
Potato Stu-miR398b-3p Negatively Regulates Cu/Zn-SOD Response to Drought Tolerance

Abstract
:1. Introduction
2. Results
2.1. Secondary Structure Analysis of Stu-miR398b-3p Precursor Sequence
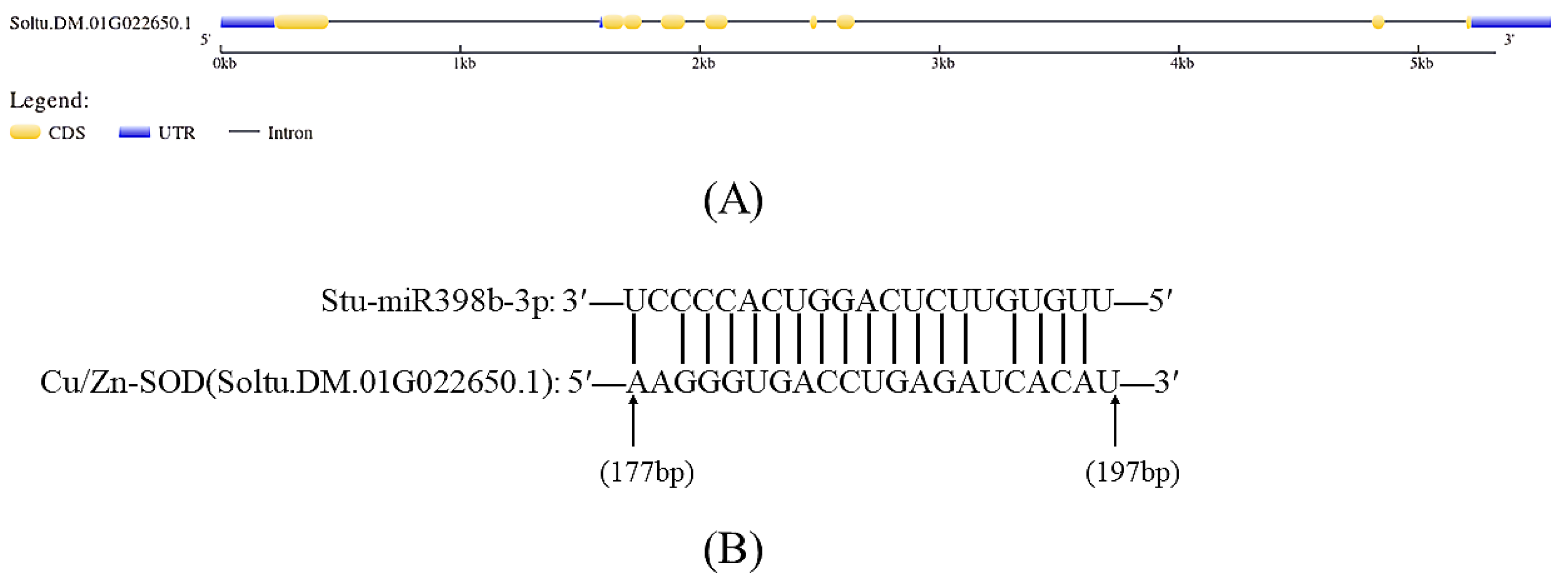
2.2. Prediction and Validation of Stu-miR398b-3p Target Genes
2.3. Bioinformatics Analysis of Stu-miR398b-3p Target Genes
2.4. Stu-miR398b-3p Construction of a miRNA Expression Vector
2.5. Potato Genetic Transformation and Identification of Transgenic Plants
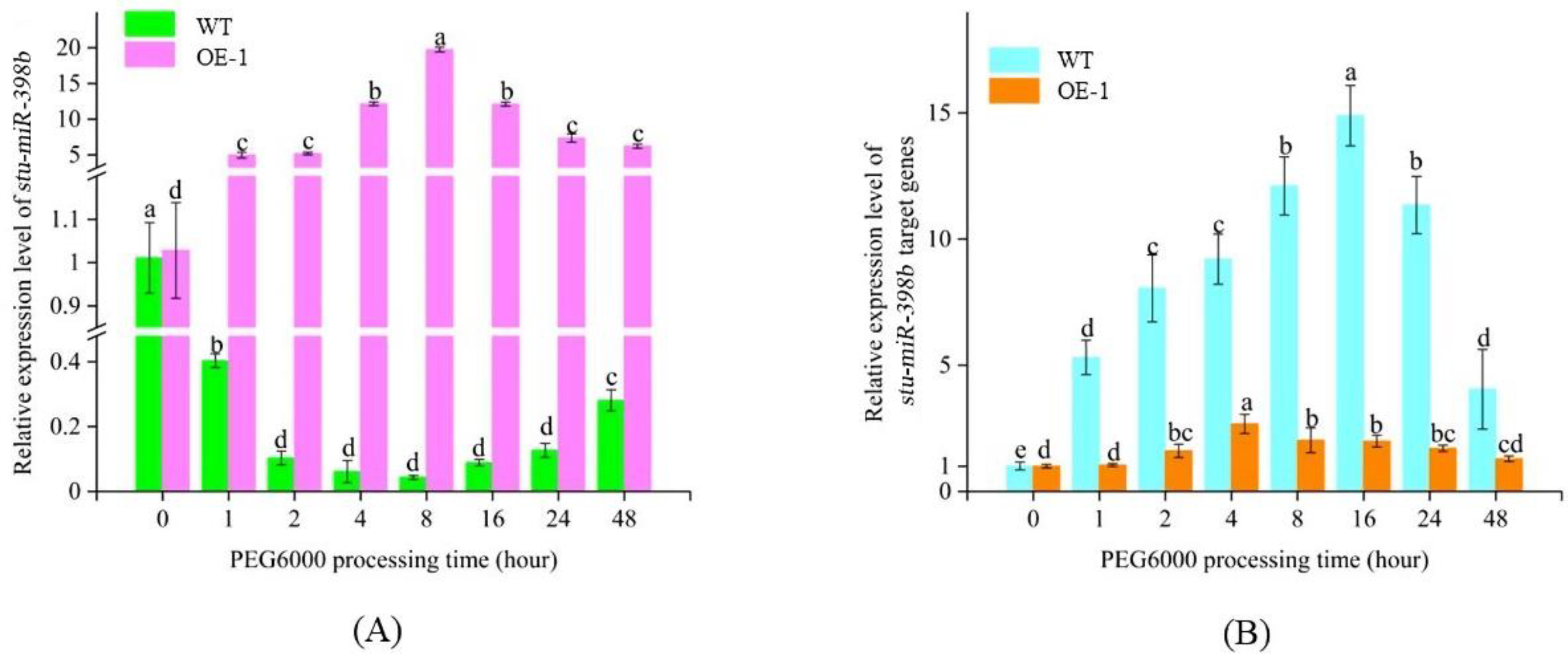
2.6. Overexpression of Stu-miR398b-3p Inhibits the Expression of Its Target Genes
2.7. Stu-miR398b-3p Negatively Regulates Drought Tolerance of Potato
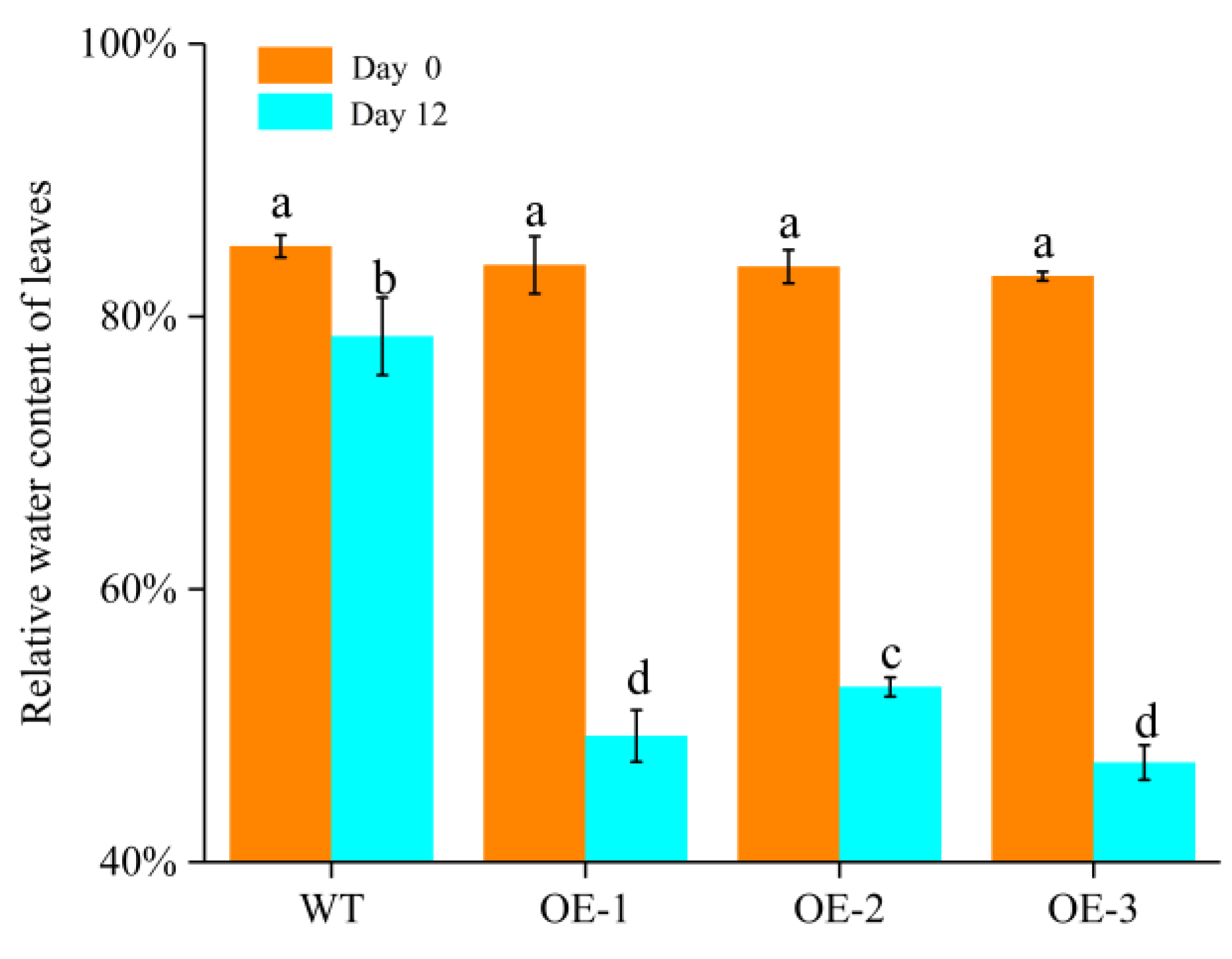
2.7.1. Natural Drought Treatment
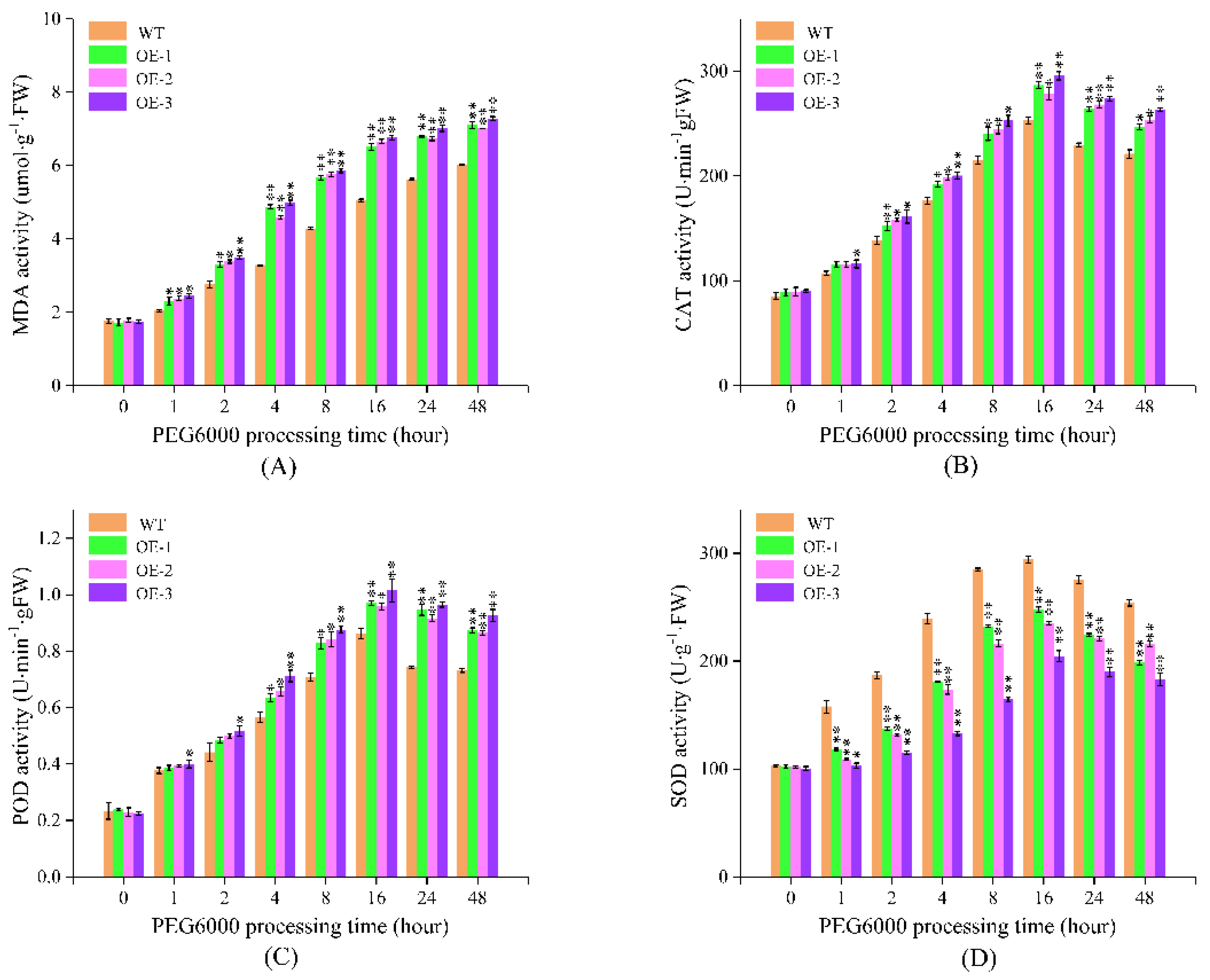
2.7.2. PEG6000 Simulated Drought Treatment
3. Discussion
3.1. In Vivo Cleavage of Cu/Zn SOD mRNA Mediated by Stu-miR398b-3p
3.2. Stu-miR398b-3p Negatively Regulates Drought Tolerance of Potato
4. Materials and Methods
4.1. Vector and Reagents
4.2. Potato Materials and Drought Treatments
4.3. Prediction of Stu-miR398b-3p Target Gene
4.4. RLM-5′RACE Method to Verify the Target Gene of Stu-miR398b-3p
4.5. Bioinformatics Analysis of Stu-miR398b-3p Target Genes
4.6. Construction of Stu-miR398b-3p Artificial miRNA Expression Vector
4.7. Potato Genetic Transformation and Identification of Transgenic Plants
4.8. Drought Resistance Analysis of Transgenic Plants
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Llave, C.; Xie, Z.; Kasschau, K.D.; Carrington, J.C. Cleavage of Scarecrow-like mRNA targets directed by a class of Arabidopsis miRNA. Science 2002, 297, 2053–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunkar, R.; Girke, T.; Jain, P.K.; Zhu, J.K. Cloning and characterization of microRNAs from rice. Plant Cell 2005, 17, 1397–1411. [Google Scholar] [CrossRef] [Green Version]
- Mica, E.; Gianfranceschi, L.; Pè, M.E. Characterization of five microRNA families in maize. J. Exp. Bot. 2006, 57, 2601–2612. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Guo, G.; Ni, Z.; Sunkar, R.; Du, J.; Zhu, J.K.; Sun, Q. Cloning and characterization of microRNAs from wheat (Triticum aestivum L.). Genome Biol. 2007, 8, R96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Hu, X.; Chen, J.; Wang, W.; Xiong, X.; He, C. Integrated mRNA and microRNA transcriptome analysis reveals miRNA regulation in response to PVA in potato. Sci. Rep. 2017, 7, 16925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bansal, K.C.; Nagaragan, S.; Sukumaran, N.P. A rapid screening technique for drought resistance in potato (Solanum tuberosum L). Potato Res. 1991, 34, 241–248. [Google Scholar] [CrossRef]
- Kirch, H.H.; van Berkel, J.; Glaczinski, H.; Salamini, F.; Gebhardt, C. Structural organization, expression and promoter activity of a cold-stress-inducible gene of potato (Solanum tuberosum L.). Plant Mol. Biol. 1997, 33, 897–909. [Google Scholar] [CrossRef]
- Zhang, N.; Yang, J.; Wang, Z.; Wen, Y.; Wang, J.; He, W.; Liu, B.; Si, H.; Wang, D. Identification of novel and conserved microRNAs related to drought stress in potato by deep sequencing. PLoS ONE 2014, 9, e95489. [Google Scholar] [CrossRef]
- Zhu, C.; Ding, Y.; Liu, H. MiR398 and plant stress responses. Physiol. Plant 2011, 143, 1–9. [Google Scholar] [CrossRef]
- Xu, J.; Duan, X.; Yang, J.; Beeching, J.R.; Zhang, P. Coupled expression of Cu/Zn-superoxide dismutase and catalase in cassava improves tolerance against cold and drought stresses. Plant Signal Behav. 2013, 8, e24525. [Google Scholar] [CrossRef]
- Tian, L.; Li, J.; Huang, C.; Zhang, D.; Xu, Y.; Yang, X.; Song, J.; Wang, D.; Qiu, N.; Short, D.P.G.; et al. Cu/Zn superoxide dismutase (VdSOD1) mediates reactive oxygen species detoxification and modulates virulence in Verticillium dahliae. Mol. Plant Pathol. 2021, 22, 1092–1108. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Liu, C.; Cheng, Y.; Ruan, M.; Ye, Q.; Wang, R.; Yao, Z.; Wan, H. Molecular evolution and functional divergence of stress-responsive Cu/Zn superoxide dismutases in plants. Int. J. Mol. Sci. 2022, 23, 7082. [Google Scholar] [CrossRef]
- Guan, Q.; Lu, X.; Zeng, H.; Zhang, Y.; Zhu, J. Heat stress induction of miR398 triggers a regulatory loop that is critical for thermotolerance in Arabidopsis. Plant J. 2013, 74, 840–851. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, J.; Song, A.; Chen, S.; Shan, H.; Luo, H.; Gu, C.; Sun, J.; Zhu, L.; Fang, W.; et al. Ambient temperature enhanced freezing tolerance of Chrysanthemum dichrum CdICE1 Arabidopsis via miR398. BMC Biol. 2013, 11, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jovanović, Ž.; Stanisavljević, N.; Mikić, A.; Radović, S.; Maksimović, V. Water deficit down-regulates miR398 and miR408 in pea (Pisum sativum L.). Plant Physiol. Biochem. 2014, 83, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Juszczak, I.; Baier, M. The strength of the miR398-Csd2-CCS1 regulon is subject to natural variation in Arabidopsis thaliana. FEBS Lett. 2012, 586, 3385–3390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, G.; Ai, Q.; Yu, D. Uncovering miRNAs involved in crosstalk between nutrient deficiencies in Arabidopsis. Sci. Rep. 2015, 5, 11813. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.; He, H.; Li, Y.; Yu, D. A new strategy for construction of artificial miRNA vectors in Arabidopsis. Planta 2012, 235, 1421–1429. [Google Scholar] [CrossRef]
- Winter, J.; Jung, S.; Keller, S.; Gregory, R.I.; Diederichs, S. Many roads to maturity: microRNA biogenesis pathways and their regulation. Nat. Cell Biol. 2009, 11, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, Q.; Zhu, X.; Cui, M.; Jia, H.; Zhang, W.; Tang, W.; Leng, X.; Shen, W. Characterization on the conservation and diversification of miRNA156 gene family from lower to higher plant species based on phylogenetic analysis at the whole genomic level. Funct. Integr. Genomics 2019, 19, 933–952. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; He, J.; Ping, Y.; Guo, J.; Hou, N.; Cao, F.; Li, X.; Geng, D.; Wang, S.; Chen, P.; et al. The positive feedback regulatory loop of miR160-Auxin Response Factor 17-HYPONASTIC LEAVES 1 mediates drought tolerance in apple trees. Plant Physiol. 2022, 188, 1686–1708. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Pandey, D.K.; Chaudhary, B. Target-mimicry based diminution of miRNA167 reinforced flowering-time phenotypes in tobacco via spatial-transcriptional biases of flowering-associated miRNAs. Gene 2019, 682, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Vakilian, K.A. Machine learning improves our knowledge about miRNA functions towards plant abiotic stresses. Sci. Rep. 2020, 10, 3041. [Google Scholar] [CrossRef] [Green Version]
- Smoczynska, A.; Szweykowska-Kulinska, Z. MicroRNA-mediated regulation of flower development in grasses. Acta Biochim. Pol. 2016, 63, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhang, X.; Wang, W.; Wang, Y.; Ming, F. The suppression of WRKY44 by GIGANTEA-miR172 pathway is involved in drought response of Arabidopsis thaliana. PLoS ONE 2013, 8, e73541. [Google Scholar] [CrossRef]
- Xiong, W.; Zhao, Y.; Gao, H.; Li, Y.; Tang, W.; Ma, L.; Yang, G.; Sun, J. Genomic characterization and expression analysis of TCP transcription factors in Setaria italica and Setaria viridis. Plant Signal Behav. 2022, 17, 2075158. [Google Scholar] [CrossRef]
- Gu, M.; Xu, K.; Chen, A.; Zhu, Y.; Tang, G.; Xu, G. Expression analysis suggests potential roles of microRNAs for phosphate and arbuscular mycorrhizal signaling in Solanum lycopersicum. Physiol. Plant. 2010, 138, 226–237. [Google Scholar] [CrossRef]
- Yan, Q.; Wu, F.; Yan, Z.; Li, J.; Ma, T.; Zhang, Y.; Zhao, Y.; Wang, Y.; Zhang, J. Differential co-expression networks of long non-coding RNAs and mRNAs in Cleistogenes songorica under water stress and during recovery. BMC Plant Biol. 2019, 19, 23. [Google Scholar] [CrossRef] [Green Version]
- Dugas, D.V.; Bartel, B. Sucrose induction of Arabidopsis miR398 represses two Cu/Zn superoxide dismutases. Plant Mol. Biol. 2008, 67, 403–417. [Google Scholar] [CrossRef]
- Sunkar, R.; Kapoor, A.; Zhu, J.K. Posttranscriptional induction of two Cu/Zn superoxide dismutase genes in Arabidopsis is mediated by downregulation of miR398 and important for oxidative stress tolerance. Plant Cell 2006, 18, 2051–2065. [Google Scholar] [CrossRef]
- Lu, Y.; Feng, Z.; Bian, L.; Xie, H.; Liang, J. miR398 regulation in rice of the responses to abiotic and biotic stresses depends on CSD1 and CSD2 expression. Funct Plant Biol. 2010, 38, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Li, M.; Zhang, Z.; Tie, W.; Chen, X.; Jin, L.; Zhai, N.; Zheng, Q.; Zhang, J.; Wang, R.; et al. Integrated mRNA and microRNA analysis identifies genes and small miRNA molecules associated with transcriptional and post-transcriptional-level responses to both drought stress and re-watering treatment in tobacco. BMC Genom. 2017, 18, 62. [Google Scholar] [CrossRef] [Green Version]
- Chávez-Hernández, E.C.; Alejandri-Ramírez, N.D.; Juárez-González, V.T.; Dinkova, T.D. Maize miRNA and target regulation in response to hormone depletion and light exposure during somatic embryogenesis. Front Plant Sci. 2015, 6, 555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, M.R.; Sambrook, J. Rapid Amplification of Sequences from the 5′ Ends of mRNAs: 5′-RACE. Cold Spring Harb. Protoc. 2019, 2019. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Primer Sequences (5′–3′) |
|---|---|
| I miR-s | gaTTGTGTTCTCAGGTCACCCCTtctctcttttgtattcc |
| II miR-a | gaAGGGGTGACCTGAGAACACAAtcaaagagaatcaatga |
| III miR*s | gaAGAGGTGACCTGACAACACATtcacaggtcgtgatatg |
| IV miR*a | gaATGTGTTGTCAGGTCACCTCTtctacatatatattcct |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Z.; Yang, J.; Wang, X.; Zhang, N.; Si, H. Potato Stu-miR398b-3p Negatively Regulates Cu/Zn-SOD Response to Drought Tolerance. Int. J. Mol. Sci. 2023, 24, 2525. https://doi.org/10.3390/ijms24032525
Zheng Z, Yang J, Wang X, Zhang N, Si H. Potato Stu-miR398b-3p Negatively Regulates Cu/Zn-SOD Response to Drought Tolerance. International Journal of Molecular Sciences. 2023; 24(3):2525. https://doi.org/10.3390/ijms24032525
Chicago/Turabian StyleZheng, Zhiyong, Jiangwei Yang, Xiaofeng Wang, Ning Zhang, and Huaijun Si. 2023. "Potato Stu-miR398b-3p Negatively Regulates Cu/Zn-SOD Response to Drought Tolerance" International Journal of Molecular Sciences 24, no. 3: 2525. https://doi.org/10.3390/ijms24032525
APA StyleZheng, Z., Yang, J., Wang, X., Zhang, N., & Si, H. (2023). Potato Stu-miR398b-3p Negatively Regulates Cu/Zn-SOD Response to Drought Tolerance. International Journal of Molecular Sciences, 24(3), 2525. https://doi.org/10.3390/ijms24032525