
Protective Effects of PEP-1-GSTA2 Protein in Hippocampal Neuronal Cell Damage Induced by Oxidative Stress

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
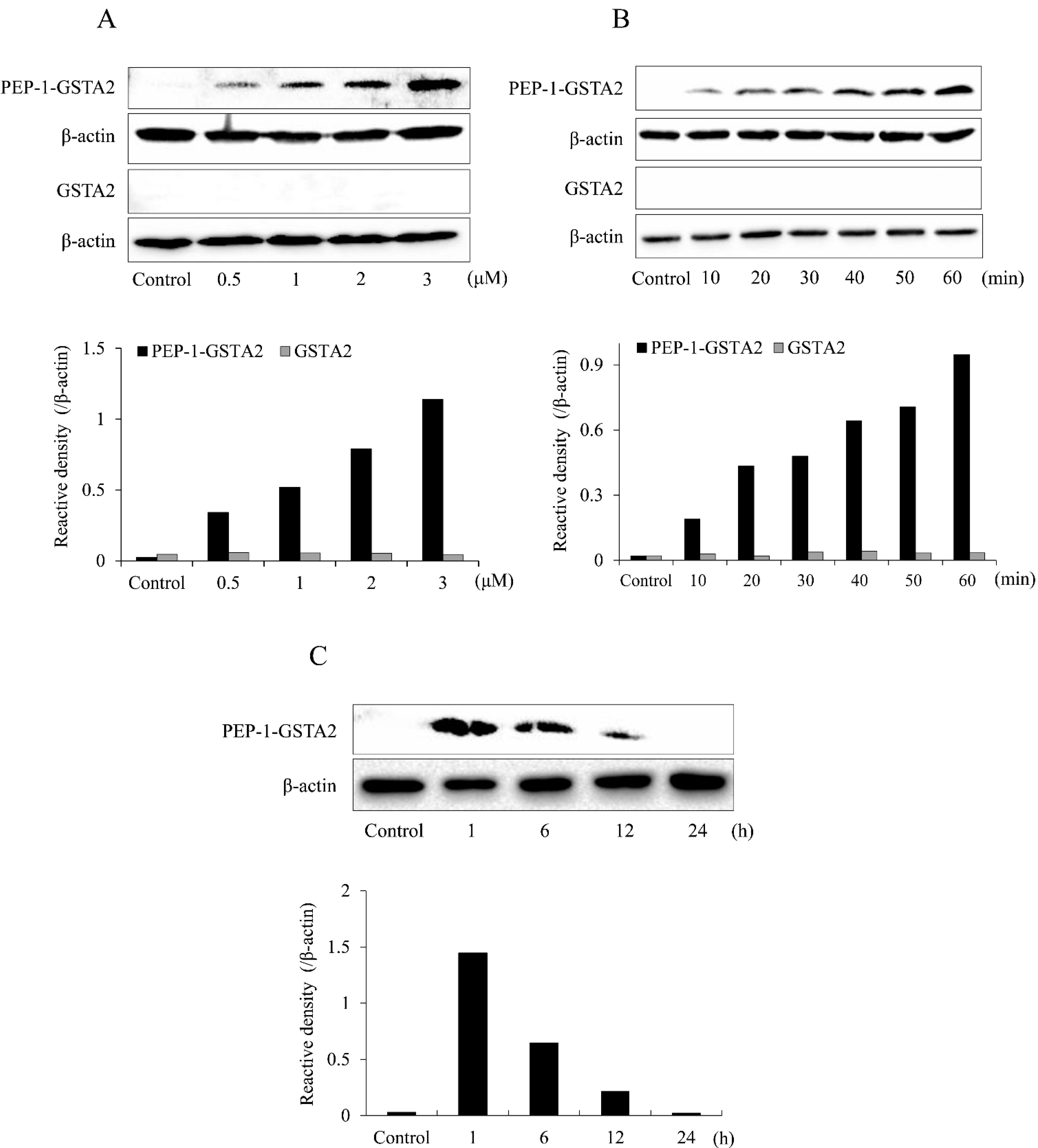
2.1. Preparation and Cell Permeation of Fusion Protein
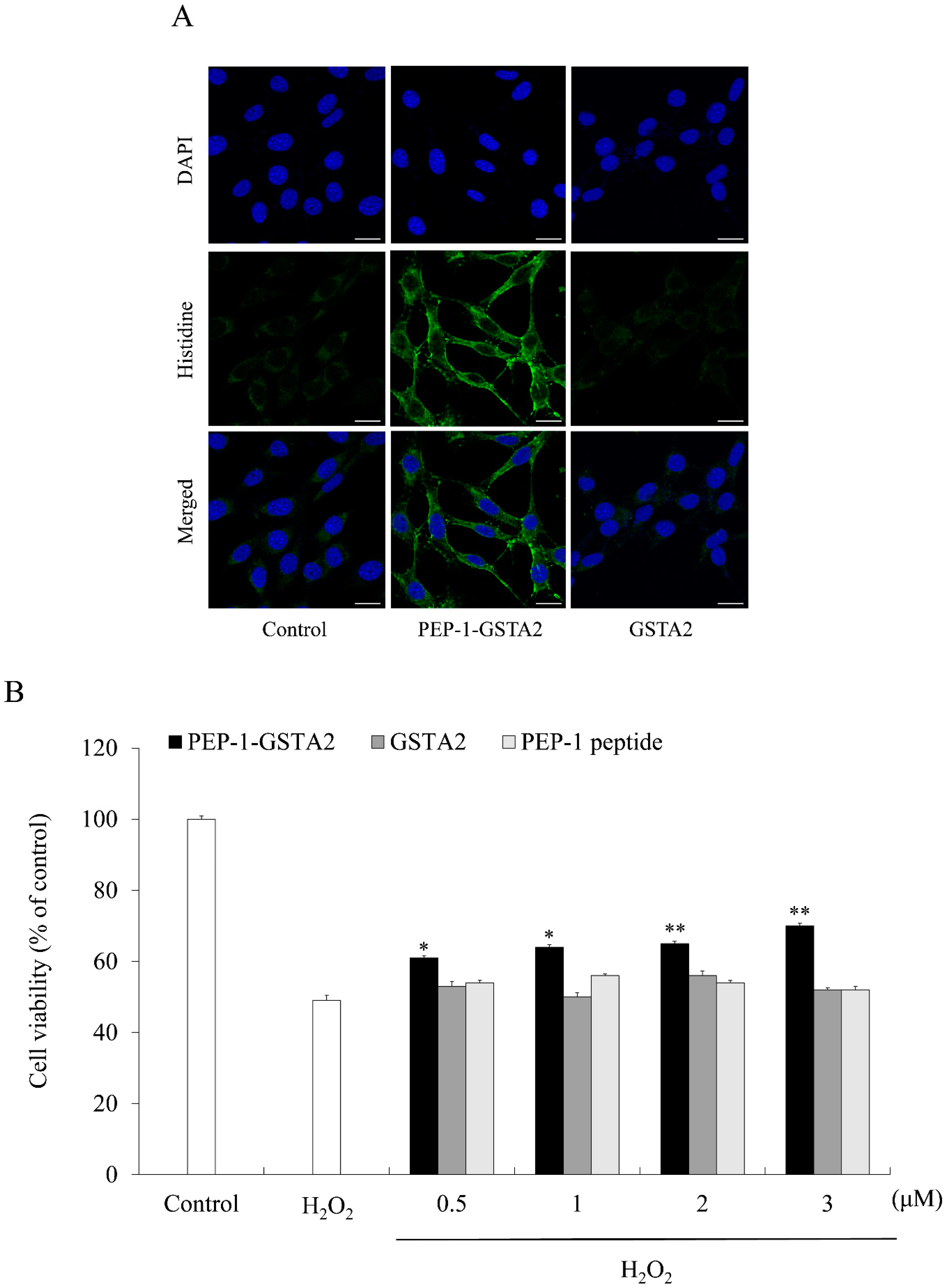
2.2. Effects of PEP-1-GSTA2 on Oxidative Stress
2.3. PEP-1-GSTA2-Activated Phosphorylation of Akt, MAPKs and p65
2.4. PEP-1-GSTA2 Regulated Bcl-2, Bax, Cleaved Caspase-3 and -9 Expressions
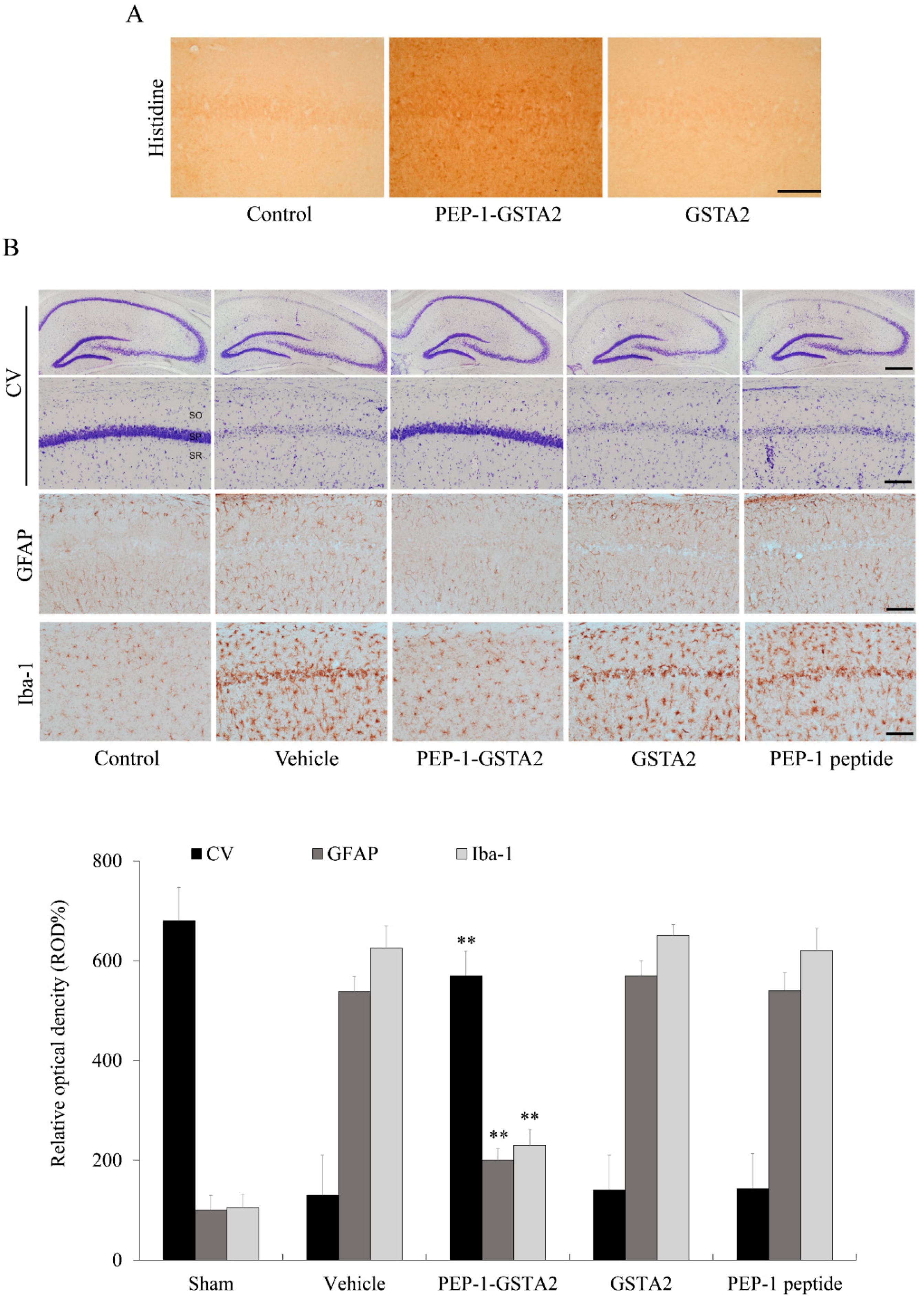
2.5. Effects of PEP-1-GSTA2 in an Ischemic Injury Animal Model
3. Discussion
4. Materials and Methods
4.1. Purification of PEP-1-GSTA2 Proteins
4.2. Cell Culture and Transduction of PEP-1-GSTA2
4.3. Assessment of Cell Viability Using MTT Assay
4.4. DCF-DA and TUNEL Staining
4.5. Experimental Animals
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Glisic, B.; Mihaljevic, I.; Popovic, M.; Zaja, R.; Loncar, J.; Fent, K.; Kovacevic, R.; Smital, T. Characterization of glutathione-S-transferases in zebrafish (Danio rerio). Aquat. Toxicol. 2015, 158, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Tian, J.; Hou, N.; Yu, N.; Zhang, Y.; Liu, Z. Identification, genomic organization and expression pattern of glutathione transferase in Pardosa pseudoannulata. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 32, 100626. [Google Scholar] [CrossRef] [PubMed]
- Henson, K.L.; Stauffer, G.; Gallagher, E.P. Induction of glutathione S-transferase activity and protein expression in brown bullhead (Ameiurus nebulosus) liver by ethoxyquin. Toxicol. Sci. 2001, 62, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Pulford, D.J. The glutathione s-transferase supergene family: Regulation of gst and the contribution of the isoenzymes to cancer chemoprotection and drug resistance. Crit. Rev. Biochem. Mol. Biol. 1995, 30, 445–600. [Google Scholar] [CrossRef] [PubMed]
- Coles, B.F.; Kadlubar, F.F. Human alpha class glutathione Stransferases: Genetic polymorphism, expression, and susceptibility to disease. Methods Enzymol. 2005, 401, 9–42. [Google Scholar]
- Ahn, J.; Gammon, M.D.; Santella, R.M.; Gaudet, M.M.; Britton, J.A.; Teitelbaum, S.L.; Terry, M.B.; Neugut, A.I.; Eng, S.M.; Zhang, Y.; et al. Effects of glutathione Stransferase A1 (GSTA1) genotype and potential modifiers on breast cancer risk. Carcinogenesis 2006, 27, 1876–1882. [Google Scholar] [CrossRef]
- Kang, K.W.; Lee, S.J.; Kim, S.G. Molecular mechanism of Nrf2 activation by oxidative stress. Antioxid. Redox Signal. 2005, 7, 1664–1673. [Google Scholar] [CrossRef]
- Bousova, I.; Kostakova, S.; Matouskova, P.; Bartikova, H.; Szotakova, B.; Skalova, L. Monosodium glutamate-induced obesity changed the expression and activity of glutathione S-transferases in mouse heart and kidney. Pharmazie 2017, 72, 257–259. [Google Scholar]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef]
- Sharma, R.; Yang, Y.; Sharma, A.; Awasthi, S.; Awasthi, Y.C. Antioxidant role of glutathione S-transferases: Protection against oxidant toxicity and regulation of stress-mediated apoptosis. Antioxid. Redox Signal. 2004, 6, 289–300. [Google Scholar] [CrossRef]
- Yang, Y.; Cheng, J.Z.; Singhal, S.S.; Saini, M.; Pandya, U.; Awasthi, S.; Awasthi, Y.C. Role of glutathione S-transferases in protection against lipid peroxidation. J. Biol. Chem. 2001, 276, 19220–19230. [Google Scholar] [CrossRef]
- Yang, Y.; Sharma, R.; Cheng, J.Z.; Saini, M.K.; Ansari, N.H.; Andley, U.P.; Awasthi, S.; Awasthi, Y.C. Protection of HLE B-3 Cells against Hydrogen Peroxide–and Naphthalene-Induced Lipid Peroxidation and Apoptosis by Transfection with hGSTA1 and hGSTA2. Investig. Ophthalmol. Vis. Sci. 2002, 43, 434–445. [Google Scholar]
- Tetlow, N.; Board, P.G. Functional polymorphism of human glutathione transferase A2. Pharmacogenetics 2004, 14, 111–116. [Google Scholar] [CrossRef]
- Gupta, S.; Agarwal, A.; Sharma, R.K. The role of placental oxidative stress and lipid peroxidation in preeclampsia. Obstet. Gynecol. Surv. 2005, 60, 807–816. [Google Scholar] [CrossRef]
- Petito, C.K.; Torres-Munoz, J.; Roberts, B.; Olarte, J.P.; Nowak, T.S., Jr.; Pulsinelli, W.A. DNA fragmentation follows delayed neuronal death in CA1 neurons exposed to transient global ischemia in the rat. J. Cereb. Blood Flow Metab. 1997, 17, 967–976. [Google Scholar] [CrossRef]
- Frantseva, M.V.; Carlen, P.L.; Perez Velazquez, J.L. Dynamics of intracellular calcium and free radical production during ischemia in pyramidal neurons. Free Radic. Biol. Med. 2001, 31, 1216–1227. [Google Scholar] [CrossRef]
- Sayre, L.M.; Smith, M.A.; Perry, G. Chemistry and biochemistry of oxidative stress in neurodegenrative disease. Curr. Med. Chem. 2001, 8, 721–738. [Google Scholar] [CrossRef]
- Floyd, R.A. Role of oxygen free radicals in carcinogenesis and brain ischemia. FASEB J. 1990, 4, 2587–2597. [Google Scholar] [CrossRef]
- Guo, C.; Tong, L.; Xi, M.; Yang, H.; Dong, H.; Wen, A. Neuroprotective effect of calycosin on cerebral ischemia and reperfusion injury in rats. J. Ethnopharmacol. 2012, 144, 768–774. [Google Scholar] [CrossRef]
- Chan, P.H. Reactive oxygen radicals in signaling and damage in the ischemic brain. J. Cereb. Blood Flow Metab. 2001, 21, 2–14. [Google Scholar] [CrossRef]
- Ang, Y.L.; Yong, W.P.; Tan, P. Translating gastric cancer genomics into targeted therapies. Crit. Rev. Oncol. Hematol. 2016, 100, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Choi, E.J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta 2010, 1802, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Shi, L.; Qi, G.; Zhao, S.; Gao, Y.; Li, Y. Gypenoside Protects cardiomyocytes against ischemia-reperfusion injury via the inhibition of mitogen-activated protein kinase mediated nuclear factor kappa B pathway in vitro and in vivo. Front. Pharmacol. 2016, 7, 148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xia, J.; Zhang, Y.; Xiao, F.; Wang, J.; Gao, H.; Liu, Y.; Rong, S.; Yao, Y.; Xu, G.; et al. HMGB1-TLR4 signaling participates in renal ischemia reperfusion injury and could be attenuated by dexamethasone-mediated inhibition of the ERK/NF-kappaB pathway. Am. J. Transl. Res. 2016, 8, 4054–4067. [Google Scholar] [PubMed]
- Van den Berg, A.; Dowdy, S.F. Protein transduction domain delivery of therapeutic macromolecules. Curr. Opin. Biotechnol. 2011, 22, 888–893. [Google Scholar] [CrossRef] [PubMed]
- Wadia, J.S.; Dowdy, S.F. Protein transduction technology. Curr. Opin. Biotechnol. 2002, 13, 52–56. [Google Scholar] [CrossRef]
- Zhou, G.; Shan, P.; Hu, X.; Zheng, X.; Zhou, S. Neuroprotective effect of TAT PTD-Ngb fusion protein on primary cortical neurons against hypoxia-induced apoptosis. Neurol. Sci. 2013, 34, 1771–1778. [Google Scholar] [CrossRef]
- Shin, M.J.; Kim, D.W.; Lee, Y.P.; Ahn, E.H.; Jo, H.S.; Kim, D.S.; Kwon, O.S.; Kang, T.C.; Cho, Y.J.; Park, J.; et al. Tat-glyoxalase protein inhibits against ischemic neuronal cell damage and ameliorates ischemic injury. Free Radic. Biol. Med. 2013, 67, 195–210. [Google Scholar] [CrossRef]
- Kim, M.J.; Park, M.; Kim, D.W.; Shin, M.J.; Son, O.; Jo, H.S.; Yeo, H.J.; Cho, S.B.; Park, J.H.; Lee, C.H.; et al. Transduced PEP-1-PON1 protein regulates microglial activation and dopaminergic neuronal death in a Parkinson’s model. Biomaterials 2015, 64, 45–56. [Google Scholar] [CrossRef]
- Yeo, H.J.; Shin, M.J.; Yeo, E.J.; Choi, Y.J.; Kim, D.W.; Kim, D.S.; Eum, W.S.; Choi, S.Y. Tat-CIAPIN1 inhibits hippocampal neuronal cell damage through the MAPK and apoptotic signaling pathways. Free Radic. Biol. Med. 2019, 135, 68–78. [Google Scholar] [CrossRef]
- Dietz, G.P. Cell-penetrating peptide technology to deliver chaperones and associated factors in diseases and basic research. Curr. Pharm. Biotechnol. 2010, 11, 167–174. [Google Scholar] [CrossRef]
- Kubo, E.; Fatma, N.; Akagi, Y.; Beier, D.R.; Singh, S.P.; Singh, D.P. TAT-mediated PRDX6 protein transduction protects against eye lens epithelial cell death and delays lens opacity. Am. J. Physiol. Cell Physiol. 2008, 294, C842–C855. [Google Scholar] [CrossRef]
- Kim, S.J.; Shin, M.J.; Kim, D.W.; Yeo, H.J.; Yeo, E.J.; Choi, Y.J.; Sohn, E.J.; Han, K.H.; Park, J.; Lee, K.W.; et al. Tat-biliverdin reductase A exerts a protective role in oxidative stress-induced hippocampal neuronal cell damage by regulating the apoptosis and MAPK Signaling. Int. J. Mol. Sci. 2020, 21, 2672. [Google Scholar] [CrossRef]
- Shin, M.J.; Kim, D.W.; Choi, Y.J.; Cha, H.J.; Lee, S.H.; Park, J.; Han, K.H.; Eum, W.S.; Choi, S.Y. PEP-1-GLRX1 protein exhibits anti-inflammatory effects by inhibiting the activation of MAPK and NF-κB pathways in Raw 264.7 cells. BMB Rep. 2020, 53, 106–111. [Google Scholar] [CrossRef]
- Yeo, H.J.; Shin, M.J.; Kim, D.W.; Kwon, H.Y.; Eum, W.S.; Choi, S.Y. Tat-CIAPIN1 protein prevents against cytokine-induced cytotoxicity in pancreatic RINm5F β-cells. BMB Rep. 2021, 54, 458–463. [Google Scholar] [CrossRef]
- Son, Y.; Cheong, Y.K.; Kim, N.H.; Chung, H.T.; Kang, D.G.; Pae, H.O. Mitogen-activated protein kinases and reactive oxygen species: How can ROS activate MAPK pathways? J. Signal Transduct. 2011, 2011, 792639. [Google Scholar] [CrossRef]
- Ichijo, H.; Nishida, E.; Irie, K.; ten Dijke, P.; Saitoh, M.; Moriguchi, T.; Takagi, M.; Matsumoto, K.; Miyazono, K.; Gotoh, Y. Induction of apoptosis by ASK1, a mammalian MAPKKK that activates SAPK/JNK and p38 signaling pathways. Science 1997, 275, 90–94. [Google Scholar] [CrossRef]
- Rowe, J.D.; Nieves, E.; Listowsky, I. Subunit diversity and tissue distribution of human glutathione S-transferases: Interpretations based on electrospray ionization-MS and peptide sequence-specific antisera. Biochem. J. 1997, 325, 481–486. [Google Scholar] [CrossRef]
- Liu, J.; Liu, H.; Zhao, Z.; Wang, J.; Guo, D.; Liu, Y. Regulation of Actg1 and Gsta2 is possible mechanism by which capsaicin alleviates apoptosis in cell model of 6-OHDA-induced Parkinson’s disease. Biosci. Rep. 2020, 40, BSR20191796. [Google Scholar] [CrossRef]
- Ng, K.T.; Yeung, O.W.; Lam, Y.F.; Liu, J.; Liu, H.; Pang, L.; Yang, X.X.; Zhu, J.Y.; Zhang, W.Y.; Lau, M.Y.; et al. Glutathion S-transferase A2 (GSTA2) promotes hepatocellular carcinoma recurrence after liver transplantation through modulating reactive oxygen species metabolism. Cell Death Discov. 2021, 7, 188. [Google Scholar] [CrossRef]
- Lee, J.Y.; Han, C.Y.; Yang, J.W.; Smith, C.; Kim, S.K.; Lee, E.Y.; Kim, S.G.; Kang, K.W. Induction of glutathione transferase in insulin-like growth factor type I receptor-overexpressed hepatoma cells. Mol. Pharmacol. 2007, 72, 1082–1093. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wang, L.L.; Shi, Z.; Ou, X.; Wang, W.; Chen, X.; Liu, G. Transcriptional profiling of liver tissues in chicken embryo at day 16 and 20 using RNA sequencing reveals differential antioxidant enzyme activity. PLoS ONE 2018, 13, e0192253. [Google Scholar] [CrossRef] [PubMed]
- Lyakhovich, V.V.; Vavilin, V.A.; Zenkov, N.K.; Menshchikova, E.B. Active defense under oxidative stress. The antioxidant responsive element. Biochemistry 2006, 71, 962–974. [Google Scholar] [CrossRef] [PubMed]
- Emerit, J.; Edeas, M.; Bricaire, F. Neurodegenerative diseases and oxidative stress. Biomed. Pharmacother. 2004, 58, 39–46. [Google Scholar] [CrossRef]
- Joliot, A.; Prochiantz, A. Transduction peptides: From technology to physiology. Nat. Cell Biol. 2004, 6, 189–196. [Google Scholar] [CrossRef]
- Luo, X.G.; Ma, D.Y.; Wang, Y.; Li, W.; Wang, C.X.; He, Y.Y.; Gu, X.C.; Li, X.M.; Zhou, H.; Zhang, T.C. Fusion with pep-1, a cell-penetrating peptide, enhances the transmembrane ability of human epidermal growth factor. Biosci. Biotechnol. Biochem. 2016, 80, 584–590. [Google Scholar] [CrossRef]
- Morris, M.C.; Depollier, J.; Mery, J.; Heitz, F.; Divita, G. A peptide carrier for the delivery of biologically active proteins into mammalian cells. Nat. Biotechnol. 2001, 19, 1173–1176. [Google Scholar] [CrossRef]
- Awasthi, S.; Srivatava, S.K.; Piper, J.T.; Singhal, S.S.; Chaubey, M.; Awasthi, Y.C. Curcumin protects against 4-hydroxy-2-trans-nonenal–induced cataract formation in rat lenses. Am. J. Clin. Nutr. 1996, 64, 761–766. [Google Scholar] [CrossRef]
- Pandya, U.; Chandra, A.; Awasthi, S.; Jin, G.F.; Piper, J.T.; Godley, B.F.; Awasthi, Y.C. Attenuation of galactose cataract by low levels of dietary curcumin. Nutr. Res. 2000, 20, 515–526. [Google Scholar] [CrossRef]
- Lahair, M.M.; Howe, C.J.; Rodriguez-Mora, O.; McCubrey, J.A.; Franklin, R.A. Molecular pathways leading to oxidative stress-induced phosphorylation of Akt. Antioxid. Redox Signal. 2006, 8, 1749–1756. [Google Scholar] [CrossRef]
- Kwon, S.H.; Hong, S.I.; Kim, J.A.; Jung, Y.H.; Kim, S.Y.; Kim, H.C.; Lee, S.Y.; Jang, C.G. The neuroprotective effects of Lonicera japonica THUNB. against hydrogen peroxide-induced apoptosis via phosphorylation of MAPKs and PI3K/Akt in SH-SY5Y cells. Food Chem. Toxicol. 2011, 49, 1011–1019. [Google Scholar] [CrossRef]
- Hwang, S.L.; Yen, G.C. Modulation of Akt, JNK, and p38 activation is involved in citrus flavonoid-mediated cytoprotection of PC12 cells challenged by hydrogen peroxide. J. Agric. Food Chem. 2009, 57, 2576–2582. [Google Scholar] [CrossRef]
- Yang, B.; Oo, T.N.; Rizzo, V. Lipid rafts mediate H2O2 prosurvival effects in cultured endothelial cells. FASEB J. 2006, 20, 1501–1503. [Google Scholar] [CrossRef]
- Ruffels, J.; Griffin, M.; Dickenson, J.M. Activation of ERK1/2, JNK and PKB by hydrogen peroxide in human SH-SY5Y neuroblastoma cells: Role of EKR1/2 in H2O2-induced cell death. Eur. J. Pharmacol. 2004, 483, 163–173. [Google Scholar] [CrossRef]
- Yano, S.; Morioka, M.; Fukunaga, K.; Kawano, T.; Hara, T.; Kai, Y.; Hamada, J.; Miyamoto, E.; Ushio, Y. Activation of Akt/protein kinase B contributes to induction of ischemic tolerance in the CA1 subfield of gerbil hippocampus. J. Cereb. Blood Flow Metab. 2001, 21, 351–360. [Google Scholar] [CrossRef]
- Fulda, S.; Gorman, A.M.; Hori, O.; Samali, A. Cellular stress responses: Cell survival and cell death. Int. J. Cell Biol. 2010, 2010, 214074. [Google Scholar] [CrossRef]
- Romero, L.; Andrews, K.; Ng, L.; O’Rourke, K.; Maslen, A.; Kirby, G. Human GSTA1-1 reduces c-Jun N-terminal kinase signaling and apoptosis in Caco-2 cells. Biochem. J. 2006, 400, 135–141. [Google Scholar] [CrossRef]
- Gilot, D.; Loyer, P.; Corlu, A.; Glaise, D.; Lagadic-Gossmann, D.; Atfi, A.; Morel, F.; Ichijo, H.; Guguen-Guillouzo, C. Liver protection from apoptosis requires both blockage of initiator caspase activities and inhibition of ASK1/JNK pathway via glutathione S-transferase regulation. J. Biol. Chem. 2002, 277, 49220–49229. [Google Scholar] [CrossRef]
- Pradeep, H.; Diya, J.B.; Shashikumar, S.; Rajanikant, G.K. Oxidative stress-assassin behind the ischemic stroke. Folia. Neuropathol. 2012, 50, 219–230. [Google Scholar] [CrossRef]
- Zhu, Y.; Bu, Q.; Liu, X.; Hu, W.; Wang, Y. Neuroprotective effect of TAT-14-3-3ε fusion protein against cerebral ischemia/reperfusion injury in rats. PLoS ONE 2014, 26, e93334. [Google Scholar] [CrossRef]
- Lim, K.S.; Cha, M.J.; Kim, J.K.; Park, E.J.; Chae, J.W.; Rhim, T.; Hwang, K.C.; Kim, Y.H. Protective effects of protein transduction domain-metallothionein fusion proteins against hypoxia- and oxidative stress-induced apoptosis in an ischemia/reperfusion rat model. J. Control. Release 2013, 169, 306–312. [Google Scholar] [CrossRef]
- Ito, D.; Tanaka, K.; Suzuki, S.; Dembo, T.; Fukuuchi, Y. Enhanced expression of Iba1, ionized calcium-binding adapter molecule 1, after transient focal cerebral ischemia in rat brain. Stroke 2001, 32, 1208–1215. [Google Scholar] [CrossRef]
- Chen, Y.; Swanson, R.A. Astrocytes and brain injury. J. Cereb. Blood Flow Metab. 2003, 23, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Go, S.; Park, J.; Rahman, S.; Jin, J.; Choi, I.; Kim, J. Adipogenic function of tetranectin mediated by enhancing mitotic clonal expansion via ERK signaling. BMB Rep. 2021, 54, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Cho, I.J.; Kim, E.O.; Lee, D.G.; Jung, D.H.; Ki, S.H.; Ku, S.K.; Kim, S.C. Hemistepsin A inhibits T0901317-induced lipogenesis in the liver. BMB Rep. 2021, 54, 106–111. [Google Scholar] [CrossRef]
- Koo, B.H.; Lee, J.; Jin, Y.; Lim, H.K.; Ryoo, S. Arginase inhibition by rhaponticin increases L-arginine concentration that contributes to Ca2+-dependent eNOS activation. BMB Rep. 2021, 54, 516–521. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.J.; Shin, M.J.; Youn, G.S.; Park, J.H.; Yeo, H.J.; Yeo, E.J.; Kwon, H.J.; Lee, L.R.; Kim, N.Y.; Kwon, S.Y.; et al. Protective Effects of PEP-1-GSTA2 Protein in Hippocampal Neuronal Cell Damage Induced by Oxidative Stress. Int. J. Mol. Sci. 2023, 24, 2767. https://doi.org/10.3390/ijms24032767
Choi YJ, Shin MJ, Youn GS, Park JH, Yeo HJ, Yeo EJ, Kwon HJ, Lee LR, Kim NY, Kwon SY, et al. Protective Effects of PEP-1-GSTA2 Protein in Hippocampal Neuronal Cell Damage Induced by Oxidative Stress. International Journal of Molecular Sciences. 2023; 24(3):2767. https://doi.org/10.3390/ijms24032767
Chicago/Turabian StyleChoi, Yeon Joo, Min Jea Shin, Gi Soo Youn, Jung Hwan Park, Hyeon Ji Yeo, Eun Ji Yeo, Hyun Jung Kwon, Lee Re Lee, Na Yeon Kim, Su Yeon Kwon, and et al. 2023. "Protective Effects of PEP-1-GSTA2 Protein in Hippocampal Neuronal Cell Damage Induced by Oxidative Stress" International Journal of Molecular Sciences 24, no. 3: 2767. https://doi.org/10.3390/ijms24032767
APA StyleChoi, Y. J., Shin, M. J., Youn, G. S., Park, J. H., Yeo, H. J., Yeo, E. J., Kwon, H. J., Lee, L. R., Kim, N. Y., Kwon, S. Y., Jung, H. Y., Cho, Y.-J., Kim, D. W., Park, J., Han, K. H., Lee, K. W., Park, J. K., Lee, C. H., Eum, W. S., & Choi, S. Y. (2023). Protective Effects of PEP-1-GSTA2 Protein in Hippocampal Neuronal Cell Damage Induced by Oxidative Stress. International Journal of Molecular Sciences, 24(3), 2767. https://doi.org/10.3390/ijms24032767