
Enhanced Antibacterial Activity of Substituted Derivatives of NCR169C Peptide

 ,
,
Abstract
1. Introduction
2. Results
2.1. NCR169C and Its Derivates Are Potential Antimicrobial Agents
2.2. Effect of the Tryptophan Residue Modifications on Antimicrobial Activity
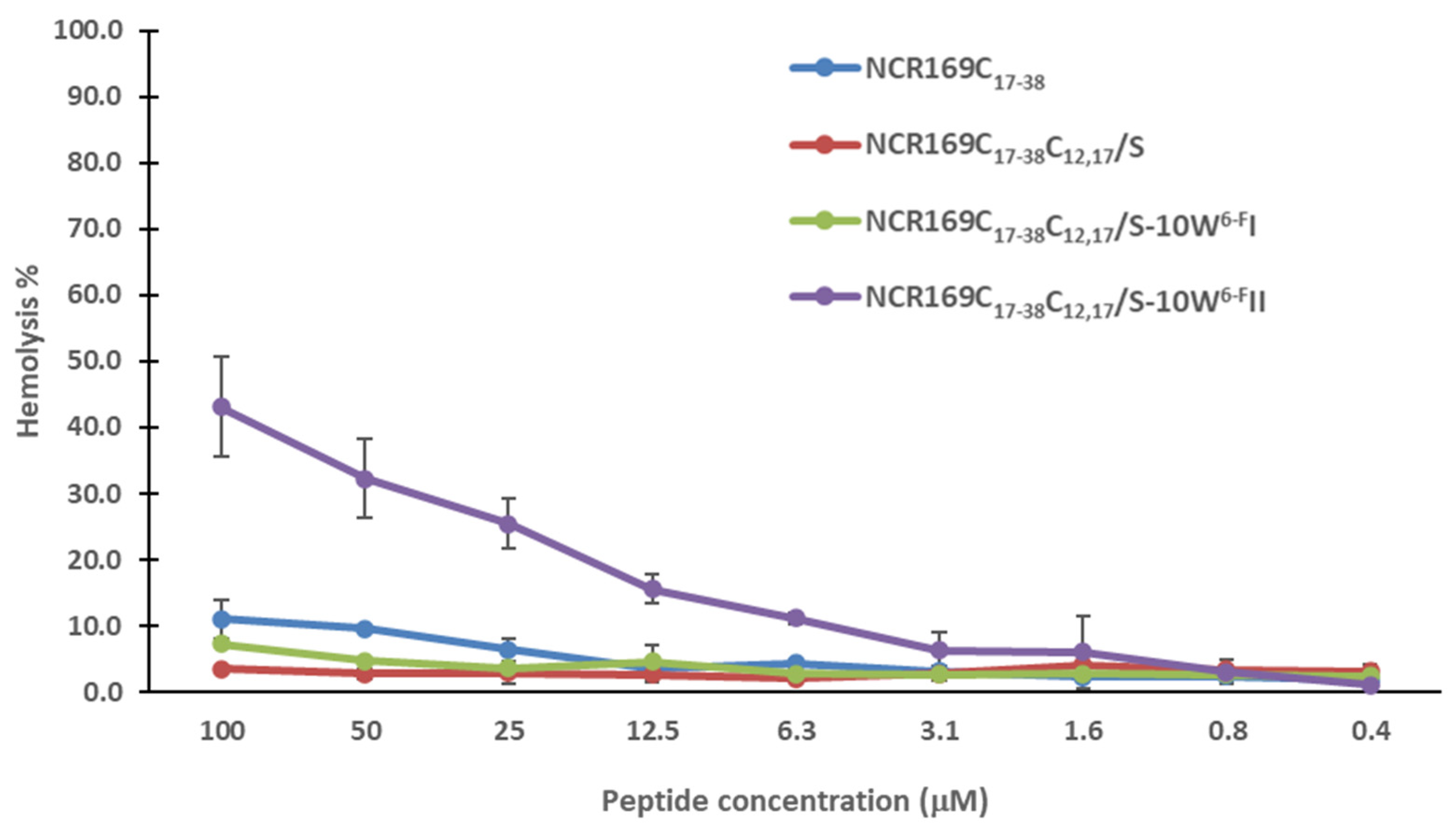
2.3. NCR169C17–38 and Its Derivatives Do Not Provoke Hemolysis of Human Red Blood Cells
3. Discussion
4. Materials and Methods
4.1. Chemical Synthesis of Peptides
4.2. Bacterial Strains
4.3. Antimicrobial Activity
4.4. Hemolysis Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shankar, P. Book Review: Tackling Drug-Resistant Infections Globally. Arch. Pharm. Pract. 2016, 7, 110–111. [Google Scholar] [CrossRef]
- Dallon, E.K. Exploration of Antimicrobial Activity in Natural Peptides and High-Throughput Discovery of Synthetic Peptides. Ph.D. Thesis, Brigham Young University, Provo, UT, USA, 2018. [Google Scholar]
- WHO. WHO Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report 2021; WHO: Geneva, Switzerland, 2021; ISBN 9789240005587. [Google Scholar]
- Bhatia, P.; Sharma, A.; George, A.J.; Anvitha, D.; Kumar, P.; Dwivedi, V.P.; Chandra, N.S. Antibacterial Activity of Medicinal Plants against ESKAPE: An Update. Heliyon 2021, 7, e06310. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.; Mishra, S.K.; Shrestha, A. Characterisation of Eskape Pathogens with Special Reference to Multidrug Resistance and Biofilm Production in a Nepalese Hospital. Infect. Drug Resist. 2021, 14, 2201–2212. [Google Scholar] [CrossRef] [PubMed]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial Resistance: A Global Multifaceted Phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Afshinnekoo, E.; Bhattacharya, C.; Burguete-García, A.; Castro-Nallar, E.; Deng, Y.; Desnues, C.; Dias-Neto, E.; Elhaik, E.; Iraola, G.; Jang, S.; et al. COVID-19 Drug Practices Risk Antimicrobial Resistance Evolution. Lancet Microbe 2021, 2, e135–e136. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Getahun, H.; Smith, I.; Trivedi, K.; Paulin, S.; Balkhy, H.H. Tackling Antimicrobial Resistance in the Community. Bull. World Health Organ. 2020, 98, 442. [Google Scholar] [CrossRef]
- Rawson, T.M.; Moore, L.S.P.; Zhu, N.; Ranganathan, N.; Skolimowska, K.; Gilchrist, M.; Satta, G.; Cooke, G.; Holmes, A. Bacterial and Fungal Coinfection in Individuals with Coronavirus: A Rapid Review to Support COVID-19 Antimicrobial Prescribing. Clin. Infect. Dis. 2020, 71, 2459–2468. [Google Scholar] [CrossRef]
- Essa, E.E.; Hamza, D.; Khalil, M.M.H.; Zaher, H.; Salah, D.; Alnemari, A.M.; Rady, M.H.; Mo`men, S.A.A. The Antibacterial Activity of Egyptian Wasp Chitosan-Based Nanoparticles against Important Antibiotic-Resistant Pathogens. Molecules 2022, 27, 7189. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente-Núñez, C.; Silva, O.N.; Lu, T.K.; Franco, O.L. Antimicrobial Peptides: Role in Human Disease and Potential as Immunotherapies. Pharmacol. Ther. 2017, 178, 132–140. [Google Scholar] [CrossRef]
- Wang, G.; Zietz, C.M.; Mudgapalli, A.; Wang, S.; Wang, Z. The Evolution of the Antimicrobial Peptide Database over 18 Years: Milestones and New Features. Protein Sci. 2022, 31, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Gergely, G.M.; Kereszt, A.; Kondorosi, É.; Mergaert, P. Natural Roles of Antimicrobial Peptides in Microbes, Plants and Animals. Res. Microbiol. 2011, 162, 363–374. [Google Scholar] [CrossRef]
- Mikuláss, K.R.; Nagy, K.; Bogos, B.; Szegletes, Z.; Kovács, E.; Farkas, A.; Váró, G.; Kondorosi, É.; Kereszt, A. Antimicrobial Nodule-Specific Cysteine-Rich Peptides Disturb the Integrity of Bacterial Outer and Inner Membranes and Cause Loss of Membrane Potential. Ann. Clin. Microbiol. Antimicrob. 2016, 15, 43. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Björn, C.; Ekblom, J. Antimicrobial Peptides as Therapeutic Agents: Opportunities and Challenges. Crit. Rev. Biotechnol. 2020, 40, 978–992. [Google Scholar] [CrossRef]
- Mishra, B.; Reiling, S.; Zarena, D.; Wang, G. Host Defense Antimicrobial Peptides as Antibiotics: Design and Application Strategies. Curr. Opin. Chem. Biol. 2017, 38, 87–96. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial Peptides of Multicellular Organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility in Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Mergaert, P. Role of Antimicrobial Peptides in Controlling Symbiotic Bacterial Populations. Nat. Prod. Rep. 2018, 35, 336–356. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Lu, T.K. Development and Challenges of Antimicrobial Peptides for Therapeutic Applications. Antibiotics 2020, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Roscia, G.; Falciani, C.; Bracci, L.; Pini, A. The Development of Antimicrobial Peptides as New Antibacterial Drugs. Curr. Protein Pept. Sci. 2013, 14, 641–649. [Google Scholar] [CrossRef]
- Lei, J.; Sun, L.C.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q.Y. The Antimicrobial Peptides and Their Potential Clinical Applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Sathoff, A.E.; Samac, D.A. Antibacterial Activity of Plant Defensins. Mol. Plant-Microbe Interact. 2019, 32, 507–514. [Google Scholar] [CrossRef]
- Kondorosi, E.; Mergaert, P.; Kereszt, A. A Paradigm for Endosymbiotic Life: Cell Differentiation of Rhizobium Bacteria Provoked by Host Plant Factors. Annu. Rev. Microbiol. 2013, 67, 611–628. [Google Scholar] [CrossRef] [PubMed]
- Horvath, B.; Domonkos, A.; Kereszt, A.; Szucs, A.; Abraham, E.; Ayaydin, F.; Boka, K.; Chen, Y.; Chen, R.; Murray, J.D.; et al. Loss of the Nodule-Specific Cysteine Rich Peptide, NCR169, Abolishes Symbiotic Nitrogen Fixation in the Medicago Truncatula Dnf7 Mutant. Proc. Natl. Acad. Sci. USA 2015, 112, 15232–15237. [Google Scholar] [CrossRef]
- Szerencsés, B.; Gácser, A.; Endre, G.; Domonkos, I.; Tiricz, H.; Vágvölgyi, C.; Szolomajer, J.; Howan, D.H.O.; Tóth, G.K.; Pfeiffer, I.; et al. Symbiotic NCR Peptide Fragments Affect the Viability, Morphology and Biofilm Formation of Candida Species. Int. J. Mol. Sci. 2021, 22, 3666. [Google Scholar] [CrossRef] [PubMed]
- Jenei, S.; Tiricz, H.; Szolomájer, J.; Tímár, E.; Klement, É.; Al Bouni, M.A.; Lima, R.M.; Kata, D.; Harmati, M.; Buzás, K.; et al. Potent Chimeric Antimicrobial Derivatives of the Medicago Truncatula NCR247 Symbiotic Peptide. Front. Microbiol. 2020, 11, 270. [Google Scholar] [CrossRef]
- Farkas, A.; Maróti, G.; Dürgo, H.; Györgypál, Z.; Lima, R.M.; Medzihradszky, K.F.; Kereszt, A.; Mergaert, P.; Kondorosi, É. Medicago Truncatula Symbiotic Peptide NCR247 Contributes to Bacteroid Differentiation through Multiple Mechanisms. Proc. Natl. Acad. Sci. USA. 2014, 111, 5183–5188. [Google Scholar] [CrossRef]
- Lima, R.M.; Rathod, B.B.; Tiricz, H.; Howan, D.H.O.; Mohamad, A.A.B.; Jenei, S.; Timár, E.; Endre, G.; Tóth, G.K.; Kondorosi, É. Legume Plant Peptides as Sources of Novel Antimicrobial Molecules Against Human Pathogens. Front. Mol. Biosci. 2022, 9, 870460. [Google Scholar] [CrossRef]
- Van de Velde, W.; Zehirov, G.; Szatmari, A.; Debreczeny, M.; Ishihara, H.; Kevei, Z.; Farkas, A.; Mikulass, K.; Nagy, A.; Tiricz, H.; et al. Plant Peptides Govern Terminal Differentiation of Bacteria in Symbiosis. Science 2010, 327, 1122–1126. [Google Scholar] [CrossRef] [PubMed]
- Isozumi, N.; Masubuchi, Y.; Imamura, T.; Mori, M.; Koga, H.; Ohki, S. Structure and Antimicrobial Activity of NCR169, a Nodule-Specific Cysteine-Rich Peptide of Medicago Truncatula. Sci. Rep. 2021, 11, 9923. [Google Scholar] [CrossRef]
- Lindström, K.; Mousavi, S.A. Effectiveness of Nitrogen Fixation in Rhizobia. Microb. Biotechnol. 2020, 13, 1314–1335. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Niu, C.; Zhou, Y.; Xue, X.; Meng, J.; Luo, X.; Hou, Z. The Disulfide Bond of the Peptide Thanatin Is Dispensible for Its Antimicrobial Activity in Vivo and in Vitro. Antimicrob. Agents Chemother. 2016, 60, 4283–4289. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Hoover, D.M.; Yang, D.; Boulègue, C.; Santamaria, F.; Oppenheim, J.J.; Lubkowski, J.; Lu, W. Engineering Disulfide Bridges to Dissect Antimicrobial and Chemotactic Activities of Human β-Defensin 3. Proc. Natl. Acad. Sci. USA 2003, 100, 8880–8885. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.E.; Aumelas, A.; Charnet, P.; Roumestand, C.; Chiche, L.; Despaux, E.; Grassy, G.; Calas, B.; Chavanieu, A. Change in Membrane Permeability Induced by Protegrin 1: Implication of Disulphide Bridges for Pore Formation. FEBS Lett. 1996, 383, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, B.O.; Wu, Z.; Nuding, S.; Groscurth, S.; Marcinowski, M.; Beisner, J.; Buchner, J.; Schaller, M.; Stange, E.F.; Wehkamp, J. Reduction of Disulphide Bonds Unmasks Potent Antimicrobial Activity of Human β 2-Defensin 1. Nature 2011, 469, 419–423. [Google Scholar] [CrossRef]
- Sharma, H.; Nagaraj, R. Human β-Defensin 4 with Non-Native Disulfide Bridges Exhibit Antimicrobial Activity. PLoS ONE 2015, 10, 14–18. [Google Scholar] [CrossRef]
- Wang, Q.; Lv, Y.; Pang, J.; Li, X.; Lu, X.; Wang, X.; Hu, X.; Nie, T.; Yang, X.; Xiong, Y.Q.; et al. In Vitro and in Vivo Activity of D-Serine in Combination with β-Lactam Antibiotics against Methicillin-Resistant Staphylococcus aureus. Acta Pharm. Sin. B 2019, 9, 496–504. [Google Scholar] [CrossRef]
- Makino, Y.; Oe, C.; Iwama, K.; Suzuki, S.; Nishiyama, A.; Hasegawa, K.; Okuda, H.; Hirata, K.; Ueno, M.; Kawaji, K.; et al. Serine Hydroxymethyltransferase as a Potential Target of Antibacterial Agents Acting Synergistically with One-Carbon Metabolism-Related Inhibitors. Commun. Biol. 2022, 5, 619. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Wang, X.; Dong, Y.; Xing, C.; Chen, X.; Li, L.; Dong, C.; Li, Y. Effects of l -Serine on Macrolide Resistance in Streptococcus suis. Microbiol. Spectr. 2022, 10, 4. [Google Scholar] [CrossRef]
- Feng, X.; Jin, S.; Wang, M.; Pang, Q.; Liu, C.; Liu, R.; Wang, Y.; Yang, H.; Liu, F.; Liu, Y. The Critical Role of Tryptophan in the Antimicrobial Activity and Cell Toxicity of the Duck Antimicrobial Peptide DCATH. Front. Microbiol. 2020, 11, 1146. [Google Scholar] [CrossRef]
- Chan, D.I.; Prenner, E.J.; Vogel, H.J. Tryptophan- and Arginine-Rich Antimicrobial Peptides: Structures and Mechanisms of Action. Biochim. Biophys. Acta Biomembr. 2006, 1758, 1184–1202. [Google Scholar] [CrossRef]
- Bi, X.; Wang, C.; Ma, L.; Sun, Y.; Shang, D. Investigation of the Role of Tryptophan Residues in Cationic Antimicrobial Peptides to Determine the Mechanism of Antimicrobial Action. J. Appl. Microbiol. 2013, 115, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.K.; Choi, J.; Moon, E.; Baek, K.H. Tryptophan-Rich and Proline-Rich Antimicrobial Peptides. Molecules 2018, 23, 815. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Shao, J.; Li, Q.; Van Heel, A.J.; De Vries, M.P.; Broos, J.; Kuipers, O.P. Incorporation of Tryptophan Analogues into the Lantibiotic Nisin. Amino Acids 2016, 48, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Genapathy, S.; Fischer, P.M.; Chan, W.C. A Facile Approach to Tryptophan Derivatives for the Total Synthesis of Argyrin Analogues. Org. Biomol. Chem. 2014, 12, 9764–9768. [Google Scholar] [CrossRef]
- Inoue, M.; Sumii, Y.; Shibata, N. Contribution of Organofluorine Compounds to Pharmaceuticals. ACS Omega 2020, 5, 10633–10640. [Google Scholar] [CrossRef] [PubMed]
- Salwiczek, M.; Nyakatura, E.K.; Gerling, U.I.M.; Ye, S.; Koksch, B. Fluorinated Amino Acids: Compatibility with Native Protein Structures and Effects on Protein–Protein Interactions. Chem. Soc. Rev. 2012, 41, 2135–2171. [Google Scholar] [CrossRef] [PubMed]
- Crowe, C.; Molyneux, S.; Sharma, S.V.; Zhang, Y.; Gkotsi, D.S.; Connaris, H.; Goss, R.J.M. Halogenases: A Palette of Emerging Opportunities for Synthetic Biology-Synthetic Chemistry and C-H Functionalisation. Chem. Soc. Rev. 2021, 50, 9443–9481. [Google Scholar] [CrossRef] [PubMed]
- Marsh, E.N.G.; Buer, B.C.; Ramamoorthy, A. Fluorine—A New Element in the Design of Membrane-Active Peptides. Mol. Biosyst. 2009, 5, 1143–1147. [Google Scholar] [CrossRef]
- Meng, H.; Kumar, K. Antimicrobial Activity and Protease Stability of Peptides Containing Fluorinated Amino Acids. J. Am. Chem. Soc. 2007, 129, 15615–15622. [Google Scholar] [CrossRef] [PubMed]
- Mardirossian, M.; Rubini, M.; Adamo, M.F.A.; Scocchi, M.; Saviano, M.; Tossi, A.; Gennaro, R.; Caporale, A. Natural and Synthetic Halogenated Amino Acids—Structural and Bioactive Features in Antimicrobial Peptides and Peptidomimetics. Molecules 2021, 26, 7401. [Google Scholar] [CrossRef] [PubMed]
- Scherer, E.M.; Scherer, E.M. Inquiry: The University of Arkansas Undergraduate Research Properties of Modified Tryptophans in a Membrane-Spanning Channel. Inq. Univ. Ark. Undergrad. Res. J. 2002, 3, 20. [Google Scholar]
- Füller, J.J.; Röpke, R.; Krausze, J.; Rennhack, K.E.; Daniel, N.P.; Blankenfeldt, W.; Schulz, S.; Jahn, D.; Moser, J. Biosynthesis of Violacein, Structure and Function of L-Tryptophan Oxidase VioA from Chromobacterium violaceum. J. Biol. Chem. 2016, 291, 20068–20084. [Google Scholar] [CrossRef] [PubMed]
- Katragadda, M.; Lambris, J.D. Expression of Compstatin in Escherichia Coli: Incorporation of Unnatural Amino Acids Enhances Its Activity. Protein Expr. Purif. 2006, 47, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Letzel, T. How to Couple and Handle Liquid Chromatography with Mass Spectrometry. In Protein and peptide Analysis by LC-MS: Experimental Strategies; Royal Society of Chemistry: Piccadilly, London, 2011; pp. 11–25. ISBN 9781849733144. [Google Scholar]
- Zhang, G.; Annan, R.S.; Carr, S.A.; Neubert, T.A. Overview of Peptide and Protein Analysis by Mass Spectrometry. Curr. Protoc. Protein Sci. 2010, 62, 16.1.1–16.1.30. [Google Scholar] [CrossRef]
- Trauger, S.A.; Webb, W.; Siuzdak, G. Peptide and Protein Analysis with Mass Spectrometry. Spectroscopy 2002, 16, 15–28. [Google Scholar] [CrossRef]


{kind=link}
| A. | ||||||||
| Peptides | Amino Acid Sequence | Molecular Mass Calculated | Molecular Mass Experimental 1 | |||||
| NCR169C17–38 | KSKKPLFKIWKCVENVCVLWYK | 2739.4 | 913.65 (M + 3H)3+ | |||||
| NCR169C17–38C12,17/S | KSKKPLFKIWKSVENVSVLWYK | 2707.3 | 903.6 (M + 3H)3+ | |||||
| NCR169C17–38W10,20/A | KSKKPLFKIAKCVENVCVLAYK | 2509.1 | 837.3 (M + 3H)3+ | |||||
| NCR169C17–38W10,20/A,C12,17/S | KSKKPLFKIAKSVENVSVLAYK | 2477 | 826.8 (M + 3H)3+ | |||||
| NCR169C17–38W10,20C12,17/A | KSKKPLFKIAKAVENVAVLAYK | 2445 | 815.7 (M + 3H)3+ | |||||
| B. | ||||||||
| Peptides | pI 2 | Hydrophobicity 2 | GRAVY 2 | Boman Index 3 (kcal/mol) | ||||
| NCR169C17–38 | 10.1 | 48.27 | −0.15 | 0.6 | ||||
| NCR169C17–38C12,17/S | 10.6 | 47.64 | −0.45 | 1.02 | ||||
| NCR169C17–38W10,20/A | 10.1 | 38.74 | 0.09 | 0.64 | ||||
| NCR169C17–38W10,20/A,C12,17/S | 10.6 | 38.04 | −0.21 | 1.07 | ||||
| NCR169C17–38W10,20C12,17/A | 10.6 | 41.04 | 0.03 | 0.59 | ||||
| C. | ||||||||
| Peptides | E. f. | S. a. | K. p. | A. b. | P. a. | E. c. | L. m. | S. e. |
| NCR169C17–38 | 6.3 | 3.1 | 3.1 | 3.1 | 3.1 | 1.6 | 3.1 | 3.1 |
| NCR169C17–38C12,17/S | 3.1 | 1.6 | 3.1 | 1.6 | 3.1 | 1.6 | 3.1 | 1.6 |
| NCR169C17–38W10,20/A | - | - | 12.5 | 3.1 | 3.1 | 3.1 | 25 | - |
| NCR169C17–38W10,20/A,C12,17/S | 25 | 6.3 | 25 | 12.5 | 3.1 | 6.3 | 3.1 | 6.3 |
| NCR169C17–38W10,20C12,17/A | - | 6.3 | 25 | 12.5 | 3.1 | 6.3 | 6.3 | 6.3 |
| D. | ||||||||
| Peptides | E.c. ATTC 25922 | E.c. ATTC 35218 | S.a. ATTC 25923 | |||||
| NCR169C17–38 | 1.6 | 1.6 | 3.1 | |||||
| NCR169C17–38C12,17/S | 1.6 | 3.1 | 3.1 | |||||
| Peptide Analogs | Amino Acid Sequence | Molecular Mass Calculated | Molecular Mass Experimental |
|---|---|---|---|
| NCR169C17–38C12,17/S-10W5-Me I * | KSKKPLFKIW5-MeKSVENVSVLWYK | 2721.26 | 907.6 (M + 3H)3+ |
| NCR169C17–38C12,17/S-10W5-Me II * | KSKKPLFKIW5-MeKSVENVSVLWYK | 2721.26 | 907.8 (M + 3H)3+ |
| NCR169C17–38C12,17/S-20W5-Me I | KSKKPLFKIWKSVENVSVLW5-Me YK | 2721.26 | 908.8 (M + 3H)3+ |
| NCR169C17–38C12,17/S-20W5-Me II | KSKKPLFKIWKSVENVSVLW5-Me YK | 2721.26 | 907.8 (M + 3H)3+ |
| NCR169C17–38C12,17/S-10W5-F-L | KSKKPLFKIW5-F-L KSVENVSVLWYK | 2725.47 | 909.6 (M + 3H)3+ |
| NCR169C17–38C12,17/S-10W5-F-D | KSKKPLFKIW5-F-D KSVENVSVLWYK | 2725.47 | 909.7 (M + 3H)3+ |
| NCR169C17–38C12,17/S-20W5-F-L | KSKKPLFKIWKSVENVSVLW5-F-LYK | 2725.47 | 908.9 (M + 3H)3+ |
| NCR169C17–38C12,17/S-10W6-FI | KSKKPLFKIW6-FKSVENVSVLWYK | 2725.47 | 909.3 (M + 3H)3+ |
| NCR169C17–38C12,17/S-10W6-FII | KSKKPLFKIW6-FKSVENVSVLWYK | 2725.47 | 909.4 (M + 3H)3+ |
| NCR169C17–38C12,17/S-10W7-Aza I | KSKKPLFKIW7-AzaKSVENVSVLWYK | 2706.46 | 903.3 (M + 3H)3+ |
| NCR169C17–38C12,17/S-10W7-Aza II | KSKKPLFKIW7-AzaKSVENVSVLWYK | 2706.46 | 903.6 (M + 3H)3+ |
| NCR169C17–38C12,17/S-20W7-Aza I | KSKKPLFKIWKSVENVSVLW7-AzaYK | 2706.46 | 903.0 (M + 3H)3+ |
| NCR169C17–38C12,17/S-20W7-Aza II | KSKKPLFKIWKSVENVSVLW7-AzaYK | 2706.46 | 903.2 (M + 3H)3+ |
| NCR169C17–38C12,17/S-10W5-MeO I | KSKKPLFKIW5-MeOKSVENVSVLWYK | 2737.49 | 913.1 (M + 3H)3+ |
| NCR169C17–38C12,17/S-10W5-MeO II | KSKKPLFKIW5-MeOKSVENVSVLWYK | 2737.49 | 913.0 (M + 3H)3+ |
| NCR169C17–38C12,17/S-20W5-MeO | KSKKPLFKIWKSVENVSVLW5-MeOYK | 2737.49 | 913.2 (M + 3H)3+ |
| A. | ||||||||
| Peptides | E. f. | S. a. | K. p. | A. b. | P. a. | E. c. | L. m. | S. e. |
| NCR169C17–38C12,17/S-10W5-Me I | 3.1 | 3.1 | 6.3 | 6.3 | 3.1 | 3.1 | 3.1 | 3.1 |
| NCR169C17–38C12,17/S-10W5-Me II | 3.1 | 3.1 | 6.3 | 3.1 | 3.1 | 3.1 | 3.1 | 3.1 |
| NCR169C17–38C12,17/S-20W5-Me I | 12.5 | 6.3 | 12.5 | 6.3 | 6.3 | 6.3 | 6.3 | 6.3 |
| NCR169C17–38C12,17/S-20W5-Me II | 25 | 3.1 | 3.1 | 3.1 | 3.1 | 1.6 | 3.1 | 3.1 |
| NCR169C17–38C12,17/S-10W5-F-L | 1.6 | 1.6 | 3.1 | 1.6 | 1.6 | 1.6 | 1.6 | 1.6 |
| NCR169C17–38C12,17/S-10W5-F-D | 1.6 | 3.1 | 3.1 | 1.6 | 3.1 | 1.6 | 6.3 | 3.1 |
| NCR169C17–38C12,17/S-20W5-F-L | 3.1 | 1.6 | 3.1 | 3.1 | 6.3 | 3.1 | 3.1 | 3.1 |
| NCR169C17–38C12,17/S-10W6-F I | 1.6 | 0.8 | 1.6 | 1.6 | 0.8 | 0.8 | 1.6 | 1.6 |
| NCR169C17–38C12,17/S-10W6-F II | 0.8 | 0.8 | 1.6 | 0.8 | 1.6 | 0.8 | 1.6 | 6.3 |
| NCR169C17–38C12,17/S-10W7-Aza I | 6.3 | 3.1 | 6.3 | 3.1 | 3.1 | 3.1 | 3.1 | 3.1 |
| NCR169C17–38C12,17/S-10W7-Aza II | 3.1 | 3.1 | 6.3 | 6.3 | 3.1 | 3.1 | 3.1 | 3.1 |
| NCR169C17–38C12,17/S-20W7-Aza I | 25 | 6.3 | 6.3 | 6.3 | 6.3 | 3.1 | 25 | 6.3 |
| NCR169C17–38C12,17/S-20W7-Aza II | 12.5 | 12.5 | 12.5 | 12.5 | 12.5 | 12.5 | 12.5 | 12.5 |
| NCR169C17–38C12,17/S-10W5-MeO I | 1.6 | 1.6 | 1.6 | 1.6 | 1.6 | 3.1 | 1.6 | 1.6 |
| NCR169C17–38C12,17/S-10W5-MeO II | 3.1 | 3.1 | 3.1 | 3.1 | 25 | 3.1 | 3.1 | 3.1 |
| NCR169C17–38C12,17/S-20W5-MeO | 3.1 | 3.1 | 3.1 | 3.1 | 0.8 | 3.1 | 3.1 | 3.1 |
| B. | ||||||||
| Peptides | E.c. ATTC 25922 | E.c. ATTC 35218 | S.a. ATTC 25923 | |||||
| NCR169C17–38C12,17/S-10W6-F I | 0.8 | 0.8 | 1.6 | |||||
| NCR169C17–38C12,17/S-10W6-F II_ | 0.8 | 0.8 | 1.6 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Howan, D.H.O.; Jenei, S.; Szolomajer, J.; Endre, G.; Kondorosi, É.; Tóth, G.K. Enhanced Antibacterial Activity of Substituted Derivatives of NCR169C Peptide. Int. J. Mol. Sci. 2023, 24, 2694. https://doi.org/10.3390/ijms24032694
Howan DHO, Jenei S, Szolomajer J, Endre G, Kondorosi É, Tóth GK. Enhanced Antibacterial Activity of Substituted Derivatives of NCR169C Peptide. International Journal of Molecular Sciences. 2023; 24(3):2694. https://doi.org/10.3390/ijms24032694
Chicago/Turabian StyleHowan, Dian H. O., Sándor Jenei, János Szolomajer, Gabriella Endre, Éva Kondorosi, and Gábor K. Tóth. 2023. "Enhanced Antibacterial Activity of Substituted Derivatives of NCR169C Peptide" International Journal of Molecular Sciences 24, no. 3: 2694. https://doi.org/10.3390/ijms24032694
APA StyleHowan, D. H. O., Jenei, S., Szolomajer, J., Endre, G., Kondorosi, É., & Tóth, G. K. (2023). Enhanced Antibacterial Activity of Substituted Derivatives of NCR169C Peptide. International Journal of Molecular Sciences, 24(3), 2694. https://doi.org/10.3390/ijms24032694