF512 | C | T | 0.84 | 9.96 × 10−8 | 1.091 | 0.079 |
| 2p23.3 | rs6738089 | 2:27815278 | ZNF512 | G | A | 0.84 | 2.14 × 10−7 | 1.091 | 0.079 |
| 2p23.3 | rs4665997 | 2:27816989 | ZNF512 | C | T | 0.84 | 1.53 × 10−7 | 1.094 | 0.072 |
| 2p23.3 | rs7604798 | 2:27824940 | ZNF512 | C | T | 0.84 | 1.81 × 10−7 | 1.089 | 0.087 |
| 2p23.3 | rs62138969 | 2:27828292 | ZNF512 | A | G | 0.84 | 2.10 × 10−7 | 1.088 | 0.090 |
| 2p23.3 | rs6547734 | 2:27830990 | ZNF512 | T | G | 0.84 | 1.52 × 10−7 | 1.088 | 0.090 |
| 2p23.3 | rs6718128 | 2:27833752 | ZNF512 | T | C | 0.84 | 3.19 × 10−7 | 1.088 | 0.091 |
| 2p23.3 | rs4665999 | 2:27834444 | ZNF512 | C | T | 0.84 | 1.04 × 10−7 | 1.088 | 0.090 |
| 2p23.3 | rs11127071 | 2:27838058 | ZNF512 | A | C | 0.84 | 1.06 × 10−7 | 1.077 | 0.140 |
| 2p23.3 | rs6737921 | 2:27839107 | ZNF512 | C | T | 0.84 | 1.08 × 10−7 | 1.076 | 0.141 |
| 2p23.3 | rs6756238 | 2:27841305 | ZNF512 | A | G | 0.84 | 1.11 × 10−7 | 1.074 | 0.153 |
| 2p23.3 | rs1528404 | 2:27846603 | C | T | 0.84 | 1.56 × 10−7 | 1.062 | 0.312 | |
| 2p23.3 | rs1528403 | 2:27846645 | G | C | 0.85 | 3.86 × 10−7 | 1.059 | 0.336 | |
| 2p23.3 | rs2384654 | 2:27847364 | T | C | 0.84 | 1.78 × 10−7 | 1.059 | 0.335 | |
| 2p23.3 | rs4666005 | 2:27855239 | GPN1 | G | A | 0.84 | 2.14 × 10−7 | 1.072 | 0.237 |
| 2p23.3 | rs2384659 | 2:27856793 | GPN1 | A | G | 0.84 | 6.71 × 10−8 | 1.072 | 0.236 |
| 2p23.3 | rs4666009 | 2:27857992 | GPN1 | A | T | 0.84 | 1.75 × 10−7 | 1.073 | 0.235 |
| 2p23.3 | rs56001553 | 2:27859926 | GPN1 | T | C | 1.19 | 2.53 × 10−7 | 1.068 | 0.265 |
| 5p15.33 | rs7726159 | 5:1282319 | TERT | A | C | 1.19 | 2.18 × 10−7 | 1.161 | 6.40 × 10−5 |
| 5p15.33 | rs60622800 | 5:1309904 | MIR4457 | A | G | 0.84 | 6.56 × 10−8 | 1.141 | 0.004 |
| 5p15.33 | rs6554758 | 5:1310152 | MIR4457 | A | G | 0.84 | 6.24 × 10−8 | 1.141 | 0.004 |
| 5p15.33 | rs380286 | 5:1320247 | CLPTM1L | A | G | 1.18 | 3.16 × 10−7 | 1.172 | 0.001 |
| 5p15.33 | rs13178866 | 5:1323212 | CLPTM1L | A | G | 1.19 | 8.00 × 10−8 | 1.149 | 0.002 |
| 5p15.33 | rs466502 | 5:1325767 | CLPTM1L | G | A | 1.19 | 1.31 × 10−7 | 1.152 | 0.002 |
| 5p15.33 | rs465498 | 5:1325803 | CLPTM1L | G | A | 1.19 | 5.48 × 10−8 | 1.156 | 0.001 |
| 5p15.33 | rs452384 | 5:1330840 | CLPTM1L | G | A | 1.19 | 5.74 × 10−8 | 1.156 | 0.001 |
| 5p15.33 | rs370348 | 5:1331219 | CLPTM1L | G | A | 1.19 | 5.83 × 10−8 | 1.149 | 0.002 |
| 9p21.3 | rs944800 | 9:22050898 | CDKN2B-AS1 | A | G | 1.21 | 7.42 × 10−8 | 1.059 | 0.342 |
| 9p21.3 | rs2383205 | 9:22060935 | CDKN2B-AS1 | A | G | 1.20 | 2.55 × 10−7 | 1.036 | 0.542 |
| 9p21.3 | rs1537378 | 9:22061614 | CDKN2B-AS1 | A | G | 1.20 | 1.68 × 10−7 | 1.033 | 0.575 |
| 9p21.3 | rs8181050 | 9:22064391 | CDKN2B-AS1 | G | A | 1.20 | 1.74 × 10−7 | 1.028 | 0.634 |
| 9q34.12 | rs3765566 | 9:133942766 | LAMC3 | G | A | 1.31 | 2.28 × 10−7 | 0.931 | 0.307 |
| 9q34.12 | rs7875478 | 9:133947180 | LAMC3 | T | A | 1.32 | 1.23 × 10−7 | 0.932 | 0.312 |
| 9q34.12 | rs7858204 | 9:133947345 | LAMC3 | A | G | 1.32 | 1.20 × 10−7 | 0.930 | 0.295 |
| 9q34.12 | rs10901348 | 9:133956465 | LAMC3 | A | G | 1.31 | 7.12 × 10−8 | 1.062 | 0.270 |
| 9q34.12 | rs72768533 | 9:133959518 | LAMC3 | G | C | 1.31 | 6.82 × 10−8 | 0.981 | 0.783 |
| 9q34.12 | rs72768534 | 9:133959740 | LAMC3 | C | G | 1.31 | 6.77 × 10−8 | 0.981 | 0.785 |
| 9q34.12 | rs77452476 | 9:133962573 | LAMC3 | G | T | 1.29 | 5.97 × 10−8 | 0.986 | 0.748 |
| 9q34.12 | rs11791030 | 9:133971049 | LOC105376298 | G | T | 1.31 | 1.68 × 10−7 | 0.985 | 0.834 |
| 15q21.2 | rs12910284 | 15:49785916 | LOC105370811 | G | A | 1.19 | 4.28 × 10−7 | 1.029 | 0.438 |
| 15q21.2 | rs10851478 | 15:49829019 | LOC105370811 | G | A | 1.19 | 3.47 × 10−7 | 1.030 | 0.419 |
| 6p21.33 | rs3132451 | 6:31582025 | AIF1 | C | G | 1.25 | 4.59 × 10−7 | 0.866 | 0.029 |
| 6p21.32 | rs9271378 | 6:32587300 | G | A | 1.19 | 2.05 × 10−7 | 1.017 | 0.694 | |
| 6p21.32 | rs17612852 | 6:32620572 | HLA-DQA1 | A | G | 1.23 | 4.21 × 10−7 | 0.929 | 0.078 |
| 6p21.32 | rs4713570 | 6:32626040 | T | C | 1.25 | 1.03 × 10−7 | 0.831 | 0.004 | |
| 6p21.32 | rs1049213 | 6:32627773 | HLA-DQB1 | G | A | 1.24 | 4.65 × 10−7 | 0.910 | 0.018 |
| 6p21.32 | rs3828805 | 6:32636120 | HLA-DQB1 | C | T | 1.23 | 2.83 × 10−7 | 0.910 | 0.217 |
| 6p21.32 | rs3135002 | 6:32668439 | C | A | 1.27 | 1.71 × 10−7 | 0.887 | 0.132 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bau, D.-T.; Liu, T.-Y.; Tsai, C.-W.; Chang, W.-S.; Gu, J.; Yang, J.-S.; Shih, L.-C.; Tsai, F.-J. A Genome-Wide Association Study Identified Novel Genetic Susceptibility Loci for Oral Cancer in Taiwan. Int. J. Mol. Sci. 2023, 24, 2789. https://doi.org/10.3390/ijms24032789
Bau D-T, Liu T-Y, Tsai C-W, Chang W-S, Gu J, Yang J-S, Shih L-C, Tsai F-J. A Genome-Wide Association Study Identified Novel Genetic Susceptibility Loci for Oral Cancer in Taiwan. International Journal of Molecular Sciences. 2023; 24(3):2789. https://doi.org/10.3390/ijms24032789
Chicago/Turabian StyleBau, Da-Tian, Ting-Yuan Liu, Chia-Wen Tsai, Wen-Shin Chang, Jian Gu, Jai-Sing Yang, Liang-Chun Shih, and Fuu-Jen Tsai. 2023. "A Genome-Wide Association Study Identified Novel Genetic Susceptibility Loci for Oral Cancer in Taiwan" International Journal of Molecular Sciences 24, no. 3: 2789. https://doi.org/10.3390/ijms24032789
APA StyleBau, D.-T., Liu, T.-Y., Tsai, C.-W., Chang, W.-S., Gu, J., Yang, J.-S., Shih, L.-C., & Tsai, F.-J. (2023). A Genome-Wide Association Study Identified Novel Genetic Susceptibility Loci for Oral Cancer in Taiwan. International Journal of Molecular Sciences, 24(3), 2789. https://doi.org/10.3390/ijms24032789