
Dissecting the Complexity of Skeletal-Malocclusion-Associated Phenotypes: Mouse for the Rescue

 , ,
, , 

Abstract
1. Introduction
2. Genetics of Skeletal Class III Malocclusion Phenotype
3. Genome-Wide Association Study (GWAS)
4. Epigenetics and Skeletal Class III Malocclusion
5. Mouse Model for Studying Mandibular Size and Prognathism
6. Quantitative Trait Loci Analysis in Mouse Model
7. Mouse Knockout Gene
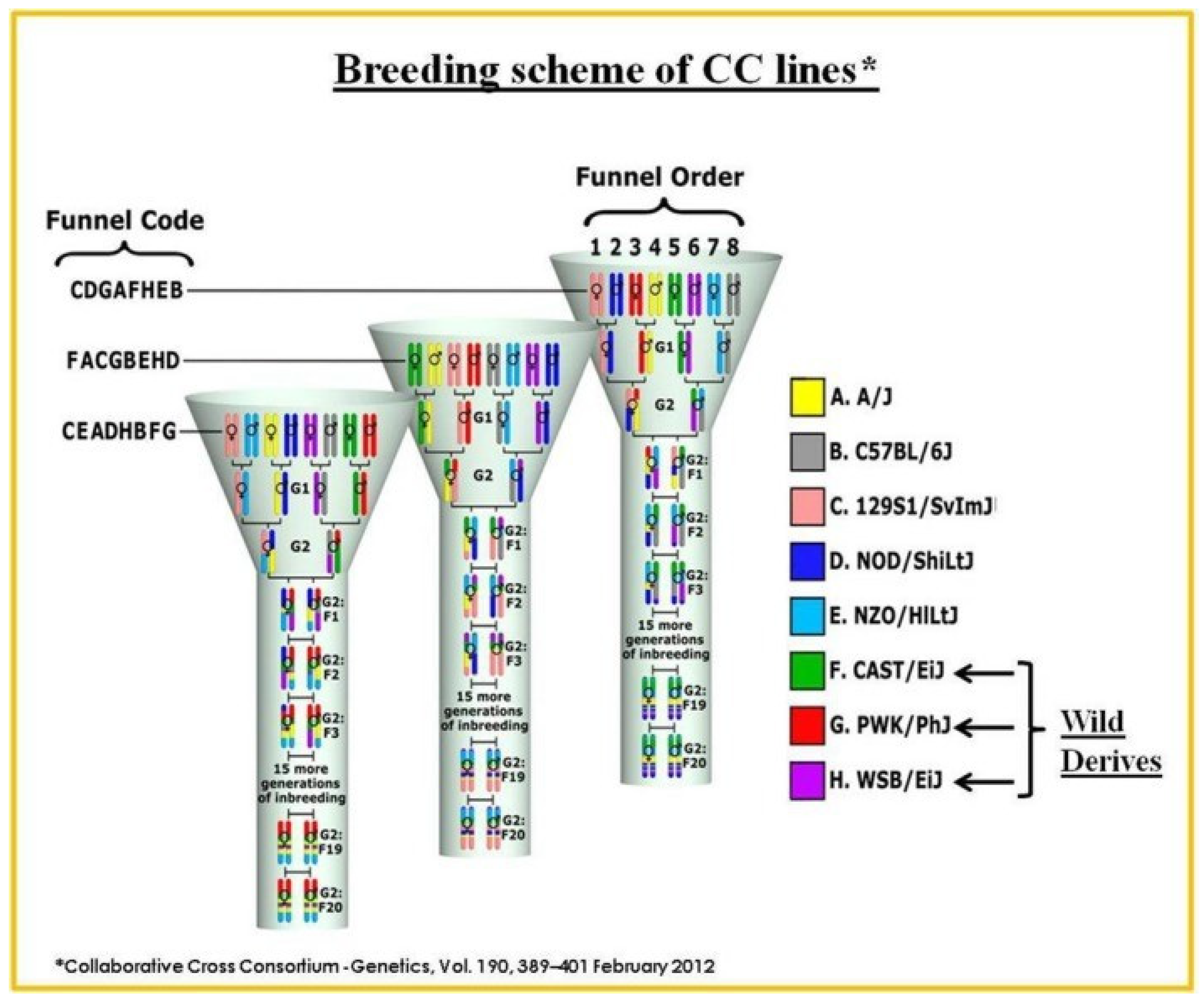
8. The Collaborative Cross (CC) Mouse Population as a Powerful Tool for Systematic Genetic Analysis of Class III Malocclusion
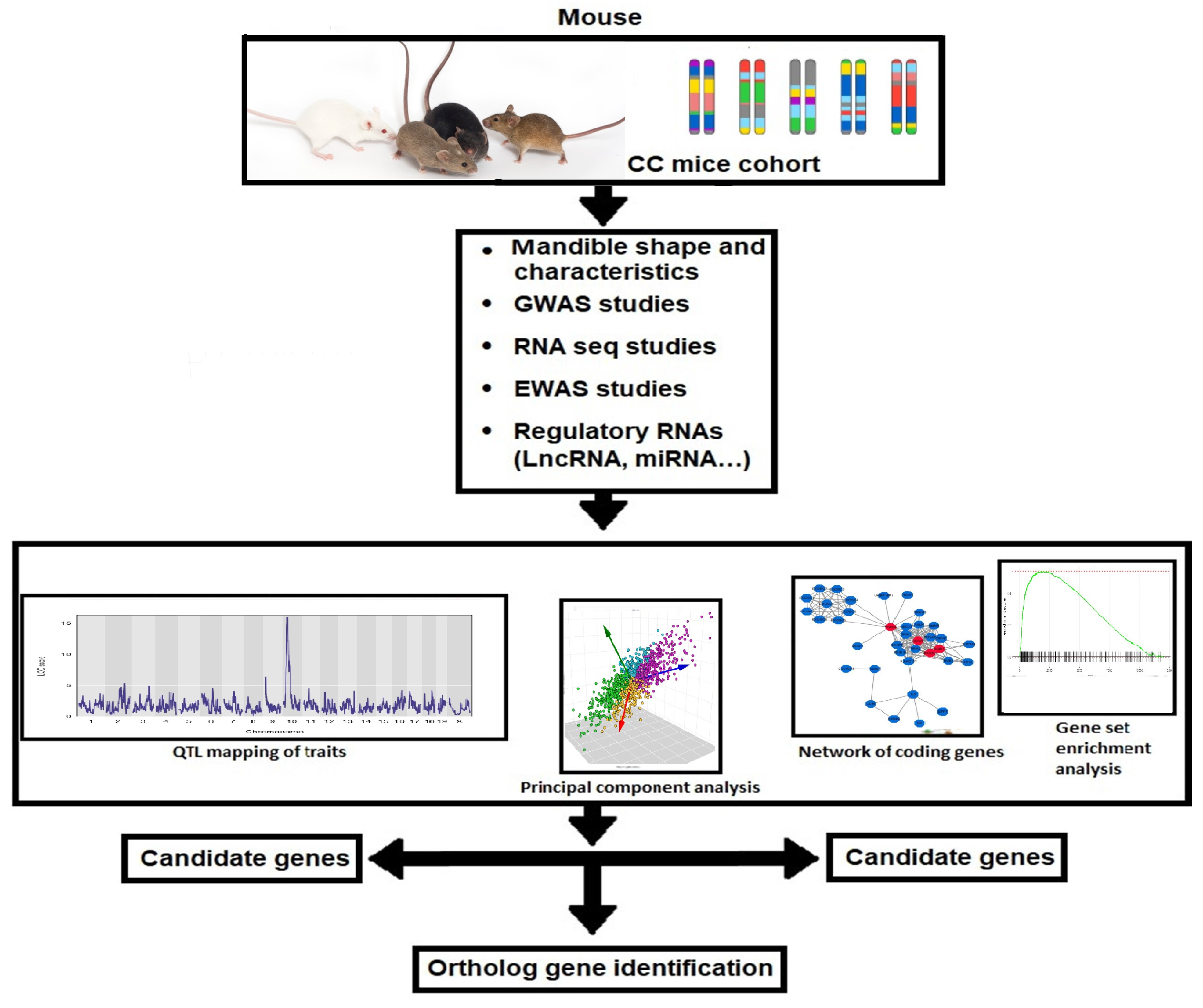
9. Future Direction for Comprehensively Dissecting the Complexity of Skeletal Class III Malocclusion Genes in CC Mice and in Humans
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Katz, D.C.; David Aponte, J.; Liu, W.; Green, R.M.; Mayeux, J.M.; Michael Pollard, K.; Pomp, D.; Munger, S.C.; Murray, S.A.; Roseman, C.C.; et al. Facial Shape and Allometry Quantitative Trait Locus Intervals in the Diversity Outbred Mouse Are Enriched for Known Skeletal and Facial Development Genes. PLoS ONE 2020, 15, e0233377. [Google Scholar] [CrossRef]
- Hu, D.; Marcucio, R.S. A SHH-Responsive Signaling Center in the Forebrain Regulates Craniofacial Morphogenesis via the Facial Ectoderm. Development 2009, 136, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Szabo-Rogers, H.L.; Smithers, L.E.; Yakob, W.; Liu, K.J. New Directions in Craniofacial Morphogenesis. Dev. Biol. 2010, 341, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Claudino, D.; Traebert, J. Malocclusion, Dental Aesthetic Self-Perception and Quality of Life in a 18 to 21 Year-Old Population: A Cross Section Study. BMC Oral Health 2013, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, R.R.; Nayme, J.G.; Garbin, A.J.; Saliba, N.; Garbin, C.A.; Moimaz, S.A. Prevalence of Malocclusion and Related Oral Habits in 5- to 6-Year-Old Children. Oral Health Prev. Dent. 2012, 10, 311–318. [Google Scholar] [CrossRef]
- Mtaya, M.; Brudvik, P.; Åstrøm, A.N. Prevalence of Malocclusion and Its Relationship with Socio-Demographic Factors, Dental Caries, and Oral Hygiene in 12- to 14-Year-Old Tanzanian Schoolchildren. Eur. J. Orthod. 2009, 31, 467–476. [Google Scholar] [CrossRef]
- Khan, M.; Fida, M. Assessment of Psychosocial Impact of Dental Aesthetics. J. Coll. Physicians Surg. Pak. 2008, 18, 559. [Google Scholar] [CrossRef]
- Gelgör, İ.E.; Karaman, İ.A.; Ercan, E. Prevalence of Malocclusion Among Adolescents in Central Anatolia. Eur. J. Dent. 2007, 01, 125–131. [Google Scholar] [CrossRef]
- Garbin, A.J.Í.; Perin, P.C.P.; Garbin, C.A.S.; Lolli, L.F. Malocclusion Prevalence and Comparison between the Angle Classification and the Dental Aesthetic Index in Scholars in the Interior of São Paulo State—Brazil. Dental Press J. Orthod. 2010, 15, 94–102. [Google Scholar] [CrossRef]
- Chan, G.K. hung Class III Malocclusion in Chinese (Cantonese): Etiology and Treatment. Am. J. Orthod. 1974, 65, 152–157. [Google Scholar] [CrossRef]
- Battagel, J.M. The Aetiological Factors in Class III Malocclusion. Eur. J. Orthod. 1993, 15, 347–370. [Google Scholar] [CrossRef] [PubMed]
- Lew, K.K.; Foong, W.C. Horizontal Skeletal Typing in an Ethnic Chinese Population with True Class III Malocclusions. Br. J. Orthod. 1993, 20, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Saleh, F.K. Prevalence of Malocclusion in a Sample of Lebanese Schoolchildren: An Epidemiological Study. East. Mediterr. Health J. 1999, 5, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Mouakeh, M. Cephalometric Evaluation of Craniofacial Pattern of Syrian Children with Class III Malocclusion. Am. J. Orthod. Dentofac. Orthop. 2001, 119, 640–649. [Google Scholar] [CrossRef] [PubMed]
- Toms, A.P. Class III Malocclusion: A Cephalometric Study of Saudi Arabians. Br. J. Orthod. 1989, 16, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Baccetti, T.; McGill, J.S.; Franchi, L.; McNamara, J.A.; Tollaro, I. Skeletal Effects of Early Treatment of Class III Malocclusion with Maxillary Expansion and Face-Mask Therapy. Am. J. Orthod. Dentofac. Orthop. 1998, 113, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Ellis, E.; McNamara, J.A. Components of Adult Class III Malocclusion. J. Oral Maxillofac. Surg. 1984, 42, 295–305. [Google Scholar] [CrossRef]
- Fatani, N.H.; Hammam, M.B.; Oraif, H.; Taher, S.; Taju, W.; Bukhari, O. Prevalence of Malocclusion among Schoolchildren in Makkah, Saudi Arabia. Open Access Maced. J. Med. Sci. 2019, 7, 856–861. [Google Scholar] [CrossRef]
- Cruz, R.M.; Krieger, H.; Ferreira, R.; Mah, J.; Hartsfield, J.; Oliveira, S. Major Gene and Multifactorial Inheritance of Mandibular Prognathism. Am. J. Med. Genet. A 2008, 146, 71–77. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Park, S.B.; Narita, A.; Maki, K.; Inoue, I. Genome-Wide Linkage Analysis of Mandibular Prognathism in Korean and Japanese Patients. J. Dent. Res. 2005, 84, 255–259. [Google Scholar] [CrossRef]
- Frazier-Bowers, S.; Rincon-Rodriguez, R.; Zhou, J.; Alexander, K.; Lange, E. Evidence of Linkage in a Hispanic Cohort with a Class III Dentofacial Phenotype. J. Dent. Res. 2009, 88, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Park, E.K.; Ryoo, H.M.; Shin, H.I.; Kim, T.H.; Jang, J.S.; Park, H.S.; Choi, J.Y.; Kwon, T.G. Polymorphisms in the Matrilin-1 Gene and Risk of Mandibular Prognathism in Koreans. J. Dent. Res. 2010, 89, 1203–1207. [Google Scholar] [CrossRef] [PubMed]
- Coussens, A.K.; van Daal, A. Linkage Disequilibrium Analysis Identifies an FGFR1 Haplotype-Tag SNP Associated with Normal Variation in Craniofacial Shape. Genomics 2005, 85, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lu, Y.; Gao, X.H.; Chen, Y.C.; Lu, J.J.; Bai, Y.X.; Shen, Y.; Wang, B.K. The Growth Hormone Receptor Gene Is Associated with Mandibular Height in a Chinese Population. J. Dent. Res. 2005, 84, 1052–1056. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Satoh, K.; Hayasaki, H.; Fukumoto, S.; Fujiwara, T.; Nonaka, K. The P561T Polymorphism of the Growth Hormone Receptor Gene Has an Inhibitory Effect on Mandibular Growth in Young Children. Eur. J. Orthod. 2009, 31, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.H.; Yamaguchi, T.; Tajima, A.; Nakajima, T.; Tomoyasu, Y.; Watanabe, M.; Yamaguchi, M.; Park, S.B.; Maki, K.; Inoue, I. Association of the Growth Hormone Receptor Gene Polymorphisms with Mandibular Height in a Korean Population. Arch. Oral Biol. 2009, 54, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Tomoyasu, Y.; Yamaguchi, T.; Tajima, A.; Nakajima, T.; Inoue, I.; Maki, K. Further Evidence for an Association between Mandibular Height and the Growth Hormone Receptor Gene in a Japanese Population. Am. J. Orthod. Dentofac. Orthop. 2009, 136, 536–541. [Google Scholar] [CrossRef]
- Li, Q.; Li, X.; Zhang, F.; Chen, F. The Identification of a Novel Locus for Mandibular Prognathism in the Han Chinese Population. J. Dent. Res. 2011, 90, 53–57. [Google Scholar] [CrossRef]
- Tassopoulou-Fishell, M.; Deeley, K.; Harvey, E.M.; Sciote, J.; Vieira, A.R. Genetic Variation in Myosin 1H Contributes to Mandibular Prognathism. Am. J. Orthod. Dentofac. Orthop. 2012, 141, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, S.J.; Gómez, M.; Rey, J.A.; Ochoa, M.; Gutiérrez, S.M.; Prieto, J.C. Polymorphisms of the Noggin Gene and Mandibular Micrognathia: A First Approximation. Acta Odontol. Latinoam. 2010, 23, 13–19. [Google Scholar]
- Brook, A.H. Multilevel Complex Interactions between Genetic, Epigenetic and Environmental Factors in the Aetiology of Anomalies of Dental Development. Arch. Oral Biol. 2009, 54, S3–S17. [Google Scholar] [CrossRef] [PubMed]
- Zabrina, S.; Ramadhanti, J.F.; Gultom, F.P.; Auerkari, E.I. Genetic and Epigenetic Aspects of Class III Malocclusion with Mandibular Prognathism Phenotypes. In Proceedings of the AIP Conference Proceedings; AIP Publishing LLC: Woodbury, NY, USA, 2021; Volume 2344. [Google Scholar]
- Desh, H.; Gray, S.L.; Horton, M.J.; Raoul, G.; Rowlerson, A.M.; Ferri, J.; Vieira, A.R.; Sciote, J.J. Molecular Motor MYO1C, Acetyltransferase KAT6B and Osteogenetic Transcription Factor RUNX2 Expression in Human Masseter Muscle Contributes to Development of Malocclusion. Arch. Oral Biol. 2014, 59, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Graber, T.M.; Vanarsdall, R.L.; Vig, K. Orthodontics: Current Principles & Techniques, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Huh, A.; Horton, M.J.; Cuenco, K.T.; Raoul, G.; Rowlerson, A.M.; Ferri, J.; Sciote, J.J. Epigenetic Influence of KAT6B and HDAC4 in the Development of Skeletal Malocclusion. Am. J. Orthod. Dentofac. Orthop. 2013, 144, 568–576. [Google Scholar] [CrossRef]
- Dohmoto, A.; Shimizu, K.; Asada, Y.; Maeda, T. Quantitative Trait Loci on Chromosomes 10 and 11 Influencing Mandible Size of SMXA RI Mouse Strains. J. Dent. Res. 2002, 81, 501–504. [Google Scholar] [CrossRef]
- Festing, M. Mouse Strain Identification. Nature 1972, 238, 351–352. [Google Scholar] [CrossRef]
- Goto, N.; Yamaoka, A.; Sudo, T.; Mannen, H.; Fukuta, K.; Nishimura, M. Morphometric Profiles of the Mandible of SMXA Recombinant Inbred Strains of Mice and Strain Identification on the Basis of Mandible Measurements. Jikken Dobutsu 1993, 42, 41–50. [Google Scholar] [CrossRef][Green Version]
- Klingenberg, C.P.; Leamy, L.J.; Routman, E.J.; Cheverud, J.M. Genetic Architecture of Mandible Shape in Mice: Effects of Quantitative Trait Loci Analyzed by Geometric Morphometrics. Genetics 2001, 157, 785–802. [Google Scholar] [CrossRef] [PubMed]
- Francis-West, P.; Ladher, R.; Barlow, A.; Graveson, A. Signalling Interactions during Facial Development. Mech. Dev. 1998, 75, 3–28. [Google Scholar] [CrossRef]
- Nadeau, J.H.; Frankel, W.N. The Roads from Phenotypic Variation to Gene Discovery: Mutagenesis versus QTLs. Nat. Genet. 2000, 25, 381–384. [Google Scholar] [CrossRef]
- Cheverud, J.M.; Routman, E.J.; Duarte, F.A.M.; van Swinderen, B.; Cothran, K.; Perel, C. Quantitative Trait Loci for Murine Growth. Genetics 1996, 142, 1305–1319. [Google Scholar] [CrossRef]
- Liu, X.; Oliver, F.; Brown, S.D.M.; Denny, P.; Keightley, P.D. High-Resolution Quantitative Trait Locus Mapping for Body Weight in Mice by Recombinant Progeny Testing. Genet. Res. 2001, 77, 191–197. [Google Scholar] [CrossRef]
- Acampora, D.; Mazan, S.; Avantaggiato, V.; Barone, P.; Tuorto, F.; Lallemand, Y.; Brulet, P.; Simeone, A. Epilepsy and Brain Abnormalities in Mice Lacking the Otx1 Gene. Nat. Genet. 1996, 14, 218–222. [Google Scholar] [CrossRef]
- Acampora, D.; Avantaggiato, V.; Tuorto, F.; Briata, P.; Corte, G.; Simeone, A. Visceral Endoderm-Restricted Translation of Otx1 Mediates Recovery of Otx2 Requirements for Specification of Anterior Neural Plate and Normal Gastrulation. Development 1998, 125, 5091–5104. [Google Scholar] [CrossRef] [PubMed]
- Leamy, L.J.; Pomp, D.; Eisen, E.J.; Cheverud, J.M. Quantitative Trait Loci for Directional but Not Fluctuating Asymmetry of Mandible Characters in Mice. Genet. Res. 2000, 76, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Da Fontoura, C.S.G.; Miller, S.F.; Wehby, G.L.; Amendt, B.A.; Holton, N.E.; Southard, T.E.; Allareddy, V.; Moreno Uribe, L.M. Candidate Gene Analyses of Skeletal Variation in Malocclusion. J. Dent. Res. 2015, 94, 913–920. [Google Scholar] [CrossRef]
- Nieto, M.A. The Snail Superfamily of Zinc-Finger Transcription Factors. Nat. Rev. Mol. Cell. Biol. 2002, 3, 155–166. [Google Scholar] [CrossRef]
- Pioli, P.D.; Dahlem, T.J.; Weis, J.J.; Weis, J.H. Deletion of Snai2 and Snai3 Results in Impaired Physical Development Compounded by Lymphocyte Deficiency. PLoS ONE 2013, 8, e69216. [Google Scholar] [CrossRef]
- Murray, S.A.; Oram, K.F.; Gridley, T. Multiple Functions of Snail Family Genes during Palate Development in Mice. Development 2007, 134, 1789–1797. [Google Scholar] [CrossRef]
- Leptin, M. Twist and Snail as Positive and Negative Regulators during Drosophila Mesoderm Development. Genes Dev. 1991, 5, 1568–1576. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.; Horsley, S.W.; Moloney, D.M.; Oldridge, M.; Twigg, S.R.F.; Walsh, S.; Barrow, M.; Njølstad, P.R.; Kunz, J.; Ashworth, G.J.; et al. A Comprehensive Screen for TWIST Mutations in Patients with Craniosynostosis Identifies a New Microdeletion Syndrome of Chromosome Band 7p21.1. Am. J. Hum. Genet. 1998, 63, 1282–1293. [Google Scholar] [CrossRef] [PubMed]
- Goos, J.A.C.; Mathijssen, I.M.J. Genetic Causes of Craniosynostosis: An Update. Mol. Syndromol. 2019, 10, 6–23. [Google Scholar] [CrossRef] [PubMed]
- Carver, E.A.; Oram, K.F.; Gridley, T. Craniosynostosis in Twist Heterozygous Mice: A Model for Saethre-Chotzen Syndrome. Anat. Rec. 2002, 268, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Blackwell, E.L.; Mcknight, M.T.; Knutsen, G.R.; Vu, W.T.; Ruest, L.B. Specific Inactivation of Twist1 in the Mandibular Arch Neural Crest Cells Affects the Development of the Ramus and Reveals Interactions with Hand2. Dev. Dyn. 2012, 241, 924–940. [Google Scholar] [CrossRef]
- Oram, K.F.; Gridley, T. Mutations in Snail Family Genes Enhance Craniosynostosis of Twist1 Haplo-Insufficient Mice: Implications for Saethre-Chotzen Syndrome. Genetics 2005, 170, 971–974. [Google Scholar] [CrossRef]
- Durrant, C.; Tayem, H.; Yalcin, B.; Cleak, J.; Goodstadt, L.; Pardo-Manuel De Villena, F.; Mott, R.; Iraqi, F.A. Collaborative Cross Mice and Their Power to Map Host Susceptibility to Aspergillus Fumigatus Infection. Genome Res. 2011, 21, 239–1248. [Google Scholar] [CrossRef] [PubMed]
- Iraqi, F.A.; Mahajne, M.; Salaymah, Y.; Sandovski, H.; Tayem, H.; Vered, K.; Balmer, L.; Hall, M.; Manship, G.; Morahan, G.; et al. The Genome Architecture of the Collaborative Cross Mouse Genetic Reference Population. Genetics 2012, 190, 389–401. [Google Scholar] [CrossRef]
- Atamni, H.J.A.T.; Mott, R.; Soller, M.; Iraqi, F.A. High-Fat-Diet Induced Development of Increased Fasting Glucose Levels and Impaired Response to Intraperitoneal Glucose Challenge in the Collaborative Cross Mouse Genetic Reference Population. BMC Genet. 2016, 17, 10. [Google Scholar] [CrossRef] [PubMed]
- Abu-Toamih Atamni, H.J.; Ziner, Y.; Mott, R.; Wolf, L.; Iraqi, F.A. Glucose Tolerance Female-Specific QTL Mapped in Collaborative Cross Mice. Mamm. Genome 2017, 28, 20–30. [Google Scholar] [CrossRef]
- Atamni, H.J.A.T.; Iraqi, F.A. Collaborative Cross as the Next-Generation Mouse Genetic Reference Population Designed for Dissecting Complex Traits. In Molecular-Genetic and Statistical Techniques for Behavioral and Neural Research; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Threadgill, D.W.; Hunter, K.W.; Williams, R.W. Genetic Dissection of Complex and Quantitative Traits: From Fantasy to Reality via a Community Effort. Mamm. Genome 2002, 13, 175–178. [Google Scholar] [CrossRef]
- Nashef, A.; Qahaz, N.; El-Naaj, I.A.; Iraqi, F.A. Systems Genetics Analysis of Oral Squamous Cell Carcinoma Susceptibility Using the Mouse Model: Current Position and New Perspective. Mamm. Genome 2021, 32, 323–331. [Google Scholar] [CrossRef]
- Nashef, A.; Matthias, M.; Weiss, E.; Loos, B.G.; Jepsen, S.; van der Velde, N.; Uitterlinden, A.G.; Wellmann, J.; Berger, K.; Hoffmann, P.; et al. Translation of Mouse Model to Human Gives Insights into Periodontitis Etiology. Sci. Rep. 2020, 10, 4892. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, L.; Xue, C.; Jun Li, M.; Li, M. A Conditional Gene-Based Association Framework Integrating Isoform-Level EQTL Data Reveals New Susceptibility Genes for Schizophrenia. Elife 2022, 11, e70779. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Chr | QTL | C.I. (cM) | Reference |
|---|---|---|---|
| 1 | QTL-SH1.1 | 38–62 | [39] |
| QTL-SH1.2 | 78–120 | [39] | |
| QTL-M1.1 | 12–38 | [46] | |
| QTL-M1.2 | 49–65 | [46] | |
| PC1 | 52–53 | [1] | |
| 2 | QTL-SH2.1 | — | [39] |
| QTL-C2.1 | — | [39] | |
| QTL-M2.1 | 21–40 | [46] | |
| QTL-M2.2 | 62–70 | [46] | |
| 3 | QTL-SH3.1 | 37–53 | [39] |
| QTL-C3.1 | 27–61 | [39] | |
| QTL-M3.1 | 14–24 | [46] | |
| QTL-M3.2 | 69–75 | [46] | |
| 4 | QTL-SH4.1 | 28–46 | [39] |
| QTL-C4.1 | 18–42 | [39] | |
| QTL-M4.1 | 40–60 | [46] | |
| 5 | QTL-SH5.1 | 37–105 | [39] |
| QTL-CU5.1 | 26–62 | [39] | |
| QTL-M5.1 | 29–70 | [46] | |
| 6 | QTL-SH6.1 | 4–16 | [39] |
| QTL-SH6.2 | 74–98 | [39] | |
| QTL-C6.1 | 70–100 | [39] | |
| QTL-M6.1 | 3–19 | [46] | |
| PC1 | 99–100.2 | [1] | |
| PC2 | 3.5–6 | [1] | |
| 7 | QTL-SH7.1 | 13–65 | [39] |
| QTL-SH7.2 | 61–77 | [39] | |
| QTL-C7.1 | 1–85 | [39] | |
| QTL-M7.1 | 37–75 | [46] | |
| PC1 | 58–66 | [1] | |
| 8 | QTL-M8.1 | 16–36 | [46] |
| 9 | QTL-SH9.1 | 16–38 | [39] |
| QTL-SH9.2 | 54–88 | [39] | |
| QTL-M9.1 | 41–65 | [46] | |
| 10 | QTL-SH10.1 | 9–41 | [39] |
| QTL-SH10.2 | 65–75 | [39] | |
| QTL-C10.1 | 63–87 | [39] | |
| QTL-CS10.1 | 55–83 | [39] | |
| QTL-M10.1 | 16–28 | [46] | |
| QTL-M10.2 | 31–57 | [46] | |
| 11 | QTL-SH11.1 | 13–31 | [39] |
| QTL-SH11.2 | 63–97 | [39] | |
| QTL-C11.1 | 17–55 | [39] | |
| QTL-C11.2 | 67–109 | [39] | |
| QTL-CS11.1 | 51–109 | [39] | |
| QTL-M11.1 | 4–47 | [46] | |
| 12 | QTL-SH12.1 | 21–33 | [39] |
| QTL-SH12.2 | 29–70 | [39] | |
| QTL-SU12.1 | 43–63 | [39] | |
| QTL-M12.1 | 32–49 | [46] | |
| PC3 | 8.5–17 | [1] | |
| 13 | QTL-SH13.1 | 79–97 | [39] |
| QTL-C13.1 | 7–45 | [39] | |
| QTL-M13.1 | 37–59 | [46] | |
| 14 | QTL-SH14.1 | 48–64 | [39] |
| QTL-C14.1 | 24–64 | [39] | |
| QTL-M14.1 | 5–27 | [46] | |
| QTL-M14.2 | 38–48 | [46] | |
| PC2 | 22–24 | [1] | |
| 15 | QTL-SH15.1 | 29–53 | [39] |
| QTL-SH15.2 | 53–85 | [39] | |
| QTL-C15.1 | 41–67 | [39] | |
| QTL-SS15.1 | 11–55 | [39] | |
| QTL-M15.1 | 27–43 | [46] | |
| 16 | QTL-SH16.1 | 14–44 | [39] |
| QTL-M16.1 | 15–33 | [46] | |
| 17 | QTL-C17.1 | 11–19 | [39] |
| QTL-M17.1 | 29–45 | [46] | |
| PC4 | 45–48 | [1] | |
| 18 | QTL-SH18.1 | 61–91 | [39] |
| QTL-M18.1 | 25–73 | [46] | |
| 19 | QTL-SH19.1 | 15–61 | [39] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lone, I.M.; Zohud, O.; Nashef, A.; Kirschneck, C.; Proff, P.; Watted, N.; Iraqi, F.A. Dissecting the Complexity of Skeletal-Malocclusion-Associated Phenotypes: Mouse for the Rescue. Int. J. Mol. Sci. 2023, 24, 2570. https://doi.org/10.3390/ijms24032570
Lone IM, Zohud O, Nashef A, Kirschneck C, Proff P, Watted N, Iraqi FA. Dissecting the Complexity of Skeletal-Malocclusion-Associated Phenotypes: Mouse for the Rescue. International Journal of Molecular Sciences. 2023; 24(3):2570. https://doi.org/10.3390/ijms24032570
Chicago/Turabian StyleLone, Iqbal M., Osayd Zohud, Aysar Nashef, Christian Kirschneck, Peter Proff, Nezar Watted, and Fuad A. Iraqi. 2023. "Dissecting the Complexity of Skeletal-Malocclusion-Associated Phenotypes: Mouse for the Rescue" International Journal of Molecular Sciences 24, no. 3: 2570. https://doi.org/10.3390/ijms24032570
APA StyleLone, I. M., Zohud, O., Nashef, A., Kirschneck, C., Proff, P., Watted, N., & Iraqi, F. A. (2023). Dissecting the Complexity of Skeletal-Malocclusion-Associated Phenotypes: Mouse for the Rescue. International Journal of Molecular Sciences, 24(3), 2570. https://doi.org/10.3390/ijms24032570