
The Intricate Role of Non-Coding RNAs in Sepsis-Associated Disseminated Intravascular Coagulation

 , , , ,
, , , ,  and
and 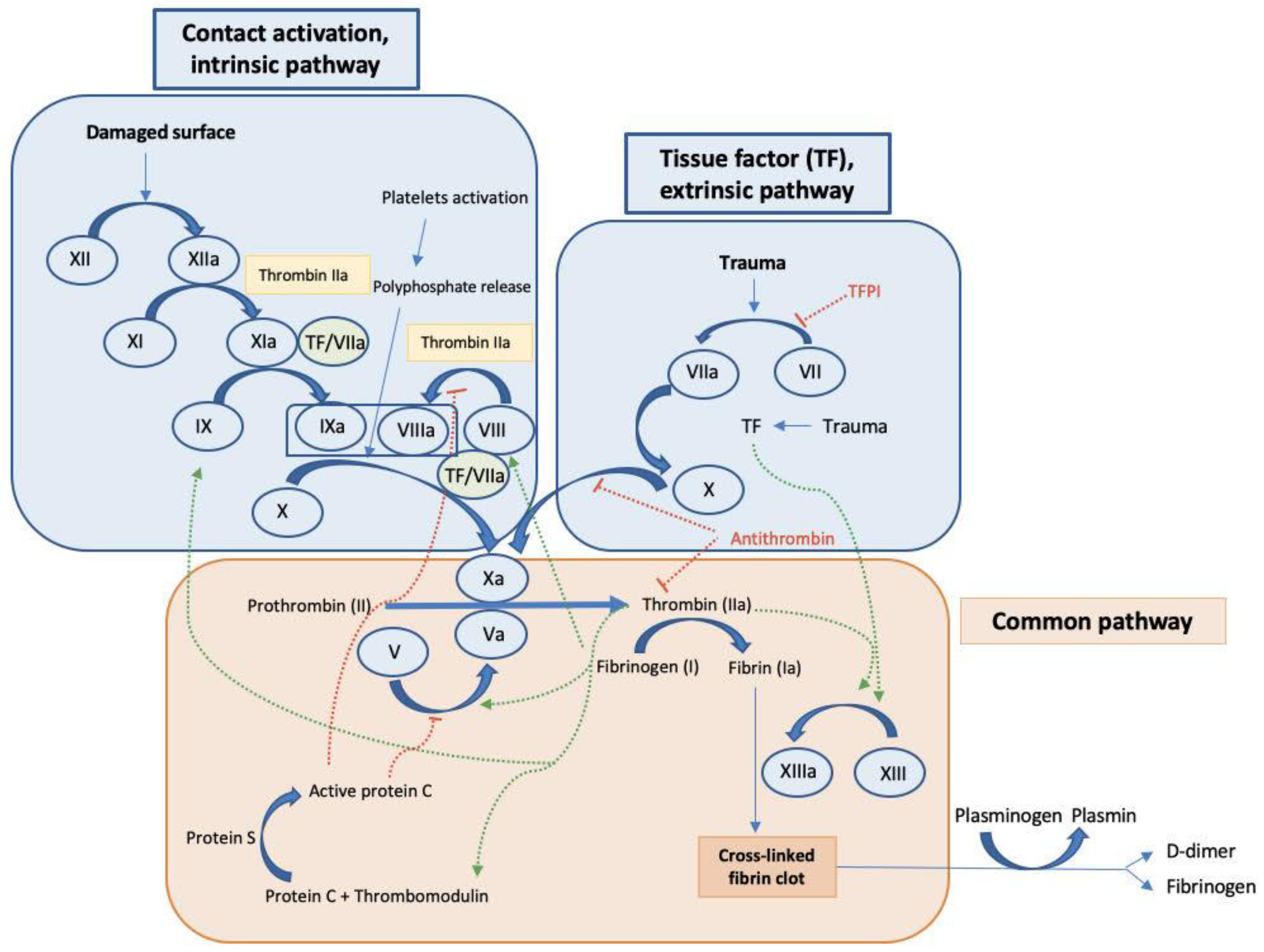{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
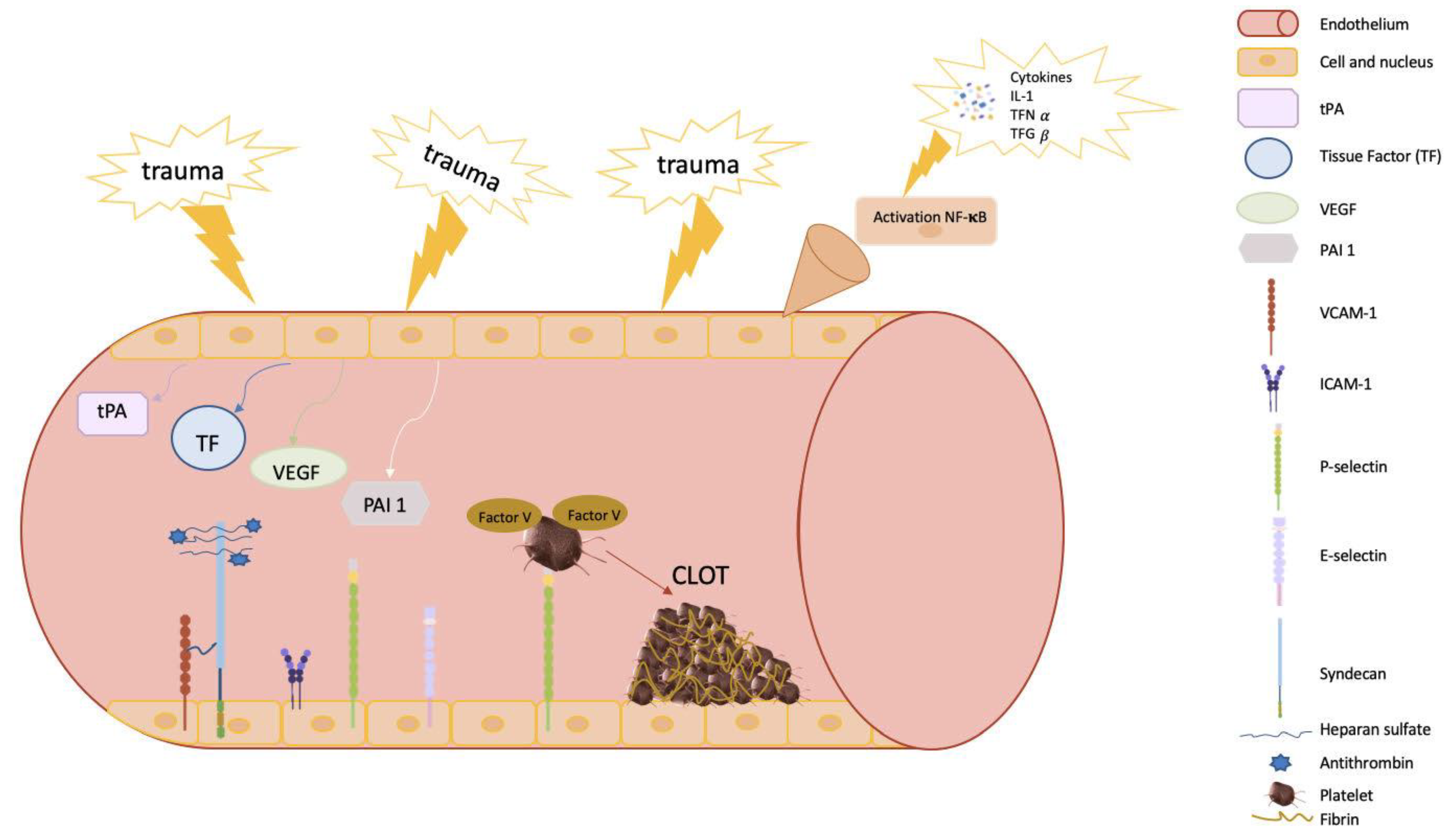
1.1. Key Mediators and Factors Regulating Coagulation
1.2. Non-Coding RNAs Involved in Disseminated Intravascular Coagulation
1.3. Key Components in the Coagulation Cascade
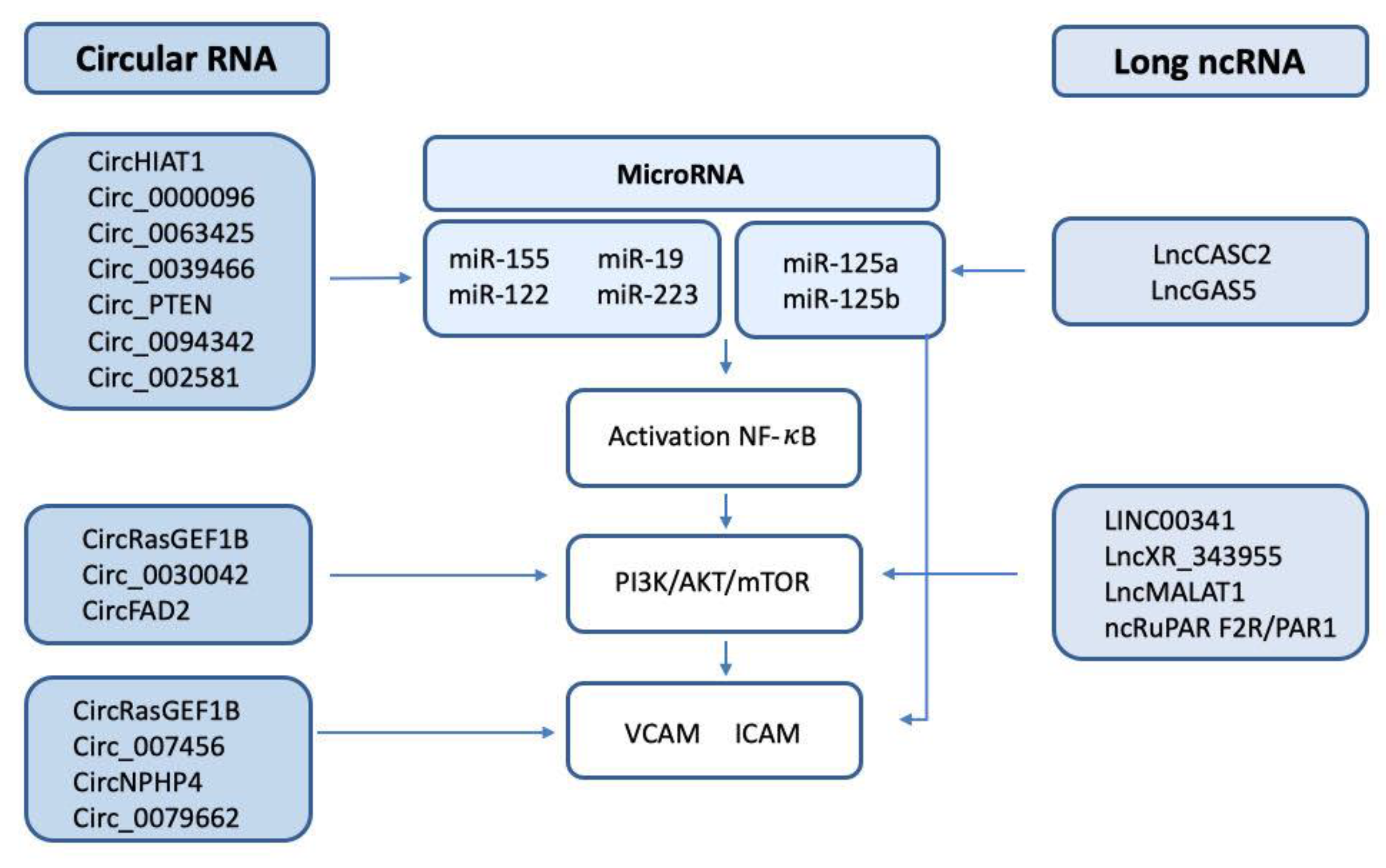
2. The Intricate Pathways Modulated by Non-Coding RNAs in DIC
2.1. The Role of microRNAs in DIC
2.2. The Role of Long Non-Coding RNAs in DIC
2.3. The Role of Circular RNAs in DIC
3. Discussion and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, Regional, and National Sepsis Incidence and Mortality, 1990–2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Ince, C.; Mayeux, P.R.; Nguyen, T.; Gomez, H.; Kellum, J.A.; Ospina-Tascón, G.A.; Hernandez, G.; Murray, P.; De Backer, D. The Endothelium In Sepsis. Shock 2016, 45, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Bermejo-Martin, J.F.; Martín-Fernandez, M.; López-Mestanza, C.; Duque, P.; Almansa, R. Shared Features of Endothelial Dysfunction between Sepsis and Its Preceding Risk Factors (Aging and Chronic Disease). J. Clin. Med. 2018, 7, 400. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, K.; Bayer, O.; Brunkhorst, F.; Meisner, M. Markers of Endothelial Damage in Organ Dysfunction and Sepsis. Crit. Care Med. 2002, 30, S302–S312. [Google Scholar] [CrossRef]
- Semeraro, N.; Colucci, M. Endothelial Cell Perturbation and Disseminated Intravascular Coagulation. In Molecular Mechanisms of Disseminated Intravascular Coagulation; Ten Cate, H., Levi, M., Eds.; Landes Bioscience: Georgetown, DC, USA, 2003; pp. 156–190. [Google Scholar]
- Semeraro, N.; Ammollo, C.T.; Semeraro, F.; Colucci, M. Sepsis, Thrombosis and Organ Dysfunction. Thromb. Res. 2012, 129, 290–295. [Google Scholar] [CrossRef]
- Taylor, F.; Toh, C.-H.; Hoots, K.; Wada, H.; Levi, M. Towards Definition, Clinical and Laboratory Criteria, and a Scoring System for Disseminated Intravascular Coagulation: On Behalf of the Scientific Subcommittee on Disseminated Intravascular Coagulation (DIC) of the International Society on Thrombosis and Haemostasis (ISTH). Thromb. Haemost. 2001, 86, 1327–1330. [Google Scholar] [CrossRef]
- Gando, S.; Levi, M.; Toh, C.-H. Disseminated Intravascular Coagulation. Nat. Rev. Dis. Primer 2016, 2, 16037. [Google Scholar] [CrossRef]
- Ikezoe, T. Thrombomodulin/Activated Protein C System in Septic Disseminated Intravascular Coagulation. J. Intensive Care 2015, 3, 1. [Google Scholar] [CrossRef]
- Gando, S.; Saitoh, D.; Ogura, H.; Mayumi, T.; Koseki, K.; Ikeda, T.; Ishikura, H.; Iba, T.; Ueyama, M.; Eguchi, Y.; et al. Natural History of Disseminated Intravascular Coagulation Diagnosed Based on the Newly Established Diagnostic Criteria for Critically Ill Patients: Results of a Multicenter, Prospective Survey. Crit. Care Med. 2008, 36, 145–150. [Google Scholar] [CrossRef]
- Vincent, J.-L.; Castro, P.; Hunt, B.J.; Jörres, A.; Praga, M.; Rojas-Suarez, J.; Watanabe, E. Thrombocytopenia in the ICU: Disseminated Intravascular Coagulation and Thrombotic Microangiopathies—What Intensivists Need to Know. Crit. Care 2018, 22, 158. [Google Scholar] [CrossRef]
- Oto, J.; Fernández-Pardo, Á.; Miralles, M.; Plana, E.; España, F.; Navarro, S.; Medina, P. Activated Protein C Assays: A Review. Clin. Chim. Acta 2020, 502, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Wada, H.; Matsumoto, T.; Yamashita, Y.; Hatada, T. Disseminated Intravascular Coagulation: Testing and Diagnosis. Clin. Chim. Acta 2014, 436, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Kuenen, B.C.; Levi, M.; Meijers, J.C.M.; Kakkar, A.K.; van Hinsbergh, V.W.M.; Kostense, P.J.; Pinedo, H.M.; Hoekman, K. Analysis of Coagulation Cascade and Endothelial Cell Activation During Inhibition of Vascular Endothelial Growth Factor/Vascular Endothelial Growth Factor Receptor Pathway in Cancer Patients. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1500–1505. [Google Scholar] [CrossRef]
- Pepper, M.S.; Ferrara, N.; Orci, L.; Montesano, R. Vascular Endothelial Growth Factor (VEGF) Induces Plasminogen Activators and Plasminogen Activator Inhibitor-1 in Microvascular Endothelial Cells. Biochem. Biophys. Res. Commun. 1991, 181, 902–906. [Google Scholar] [CrossRef]
- Mandriota, S.J.; Pepper, M.S. Vascular Endothelial Growth Factor-Induced in Vitro Angiogenesis and Plasminogen Activator Expression Are Dependent on Endogenous Basic Fibroblast Growth Factor. J. Cell Sci. 1997, 110, 2293–2302. [Google Scholar] [CrossRef]
- Kroon, M.E.; Koolwijk, P.; Vermeer, M.A.; van der Vecht, B.; van Hinsbergh, V.W.M. Vascular Endothelial Growth Factor Enhances the Expression of Urokinase Receptor in Human Endothelial Cells via Protein Kinase C Activation. Thromb. Haemost. 2001, 85, 296–302. [Google Scholar] [CrossRef]
- Chavda, V.; Madhwani, K. Coding and Non-Coding Nucleotides’: The Future of Stroke Gene Therapeutics. Genomics 2021, 113, 1291–1307. [Google Scholar] [CrossRef]
- Liu, N.; Wang, Z.-Z.; Zhao, M.; Zhang, Y.; Chen, N.-H. Role of Non-Coding RNA in the Pathogenesis of Depression. Gene 2020, 735, 144276. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hamblin, M.H.; Yin, K.-J. Noncoding RNAs and Stroke. Neuroscientist 2019, 25, 22–26. [Google Scholar] [CrossRef]
- Faraldi, M.; Gomarasca, M.; Banfi, G.; Lombardi, G. Free Circulating MiRNAs Measurement in Clinical Settings. Adv. Clin. Chem. 2018, 87, 113–139. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a Big Role in Gene Regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Willeit, P.; Zampetaki, A.; Dudek, K.; Kaudewitz, D.; King, A.; Kirkby, N.S.; Crosby-Nwaobi, R.; Prokopi, M.; Drozdov, I.; Langley, S.R.; et al. Circulating MicroRNAs as Novel Biomarkers for Platelet Activation. Circ. Res. 2013, 112, 595–600. [Google Scholar] [CrossRef]
- Salloum-Asfar, S.; Teruel-Montoya, R.; Arroyo, A.B.; García-Barberá, N.; Chaudhry, A.; Schuetz, E.; Luengo-Gil, G.; Vicente, V.; González-Conejero, R.; Martínez, C. Regulation of Coagulation Factor XI Expression by MicroRNAs in the Human Liver. PLoS ONE 2014, 9, e111713. [Google Scholar] [CrossRef]
- Morelli, V.M.; Brækkan, S.K.; Hansen, J.-B. Role of MicroRNAs in Venous Thromboembolism. Int. J. Mol. Sci. 2020, 21, 2602. [Google Scholar] [CrossRef]
- Fort, A.; Borel, C.; Migliavacca, E.; Antonarakis, S.E.; Fish, R.J.; Neerman-Arbez, M. Regulation of Fibrinogen Production by MicroRNAs. Blood 2010, 116, 2608–2615. [Google Scholar] [CrossRef]
- Teruel, R.; Pérez-Sánchez, C.; Corral, J.; Herranz, M.T.; Pérez-Andreu, V.; Saiz, E.; García-Barberá, N.; Martínez-Martínez, I.; Roldán, V.; Vicente, V.; et al. Identification of MiRNAs as Potential Modulators of Tissue Factor Expression in Patients with Systemic Lupus Erythematosus and Antiphospholipid Syndrome. J. Thromb. Haemost. 2011, 9, 1985–1992. [Google Scholar] [CrossRef]
- Bhatti, G.K.; Khullar, N.; Sidhu, I.S.; Navik, U.S.; Reddy, A.P.; Reddy, P.H.; Bhatti, J.S. Emerging Role of Non-coding RNA in Health and Disease. Metab. Brain Dis. 2021, 36, 1119–1134. [Google Scholar] [CrossRef]
- Morillon, A. 2-Definition and Families of Long Non-Coding RNA. In Long Non-coding RNA; Morillon, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 25–53. ISBN 978-1-78548-265-6. [Google Scholar]
- Nie, M.-W.; Han, Y.-C.; Shen, Z.-J.; Xie, H.-Z. Identification of CircRNA and MRNA Expression Profiles and Functional Networks of Vascular Tissue in Lipopolysaccharide-Induced Sepsis. J. Cell. Mol. Med. 2020, 24, 7915–7927. [Google Scholar] [CrossRef]
- Yu, Z.; Huang, Q.; Zhang, Q.; Wu, H.; Zhong, Z. CircRNAs Open a New Era in the Study of Cardiovascular Disease (Review). Int. J. Mol. Med. 2021, 47, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Rybak-Wolf, A.; Stottmeister, C.; Glažar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef]
- Huang, S.; Yang, B.; Chen, B.J.; Bliim, N.; Ueberham, U.; Arendt, T.; Janitz, M. The Emerging Role of Circular RNAs in Transcriptome Regulation. Genomics 2017, 109, 401–407. [Google Scholar] [CrossRef]
- Bolha, L.; Ravnik-Glavač, M.; Glavač, D. Circular RNAs: Biogenesis, Function, and a Role as Possible Cancer Biomarkers. Int. J. Genom. 2017, 2017, 6218353. [Google Scholar] [CrossRef] [PubMed]
- Dragomir, M.; Calin, G.A. Circular RNAs in Cancer-Lessons Learned From MicroRNAs. Front. Oncol. 2018, 8, 179. [Google Scholar] [CrossRef] [PubMed]
- Beltrán-García, J.; Osca-Verdegal, R.; Nacher-Sendra, E.; Pallardó, F.V.; García-Giménez, J.L. Circular RNAs in Sepsis: Biogenesis, Function, and Clinical Significance. Cells 2020, 9, 1544. [Google Scholar] [CrossRef] [PubMed]
- Hashemian, S.M.; Pourhanifeh, M.H.; Fadaei, S.; Velayati, A.A.; Mirzaei, H.; Hamblin, M.R. Non-Coding RNAs and Exosomes: Their Role in the Pathogenesis of Sepsis. Mol. Ther. Nucleic Acids 2020, 21, 51–74. [Google Scholar] [CrossRef]
- Panda, A.C. Circular RNAs Act as MiRNA Sponges. In Circular RNAs: Biogenesis and Functions; Xiao, J., Ed.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2018; pp. 67–79. ISBN 9789811314261. [Google Scholar]
- Huang, A.; Zheng, H.; Wu, Z.; Chen, M.; Huang, Y. Circular RNA-Protein Interactions: Functions, Mechanisms, and Identification. Theranostics 2020, 10, 3503–3517. [Google Scholar] [CrossRef]
- Memtsas, V.P.; Arachchillage, D.R.J.; Gorog, D.A. Role, Laboratory Assessment and Clinical Relevance of Fibrin, Factor XIII and Endogenous Fibrinolysis in Arterial and Venous Thrombosis. Int. J. Mol. Sci. 2021, 22, 1472. [Google Scholar] [CrossRef]
- Song, J.W.; Choi, J.R.; Song, K.S.; Rhee, J.H. Plasma Factor XIII Activity in Patients with Disseminated Intravascular Coagulation. Yonsei Med. J. 2006, 47, 196–200. [Google Scholar] [CrossRef]
- Gailani, D.; Broze, G.J. Factor XI Activation in a Revised Model of Blood Coagulation. Science 1991, 253, 909–912. [Google Scholar] [CrossRef]
- Lippi, G.; Favaloro, E.J.; Franchini, M.; Guidi, G.C. Milestones and Perspectives in Coagulation and Hemostasis. Semin. Thromb. Hemost. 2009, 35, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Levi, M.; Van Der Poll, T.; Ten Cate, H.; Van Deventer, S.J.H. The Cytokine-Mediated Imbalance between Coagulant and Anticoagulant Mechanisms in Sepsis and Endotoxaemia. Eur. J. Clin. Investig. 1997, 27, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Nightingale, T.; Cutler, D. The Secretion of von Willebrand Factor from Endothelial Cells; an Increasingly Complicated Story. J. Thromb. Haemost. 2013, 11, 192–201. [Google Scholar] [CrossRef]
- Federici, A.B. The Factor VIII/von Willebrand Factor Complex: Basic and Clinical Issues. Haematologica 2003, 88, EREP02. [Google Scholar] [CrossRef] [PubMed]
- Lam, F.W.; Cruz, M.A.; Leung, H.-C.E.; Parikh, K.S.; Smith, C.W.; Rumbaut, R.E. Histone Induced Platelet Aggregation Is Inhibited by Normal Albumin. Thromb. Res. 2013, 132, 69–76. [Google Scholar] [CrossRef]
- Joffre, J.; Hellman, J.; Ince, C.; Ait-Oufella, H. Endothelial Responses in Sepsis. Am. J. Respir. Crit. Care Med. 2020, 202, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Dolmatova, E.V.; Wang, K.; Mandavilli, R.; Griendling, K.K. The Effects of Sepsis on Endothelium and Clinical Implications. Cardiovasc. Res. 2020, 117, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Sano, H.; Nakagawa, N.; Nakajima, H.; Yoshida, S.; Iwamoto, I. Role of Vascular Cell Adhesion Molecule-1 and Platelet-Activating Factor in Selective Eosinophil Migration across Vascular Endothelial Cells. Int. Arch. Allergy Immunol. 1995, 107, 533–540. [Google Scholar] [CrossRef]
- Hack, C.E. Tissue Factor Pathway of Coagulation in Sepsis. Crit. Care Med. 2000, 28, S25–S30. [Google Scholar] [CrossRef]
- Sharma, S.; Tyagi, T.; Antoniak, S. Platelet in Thrombo-Inflammation: Unraveling New Therapeutic Targets. Front. Immunol. 2022, 13, 1039843. [Google Scholar] [CrossRef]
- Mammen, E.F. Antithrombin: Its Physiological Importance and Role in DIC. Semin. Thromb. Hemost. 1998, 24, 19–25. [Google Scholar] [CrossRef]
- Morello, F.; Perino, A.; Hirsch, E. Phosphoinositide 3-Kinase Signalling in the Vascular System. Cardiovasc. Res. 2009, 82, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Puri, K.D.; Doggett, T.A.; Huang, C.-Y.; Douangpanya, J.; Hayflick, J.S.; Turner, M.; Penninger, J.; Diacovo, T.G. The Role of Endothelial PI3Kγ Activity in Neutrophil Trafficking. Blood 2005, 106, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Puri, K.D.; Doggett, T.A.; Douangpanya, J.; Hou, Y.; Tino, W.T.; Wilson, T.; Graf, T.; Clayton, E.; Turner, M.; Hayflick, J.S.; et al. Mechanisms and Implications of Phosphoinositide 3-Kinase δ in Promoting Neutrophil Trafficking into Inflamed Tissue. Blood 2004, 103, 3448–3456. [Google Scholar] [CrossRef] [PubMed]
- Lazaar, A.L.; Krymskaya, V.P.; Das, S.K.P. VCAM-1 Activates Phosphatidylinositol 3-Kinase and Induces P120Cbl Phosphorylation in Human Airway Smooth Muscle Cells 1. J. Immunol. 2001, 166, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Tsoyi, K.; Jang, H.J.; Nizamutdinova, I.T.; Park, K.; Kim, Y.M.; Kim, H.J.; Seo, H.G.; Lee, J.H.; Chang, K.C. PTEN Differentially Regulates Expressions of ICAM-1 and VCAM-1 through PI3K/Akt/GSK-3β/GATA-6 Signaling Pathways in TNF-α-Activated Human Endothelial Cells. Atherosclerosis 2010, 213, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.Y.; Calin, G.A. The Mix of Two Worlds: Non-Coding RNAs and Hormones. Nucleic Acid Ther. 2013, 23, 2–8. [Google Scholar] [CrossRef]
- Wang, H.; Bei, Y.; Huang, P.; Zhou, Q.; Shi, J.; Sun, Q.; Zhong, J.; Li, X.; Kong, X.; Xiao, J. Inhibition of MiR-155 Protects Against LPS-Induced Cardiac Dysfunction and Apoptosis in Mice. Mol. Ther. Nucleic Acids 2016, 5, e374. [Google Scholar] [CrossRef]
- Etzrodt, V.; Idowu, T.O.; Schenk, H.; Seeliger, B.; Prasse, A.; Thamm, K.; Pape, T.; Müller-Deile, J.; van Meurs, M.; Thum, T.; et al. Role of Endothelial MicroRNA 155 on Capillary Leakage in Systemic Inflammation. Crit. Care 2021, 25, 76. [Google Scholar] [CrossRef]
- Ntanyane Phasha, M.-A.; Soma, P.; Van Rooy, M.; Phulukdaree, A. MicroRNA 155, Factor XIII and Type 2 Diabetes Mellitus and Coronary Heart Disease. Curr. Diabetes Rev. 2023, 19, e190822207740. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, P.; Chen, W.; Feng, D.; Jia, Y.; Xie, L. Serum MicroRNA Signatures Identified by Solexa Sequencing Predict Sepsis Patients’ Mortality: A Prospective Observational Study. PLoS ONE 2012, 7, e38885. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, P.; Chen, W.; Feng, D.; Jia, Y.; Xie, L. Four Serum MicroRNAs Identified as Diagnostic Biomarkers of Sepsis. J. Trauma Acute Care Surg. 2012, 73, 850. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-J.; Deng, J.; Wang, J.-Y.; Zhang, P.-J.; Xin, Z.; Xiao, K.; Feng, D.; Jia, Y.-H.; Liu, Y.-N.; Xie, L.-X. Serum MiR-122 Levels Are Related to Coagulation Disorders in Sepsis Patients. Clin. Chem. Lab. Med. CCLM 2014, 52, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Wang, X.; Ha, T.; Gao, M.; Liu, L.; Wang, R.; Yu, K.; Kalbfleisch, J.H.; Kao, R.L.; Williams, D.L.; et al. MicroRNA-125b Prevents Cardiac Dysfunction in Polymicrobial Sepsis by Targeting TRAF6-Mediated Nuclear Factor ΚB Activation and P53-Mediated Apoptotic Signaling. J. Infect. Dis. 2016, 214, 1773–1783. [Google Scholar] [CrossRef]
- Zhu, X. MiR-125b but Not MiR-125a Is Upregulated and Exhibits a Trend to Correlate with Enhanced Disease Severity, Inflammation, and Increased Mortality in Sepsis Patients. J. Clin. Lab. Anal. 2019, 34, e23094. [Google Scholar] [CrossRef]
- Nourse, J.; Danckwardt, S. A Novel Rationale for Targeting FXI: Insights from the Hemostatic MicroRNA Targetome for Emerging Anticoagulant Strategies. Pharmacol. Ther. 2021, 218, 107676. [Google Scholar] [CrossRef]
- Chen, L.-J.; Yang, L.; Cheng, X.; Xue, Y.-K.; Chen, L.-B. Overexpression of MiR-24 Is Involved in the Formation of Hypocoagulation State after Severe Trauma by Inhibiting the Synthesis of Coagulation Factor X. Dis. Markers 2017, 2017, 3649693. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Lu, S.; Yang, X.; Li, M.; Jia, H.; Liao, J.; Jing, Q.; Wu, Y.; Wang, H.; Xiao, F.; et al. MiR-19a-3p Downregulates Tissue Factor and Functions as a Potential Therapeutic Target for Sepsis-Induced Disseminated Intravascular Coagulation. Biochem. Pharmacol. 2021, 192, 114671. [Google Scholar] [CrossRef]
- Jiang, R.; Wang, N.-P.; Tanaka, K.A.; Levy, J.H.; Guyton, R.A.; Zhao, Z.-Q.; Vinten-Johansen, J. Factor Xa Induces Tissue Factor Expression in Endothelial Cells by P44/42 MAPK and NF-ΚB-Dependent Pathways. J. Surg. Res. 2011, 169, 319–327. [Google Scholar] [CrossRef]
- Schabbauer, G.; Tencati, M.; Pedersen, B.; Pawlinski, R.; Mackman, N. PI3K-Akt Pathway Suppresses Coagulation and Inflammation in Endotoxemic Mice. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1963–1969. [Google Scholar] [CrossRef] [PubMed]
- Laffont, B.; Corduan, A.; Plé, H.; Duchez, A.-C.; Cloutier, N.; Boilard, E.; Provost, P. Activated Platelets Can Deliver MRNA Regulatory Ago2•microRNA Complexes to Endothelial Cells via Microparticles. Blood 2013, 122, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, H.; Ren, J.; Geng, Q.; Song, J.; Lee, C.; Cao, C.; Zhang, J.; Xu, N. MicroRNA-223 Inhibits Tissue Factor Expression in Vascular Endothelial Cells. Atherosclerosis 2014, 237, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yu, H.; Lou, J.R.; Zheng, J.; Zhu, H.; Popescu, N.-I.; Lupu, F.; Lind, S.E.; Ding, W.-Q. MicroRNA-19 (MiR-19) Regulates Tissue Factor Expression in Breast Cancer Cells. J. Biol. Chem. 2011, 286, 1429–1435. [Google Scholar] [CrossRef]
- Li, J.; Tan, M.; Xiang, Q.; Zhou, Z.; Yan, H. Thrombin-Activated Platelet-Derived Exosomes Regulate Endothelial Cell Expression of ICAM-1 via MicroRNA-223 during the Thrombosis-Inflammation Response. Thromb. Res. 2017, 154, 96–105. [Google Scholar] [CrossRef]
- Dai, G.-H.; Ma, P.-Z.; Song, X.-B.; Liu, N.; Zhang, T.; Wu, B. MicroRNA-223-3p Inhibits the Angiogenesis of Ischemic Cardiac Microvascular Endothelial Cells via Affecting RPS6KB1/Hif-1a Signal Pathway. PLoS ONE 2014, 9, e108468. [Google Scholar] [CrossRef]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: LncRNA Localization and Function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef]
- Wang, C.; Liang, G.; Shen, J.; Kong, H.; Wu, D.; Huang, J.; Li, X. Long Non-Coding RNAs as Biomarkers and Therapeutic Targets in Sepsis. Front. Immunol. 2021, 12, 722004. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Zhang, D.; Li, Y. The Role of Long Non-Coding RNAs in Sepsis-Induced Cardiac Dysfunction. Front. Cardiovasc. Med. 2021, 8, 684348. [Google Scholar] [CrossRef]
- Shen, Z.-J.; Han, Y.-C.; Nie, M.-W.; Wang, Y.-N.; Xiang, R.-L.; Xie, H.-Z. Genome-Wide Identification of Altered RNA M6A Profiles in Vascular Tissue of Septic Rats. Aging 2021, 13, 21610–21627. [Google Scholar] [CrossRef]
- Huang, T.-S.; Wang, K.-C.; Quon, S.; Nguyen, P.; Chang, T.-Y.; Chen, Z.; Li, Y.-S.; Subramaniam, S.; Shyy, J.; Chien, S. LINC00341 Exerts an Anti-Inflammatory Effect on Endothelial Cells by Repressing VCAM1. Physiol. Genom. 2017, 49, 339–345. [Google Scholar] [CrossRef]
- Wang, R.; Zhao, J.; Wei, Q.; Wang, H.; Zhao, C.; Hu, C.; Han, Y.; Hui, Z.; Yang, L.; Dai, Q.; et al. Potential of Circulating LncRNA CASC2 as a Biomarker in Reflecting the Inflammatory Cytokines, Multi-organ Dysfunction, Disease Severity, and Mortality in Sepsis Patients. J. Clin. Lab. Anal. 2022, 36, e24569. [Google Scholar] [CrossRef]
- Yang, X.; Li, L.; Liu, J.; Lv, B.; Chen, F. Extracellular Histones Induce Tissue Factor Expression in Vascular Endothelial Cells via TLR and Activation of NF-ΚB and AP-1. Thromb. Res. 2016, 137, 211–218. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, B.; Chen, W. LncRNA GAS5 Relates to Th17 Cells and Serves as a Potential Biomarker for Sepsis Inflammation, Organ Dysfunctions and Mortality Risk. J. Clin. Lab. Anal. 2022, 36, e24309. [Google Scholar] [CrossRef]
- Gao, H.; Ma, H.; Gao, M.; Chen, A.; Zha, S.; Yan, J. Long Non-Coding RNA GAS5 Aggravates Myocardial Depression in Mice with Sepsis via the MicroRNA-449b/HMGB1 Axis and the NF-ΚB Signaling Pathway. Biosci. Rep. 2021, 41, BSR20201738. [Google Scholar] [CrossRef]
- Zheng, D.; Hou, Y.; Li, Y.; Bian, Y.; Khan, M.; Li, F.; Huang, L.; Qiao, C. Long Non-Coding RNA Gas5 Is Associated With Preeclampsia and Regulates Biological Behaviors of Trophoblast via MicroRNA-21. Front. Genet. 2020, 11, 188. [Google Scholar] [CrossRef]
- Yu, B.; Cui, R.; Lan, Y.; Zhang, J.; Liu, B. Long Non-Coding RNA H19 as a Diagnostic Marker in Peripheral Blood of Patients with Sepsis. Am. J. Transl. Res. 2021, 13, 2923–2930. [Google Scholar]
- Liu, W.; Geng, F.; Yu, L. Long Non-coding RNA MALAT1/MicroRNA 125a Axis Presents Excellent Value in Discriminating Sepsis Patients and Exhibits Positive Association with General Disease Severity, Organ Injury, Inflammation Level, and Mortality in Sepsis Patients. J. Clin. Lab. Anal. 2020, 34, e23222. [Google Scholar] [CrossRef]
- Liang, W.-J.; Zeng, X.-Y.; Jiang, S.-L.; Tan, H.-Y.; Yan, M.-Y.; Yang, H.-Z. Long Non-Coding RNA MALAT1 Sponges MiR-149 to Promote Inflammatory Responses of LPS-Induced Acute Lung Injury by Targeting MyD88. Cell Biol. Int. 2020, 44, 317–326. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, L.; Liu, Z.; Wang, Y.; Yang, J. Long Noncoding RNA NEAT 1 and Its Target MicroRNA-125a in Sepsis: Correlation with Acute Respiratory Distress Syndrome Risk, Biochemical Indexes, Disease Severity, and 28-day Mortality. J. Clin. Lab. Anal. 2020, 34, e23509. [Google Scholar] [CrossRef]
- Huang, Q.; Huang, C.; Luo, Y.; He, F.; Zhang, R. Circulating LncRNA NEAT1 Correlates with Increased Risk, Elevated Severity and Unfavorable Prognosis in Sepsis Patients. Am. J. Emerg. Med. 2018, 36, 1659–1663. [Google Scholar] [CrossRef]
- Xu, R.; Yu, J.; Song, S.; Sun, D.; Xiu, L.; Xu, J.; Zhao, J.; Liu, X.; Ji, Q.; Yue, X. Long Non-Coding RNA NcRuPAR Regulates Gastric Cancer Cell Proliferation and Apoptosis via Phosphoinositide 3-Kinase/Protein Kinase B Signaling. Int. J. Med. Sci. 2022, 19, 1835–1846. [Google Scholar] [CrossRef]
- Liu, L.; Yan, B.; Yang, Z.; Zhang, X.; Gu, Q.; Yue, X. NcRuPAR Inhibits Gastric Cancer Progression by Down-Regulating Protease-Activated Receptor-1. Tumor Biol. 2014, 35, 7821–7829. [Google Scholar] [CrossRef]
- Jiao, W.; Zhou, X.; Wu, J.; Zhang, X.; Ding, J. Potential of Long Non-coding RNA KCNQ1OT1 as a Biomarker Reflecting Systemic Inflammation, Multiple Organ Dysfunction, and Mortality Risk in Sepsis Patients. J. Clin. Lab. Anal. 2021, 35, e24047. [Google Scholar] [CrossRef]
- Luo, Y.; Fu, Y.; Tan, T.; Hu, J.; Li, F.; Liao, Z.; Peng, J. Screening of LncRNA-MiRNA-MRNA Coexpression Regulatory Networks Involved in Acute Traumatic Coagulation Dysfunction Based on CTD, GeneCards, and PharmGKB Databases. Oxid. Med. Cell. Longev. 2022, 2022, 7280312. [Google Scholar] [CrossRef]
- Du, J.; Chen, M.; Liu, J.; Hu, P.; Guan, H.; Jiao, X. LncRNA F11-AS1 Suppresses Liver Hepatocellular Carcinoma Progression by Competitively Binding with MiR-3146 to Regulate PTEN Expression. J. Cell. Biochem. 2019, 120, 18457–18464. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, T.; Xiao, J. Circular RNAs: Promising Biomarkers for Human Diseases. EBioMedicine 2018, 34, 267–274. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs Are a Large Class of Animal RNAs with Regulatory Potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Salzman, J. Circular RNA Expression: Its Potential Regulation and Function. Trends Genet. TIG 2016, 32, 309–316. [Google Scholar] [CrossRef]
- Beltrán-García, J.; Osca-Verdegal, R.; Nácher-Sendra, E.; Cardona-Monzonís, A.; Sanchis-Gomar, F.; Carbonell, N.; Pallardó, F.V.; Lavie, C.J.; García-Giménez, J.L. Role of Non-Coding RNAs as Biomarkers of Deleterious Cardiovascular Effects in Sepsis. Prog. Cardiovasc. Dis. 2021, 68, 70–77. [Google Scholar] [CrossRef]
- Han, B.; Chao, J.; Yao, H. Circular RNA and Its Mechanisms in Disease: From the Bench to the Clinic. Pharmacol. Ther. 2018, 187, 31–44. [Google Scholar] [CrossRef]
- Ng, W.L.; Marinov, G.K.; Liau, E.S.; Lam, Y.L.; Lim, Y.-Y.; Ea, C.-K. Inducible RasGEF1B Circular RNA Is a Positive Regulator of ICAM-1 in the TLR4/LPS Pathway. RNA Biol. 2016, 13, 861–871. [Google Scholar] [CrossRef]
- Shi, M.; Li, Z.-Y.; Zhang, L.-M.; Wu, X.-Y.; Xiang, S.-H.; Wang, Y.-G.; Zhang, Y.-Q. Hsa_circ_0007456 Regulates the Natural Killer Cell-Mediated Cytotoxicity toward Hepatocellular Carcinoma via the MiR-6852-3p/ICAM-1 Axis. Cell Death Dis. 2021, 12, 94. [Google Scholar] [CrossRef]
- Xiong, F.; Mao, R.; Zhang, L.; Zhao, R.; Tan, K.; Liu, C.; Xu, J.; Du, G.; Zhang, T. CircNPHP4 in Monocyte-Derived Small Extracellular Vesicles Controls Heterogeneous Adhesion in Coronary Heart Atherosclerotic Disease. Cell Death Dis. 2021, 12, 948. [Google Scholar] [CrossRef]
- Liu, S.; Gao, J.; Wang, S. HOXA9 Inhibitors Promote Microcirculation of Coronary Arteries in Rats via Downregulating E-Selectin/VCAM-1. Exp. Ther. Med. 2021, 22, 871. [Google Scholar] [CrossRef]
- Niu, F.; Liang, X.; Ni, J.; Xia, Z.; Jiang, L.; Wang, H.; Liu, H.; Shen, G.; Li, X. CircRNA CircFADS2 Is Under-Expressed in Sepsis and Protects Lung Cells from LPS-Induced Apoptosis by Downregulating MiR-133a. J. Inflamm. 2022, 19, 4. [Google Scholar] [CrossRef]
- Tacke, F.; Roderburg, C.; Benz, F.; Cardenas, D.V.; Luedde, M.; Hippe, H.-J.; Frey, N.; Vucur, M.; Gautheron, J.; Koch, A.; et al. Levels of Circulating MiR-133a Are Elevated in Sepsis and Predict Mortality in Critically Ill Patients. Crit. Care Med. 2014, 42, 1096. [Google Scholar] [CrossRef]
- Weyrich, A.S.; Denis, M.M.; Schwertz, H.; Tolley, N.D.; Foulks, J.; Spencer, E.; Kraiss, L.W.; Albertine, K.H.; McIntyre, T.M.; Zimmerman, G.A. MTOR-Dependent Synthesis of Bcl-3 Controls the Retraction of Fibrin Clots by Activated Human Platelets. Blood 2007, 109, 1975–1983. [Google Scholar] [CrossRef]
- Woulfe, D.S. Akt Signaling in Platelets and Thrombosis. Expert Rev. Hematol. 2010, 3, 81–91. [Google Scholar] [CrossRef]
- Yu, F.; Zhang, Y.; Wang, Z.; Gong, W.; Zhang, C. Hsa_circ_0030042 Regulates Abnormal Autophagy and Protects Atherosclerotic Plaque Stability by Targeting EIF4A3. Theranostics 2021, 11, 5404–5417. [Google Scholar] [CrossRef]
- Zeng, Y.; Du, W.W.; Wu, Y.; Yang, Z.; Awan, F.M.; Li, X.; Yang, W.; Zhang, C.; Yang, Q.; Yee, A.; et al. A Circular RNA Binds To and Activates AKT Phosphorylation and Nuclear Localization Reducing Apoptosis and Enhancing Cardiac Repair. Theranostics 2017, 7, 3842–3855. [Google Scholar] [CrossRef] [PubMed]
- Cong, Y.; Li, Q.; Zhang, X.; Chen, Y.; Yu, K. MTOR Promotes Tissue Factor Expression and Activity in EGFR-Mutant Cancer. Front. Oncol. 2020, 10, 1615. [Google Scholar] [CrossRef]
- Hu, J.; Wang, R.; Liu, Y.; Zhou, J.; Shen, K.; Dai, Y. Baicalein Represses Cervical Cancer Cell Growth, Cell Cycle Progression and Promotes Apoptosis via Blocking AKT/MTOR Pathway by the Regulation of CircHIAT1/MiR-19a-3p Axis. OncoTargets Ther. 2021, 14, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.-K.; Chu, X.; Wang, S.; Sun, Y.; Zhang, J.; Dong, J.; Yan, Y.-X. Identification of Circulating Hsa_circ_0063425 and Hsa_circ_0056891 as Novel Biomarkers for Detection of Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2021, 106, e2688–e2699. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Huang, L.; Gest, C.; Xi, X.; Janin, A.; Soria, C.; Li, H.; Lu, H. Opposite Regulation by PI3K/Akt and MAPK/ERK Pathways of Tissue Factor Expression, Cell-Associated Procoagulant Activity and Invasiveness in MDA-MB-231 Cells. J. Hematol. Oncol. 2012, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Tang, N.; Hadden, T.J.; Rishi, A.K. Akt, FoxO and Regulation of Apoptosis. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2011, 1813, 1978–1986. [Google Scholar] [CrossRef]
- Yuan, F.; Tang, Y.; Cao, M.; Ren, Y.; Li, Y.; Yang, G.; Ou, Q.; Tustumi, F.; Levi Sandri, G.B.; Raissi, D.; et al. Identification of the Hsa_circ_0039466/MiR-96-5p/FOXO1 Regulatory Network in Hepatocellular Carcinoma by Whole-Transcriptome Analysis. Ann. Transl. Med. 2022, 10, 769. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z.; Lu, J.; Zhang, H. Circular RNA Circ-PTEN Elevates PTEN Inhibiting the Proliferation of Non-Small Cell Lung Cancer Cells. Hum. Cell 2021, 34, 1174–1184. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Yang, H.; Bai, M.; Ning, T.; Deng, T.; Liu, R.; Fan, Q.; Zhu, K.; Li, J.; et al. Exosome-delivered CircRNA Promotes Glycolysis to Induce Chemoresistance through the MiR-122-PKM2 Axis in Colorectal Cancer. Mol. Oncol. 2020, 14, 539–555. [Google Scholar] [CrossRef]
- Zhao, X.; Tian, Z.; Liu, L. CircATP2B1 Promotes Aerobic Glycolysis in Gastric Cancer Cells Through Regulation of the MiR-326 Gene Cluster. Front. Oncol. 2021, 11, 628624. [Google Scholar] [CrossRef]
- Chen, C.; Deng, L.; Nie, D.-K.; Jia, F.; Fu, L.-S.; Wan, Z.-Q.; Lan, Q. Circular RNA Pleiotrophin Promotes Carcinogenesis in Glioma via Regulation of MicroRNA-122/SRY-Box Transcription Factor 6 Axis. Eur. J. Cancer Prev. 2020, 29, 165. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Gao, J.; Zheng, R.; Yu, M.; Ren, Y.; Yan, T.; Huang, Y.; Li, Y. Antagonizing CircRNA_002581–MiR-122–CPEB1 Axis Alleviates NASH through Restoring PTEN–AMPK–MTOR Pathway Regulated Autophagy. Cell Death Dis. 2020, 11, 123. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Feng, C.; Xiang, Z.; Chen, Y.; Li, Y. CircRNA Expression Pattern and CircRNA-MiRNA-MRNA Network in the Pathogenesis of Nonalcoholic Steatohepatitis. Oncotarget 2016, 7, 66455–66467. [Google Scholar] [CrossRef]
- Calderone, V.; Gallego, J.; Fernandez-Miranda, G.; Garcia-Pras, E.; Maillo, C.; Berzigotti, A.; Mejias, M.; Bava, F.-A.; Angulo-Urarte, A.; Graupera, M.; et al. Sequential Functions of CPEB1 and CPEB4 Regulate Pathologic Expression of Vascular Endothelial Growth Factor and Angiogenesis in Chronic Liver Disease. Gastroenterology 2016, 150, 982–997. [Google Scholar] [CrossRef]
- Garikipati, V.N.S.; Verma, S.K.; Cheng, Z.; Liang, D.; Truongcao, M.M.; Cimini, M.; Yue, Y.; Huang, G.; Wang, C.; Benedict, C.; et al. Circular RNA CircFndc3b Modulates Cardiac Repair after Myocardial Infarction via FUS/VEGF-A Axis. Nat. Commun. 2019, 10, 4317. [Google Scholar] [CrossRef]
- Dvorak, H.F.; Brown, L.F.; Detmar, M.; Dvorak, A.M. Vascular Permeability Factor/Vascular Endothelial Growth Factor, Microvascular Hyperpermeability, and Angiogenesis. Am. J. Pathol. 1995, 146, 1029–1039. [Google Scholar]
- Nakasaki, T.; Wada, H.; Shigemori, C.; Miki, C.; Gabazza, E.C.; Nobori, T.; Nakamura, S.; Shiku, H. Expression of Tissue Factor and Vascular Endothelial Growth Factor Is Associated with Angiogenesis in Colorectal Cancer. Am. J. Hematol. 2002, 69, 247–254. [Google Scholar] [CrossRef]
- Gadomska, G.; Ziołkowska, K.; Boinska, J.; Filipiak, J.; Rość, D. Activation of TF-Dependent Blood Coagulation Pathway and VEGF-A in Patients with Essential Thrombocythemia. Medicina 2019, 55, 54. [Google Scholar] [CrossRef]
- Clauss, M.; Gerlach, M.; Gerlach, H.; Brett, J.; Wang, F.; Familletti, P.C.; Pan, Y.C.; Olander, J.V.; Connolly, D.T.; Stern, D. Vascular Permeability Factor: A Tumor-Derived Polypeptide That Induces Endothelial Cell and Monocyte Procoagulant Activity, and Promotes Monocyte Migration. J. Exp. Med. 1990, 172, 1535–1545. [Google Scholar] [CrossRef]
- Mechtcheriakova, D.; Wlachos, A.; Holzmüller, H.; Binder, B.R.; Hofer, E. Vascular Endothelial Cell Growth Factor–Induced Tissue Factor Expression in Endothelial Cells Is Mediated by EGR-1. Blood 1999, 93, 3811–3823. [Google Scholar] [CrossRef]
- Camera, M.; Giesen, P.L.A.; Fallon, J.; Aufiero, B.M.; Taubman, M.; Tremoli, E.; Nemerson, Y. Cooperation Between VEGF and TNF-α Is Necessary for Exposure of Active Tissue Factor on the Surface of Human Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Bajzar, L.; Nesheim, M.; Morser, J.; Tracy, P.B. Both Cellular and Soluble Forms of Thrombomodulin Inhibit Fibrinolysis by Potentiating the Activation of Thrombin-Activable Fibrinolysis Inhibitor. J. Biol. Chem. 1998, 273, 2792–2798. [Google Scholar] [CrossRef]
- Wu, W.; Zou, J. Studies on the Role of CircRNAs in Osteoarthritis. BioMed Res. Int. 2021, 2021, 8231414. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Yin, R.; Wang, Y.; Yang, S.; Ma, A.; Pan, X.; Zhu, X. Comprehensive Analysis of Peripheral Exosomal CircRNAs in Large Artery Atherosclerotic Stroke. Front. Cell Dev. Biol. 2021, 9, 685741. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.B.; Aggerbeck, M.A.; Granfeldt, A.; Schmidt, M.; Hvas, A.; Adelborg, K. Disseminated Intravascular Coagulation Diagnosis: Positive Predictive Value of the ISTH Score in a Danish Population. Res. Pract. Thromb. Haemost. 2021, 5, e12636. [Google Scholar] [CrossRef]
- Edwards, E.; Geng, L.; Tan, J.; Onishko, H.; Donnelly, E.; Hallahan, D.E. Phosphatidylinositol 3-Kinase/Akt Signaling in the Response of Vascular Endothelium to Ionizing Radiation. Cancer Res. 2002, 62, 4671–4677. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cánovas-Cervera, I.; Nacher-Sendra, E.; Osca-Verdegal, R.; Dolz-Andrés, E.; Beltrán-García, J.; Rodríguez-Gimillo, M.; Ferrando-Sánchez, C.; Carbonell, N.; García-Giménez, J.L. The Intricate Role of Non-Coding RNAs in Sepsis-Associated Disseminated Intravascular Coagulation. Int. J. Mol. Sci. 2023, 24, 2582. https://doi.org/10.3390/ijms24032582
Cánovas-Cervera I, Nacher-Sendra E, Osca-Verdegal R, Dolz-Andrés E, Beltrán-García J, Rodríguez-Gimillo M, Ferrando-Sánchez C, Carbonell N, García-Giménez JL. The Intricate Role of Non-Coding RNAs in Sepsis-Associated Disseminated Intravascular Coagulation. International Journal of Molecular Sciences. 2023; 24(3):2582. https://doi.org/10.3390/ijms24032582
Chicago/Turabian StyleCánovas-Cervera, Irene, Elena Nacher-Sendra, Rebeca Osca-Verdegal, Enric Dolz-Andrés, Jesús Beltrán-García, María Rodríguez-Gimillo, Carolina Ferrando-Sánchez, Nieves Carbonell, and José Luis García-Giménez. 2023. "The Intricate Role of Non-Coding RNAs in Sepsis-Associated Disseminated Intravascular Coagulation" International Journal of Molecular Sciences 24, no. 3: 2582. https://doi.org/10.3390/ijms24032582
APA StyleCánovas-Cervera, I., Nacher-Sendra, E., Osca-Verdegal, R., Dolz-Andrés, E., Beltrán-García, J., Rodríguez-Gimillo, M., Ferrando-Sánchez, C., Carbonell, N., & García-Giménez, J. L. (2023). The Intricate Role of Non-Coding RNAs in Sepsis-Associated Disseminated Intravascular Coagulation. International Journal of Molecular Sciences, 24(3), 2582. https://doi.org/10.3390/ijms24032582