
DDC-Promoter-Driven Chemogenetic Activation of SNpc Dopaminergic Neurons Alleviates Parkinsonian Motor Symptoms

 , and
, and 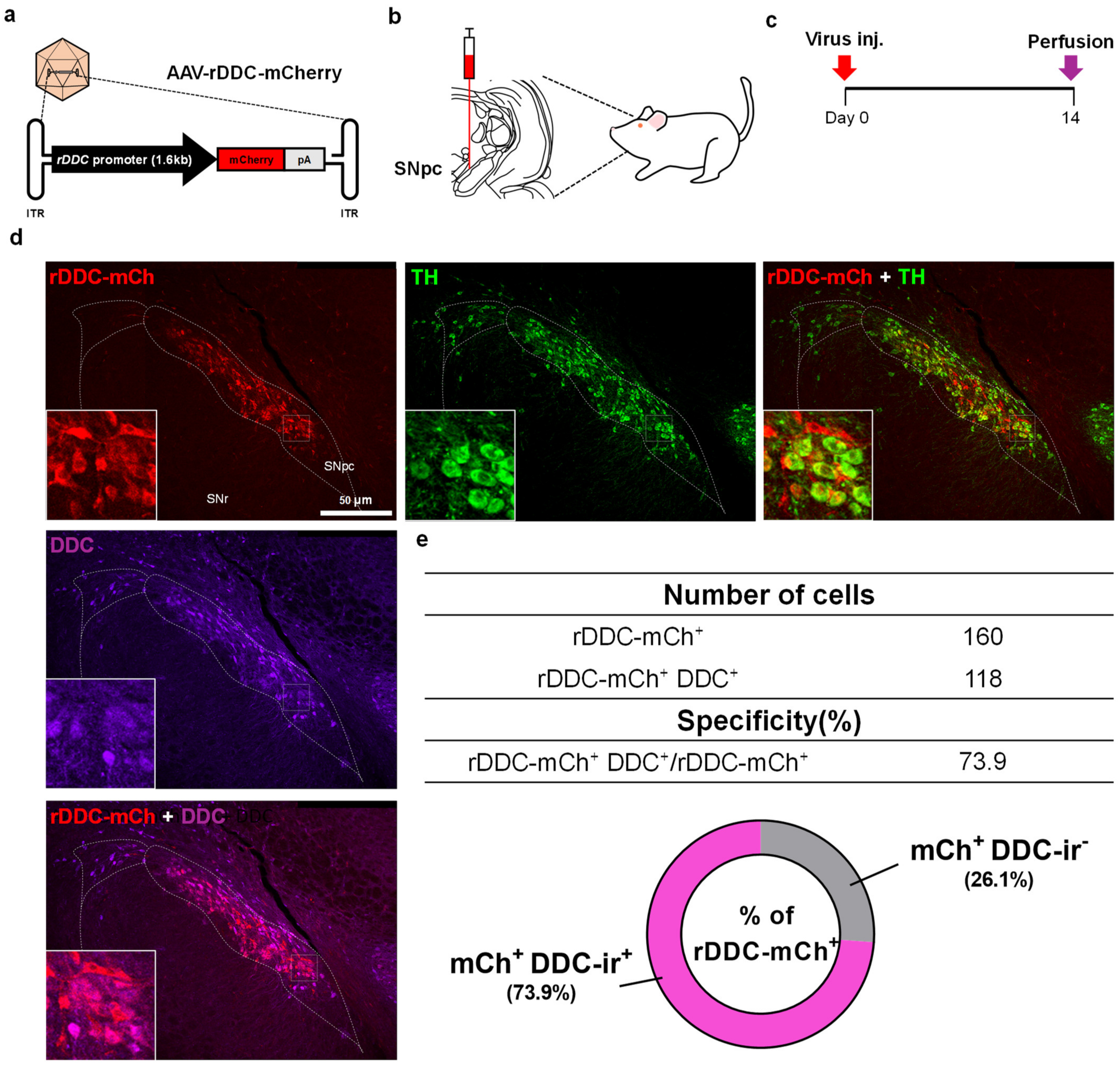{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Design and Specificity Validation of the DDC-Promoter in an AAV Vector
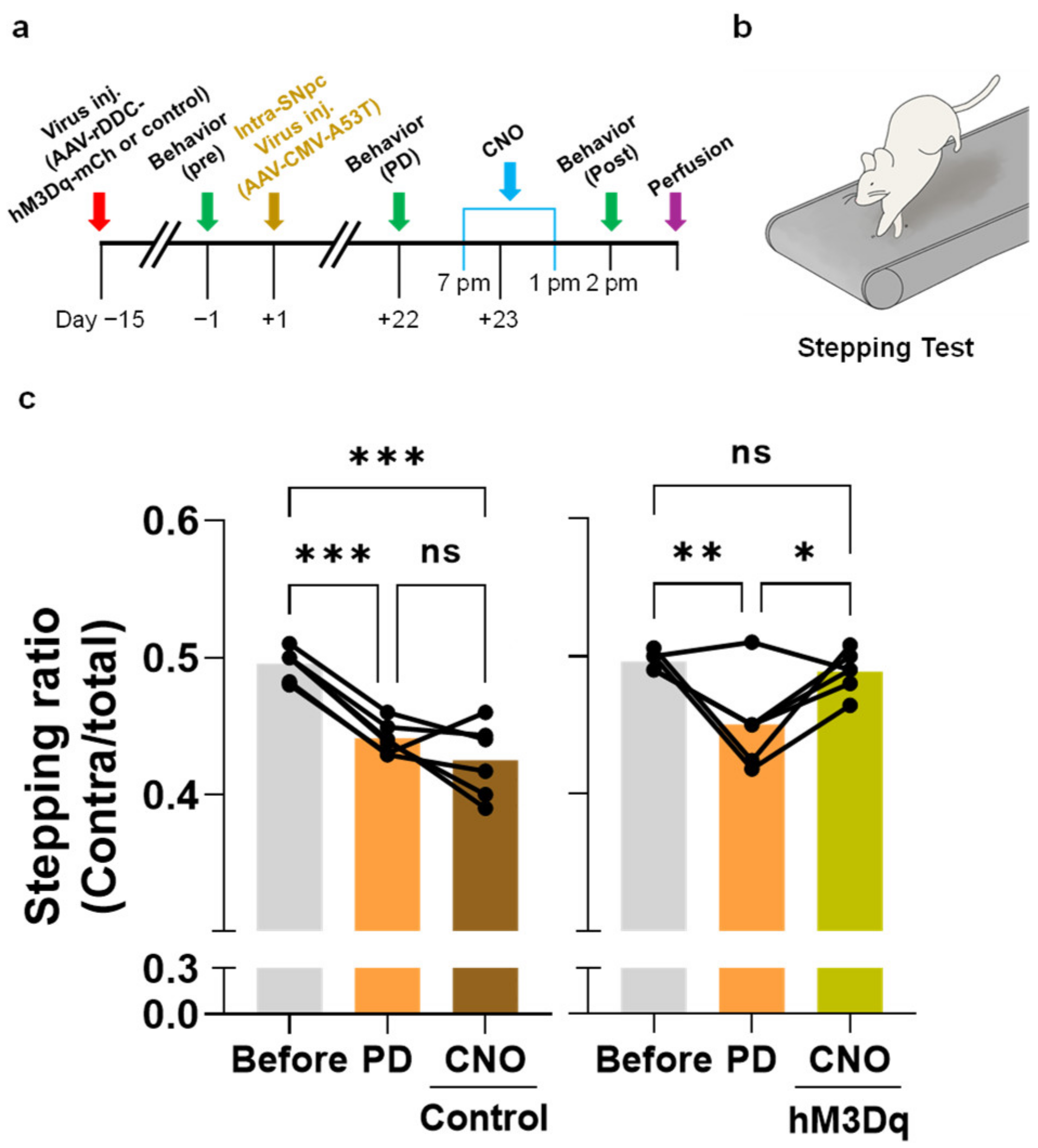
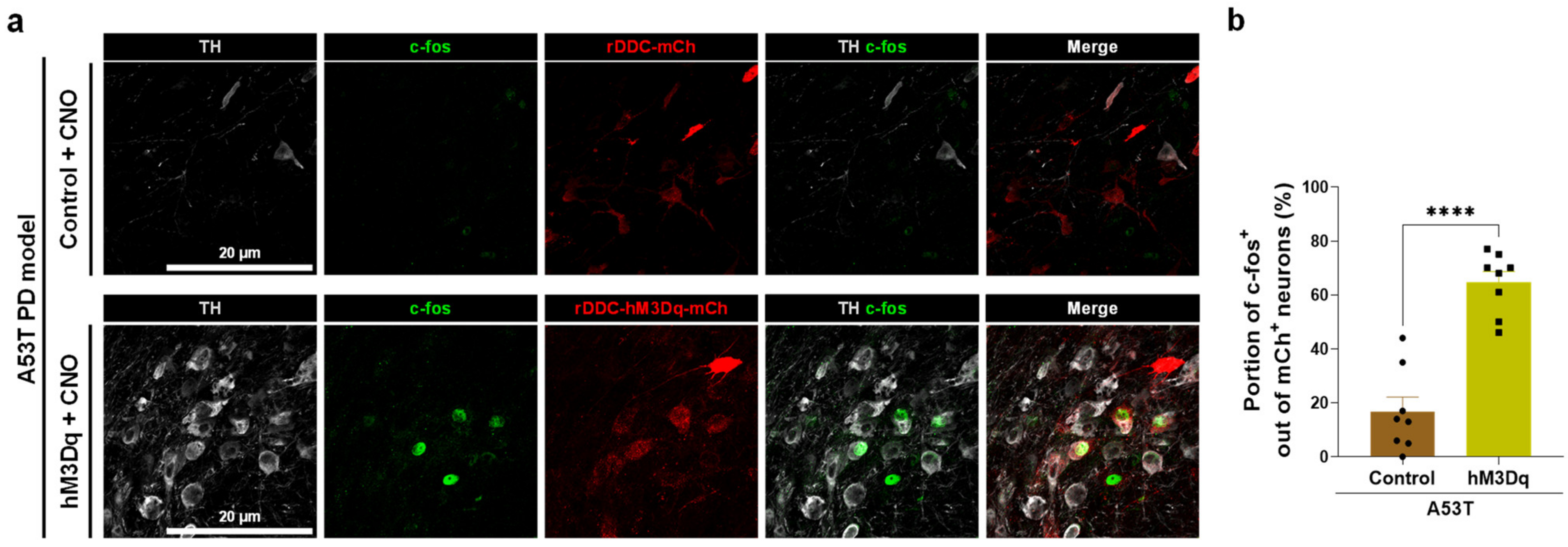
2.2. Chemogenetic Activation of DDC+ Neurons Alleviates PD Motor Symptoms
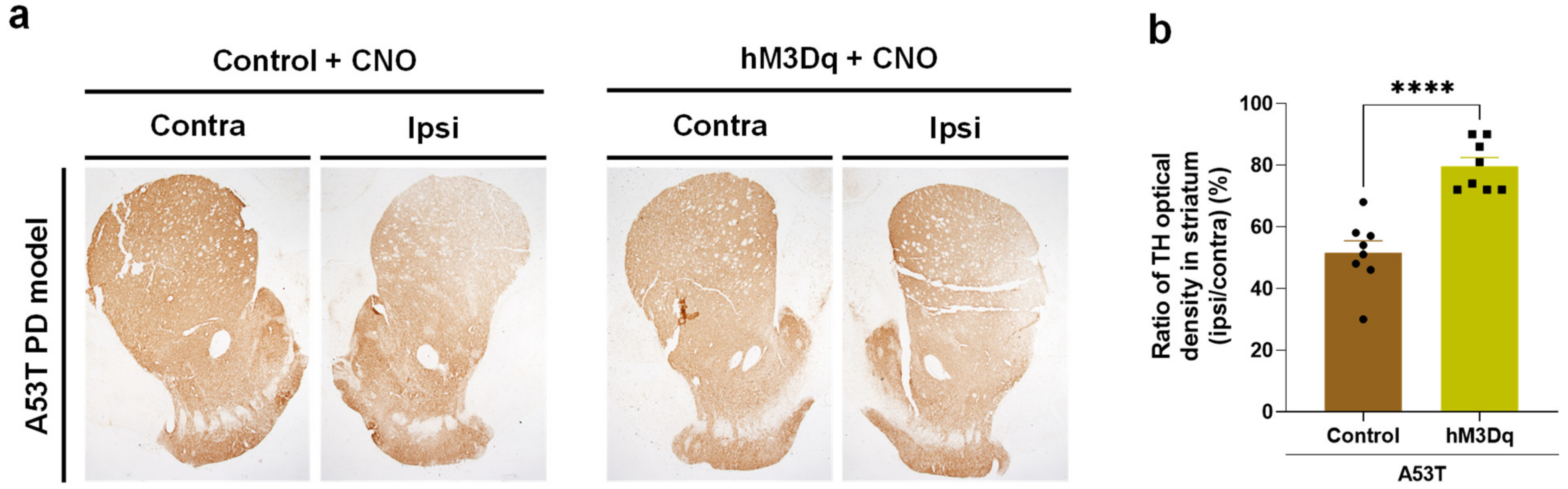
2.3. Chemogenetic Activation of DDC+ Neurons Rescues Nigrostriatal TH Loss
3. Discussion
3.1. The Necessity of DDC Promoter Validation
3.2. Neuromodulation Strategy for PD Treatment
3.3. Advantages of AAV-Mediated Gene Therapy
3.4. Limitation of Chemogenetic Manipulation as a Therapeutic Strategy
4. Materials and Methods
4.1. Vector Design and Virus Production
4.2. Animals
A53T PD Model Preparation
4.3. Chemogenetics
4.4. Spontaneous Action Potential Firing Recording
4.5. Immunofluorescence Staining and Confocal Imaging
4.6. DAB Staining
4.7. Behavioral Tests
4.7.1. Stepping Test
4.7.2. Balance Beam Test
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shalaby, K.E.; El-Agnaf, O.M.A. Gene-Based Therapeutics for Parkinson’s Disease. Biomedicines 2022, 10, 1790. [Google Scholar] [CrossRef] [PubMed]
- Fahn, S. Description of Parkinson’s Disease as a Clinical Syndrome. Ann. N. Y. Acad. Sci. 2003, 991, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Damier, P.; Hirsch, E.C.; Agid, Y.; Graybiel, A. The substantia nigra of the human brain: II. Patterns of loss of dopamine-containing neurons in Parkinson’s disease. Brain 1999, 122, 1437–1448. [Google Scholar] [CrossRef] [PubMed]
- Samii, A.; Nutt, J.G.; Ransom, B.R. Parkinson’s disease. Lancet 2004, 363, 1783–1793. [Google Scholar] [CrossRef]
- Winslow, A.R.; Chen, C.-W.; Corrochano, S.; Acevedo-Arozena, A.; Gordon, D.E.; Peden, A.A.; Lichtenberg, M.; Menzies, F.M.; Ravikumar, B.; Imarisio, S.; et al. α-Synuclein impairs macroautophagy: Implications for Parkinson’s disease. J. Cell Biol. 2010, 190, 1023–1037. [Google Scholar] [CrossRef]
- Hoozemans, J.J.M.; Van Haastert, E.S.; Eikelenboom, P.; De Vos, R.A.; Rozemuller, J.M.; Scheper, W. Activation of the unfolded protein response in Parkinson’s disease. Biochem. Biophys. Res. Commun. 2007, 354, 707–711. [Google Scholar] [CrossRef]
- Selvaraj, S.; Sun, Y.; Watt, J.A.; Wang, S.; Lei, S.; Birnbaumer, L.; Singh, B.B. Neurotoxin-induced ER stress in mouse dopaminergic neurons involves downregulation of TRPC1 and inhibition of AKT/mTOR signaling. J. Clin. Investig. 2012, 122, 1354–1367. [Google Scholar] [CrossRef]
- Conn, K.J.; Gao, W.; McKee, A.; Lan, M.S.; Ullman, M.D.; Eisenhauer, P.B.; Fine, R.E.; Wells, J.M. Identification of the protein disulfide isomerase family member PDIp in experimental Parkinson’s disease and Lewy body pathology. Brain Res. 2004, 1022, 164–172. [Google Scholar] [CrossRef]
- Anglade, P. Apoptosis and autophagy in nigral neurons of patients with Parkinson’s disease. Histol. Histopathol. 1997, 12, 25–31. [Google Scholar]
- Qiu, J.; Tan, Y.-W.; Hagenston, A.M.; Martel, M.-A.; Kneisel, N.; Skehel, P.A.; Wyllie, D.; Bading, H.; Hardingham, G.E. Mitochondrial calcium uniporter Mcu controls excitotoxicity and is transcriptionally repressed by neuroprotective nuclear calcium signals. Nat. Commun. 2013, 4, 2034. [Google Scholar] [CrossRef]
- Guzman, J.N.; Sanchez-Padilla, J.; Wokosin, D.; Kondapalli, J.; Ilijic, E.; Schumacker, P.T.; Surmeier, D.J. Oxidant stress evoked by pacemaking in dopaminergic neurons is attenuated by DJ-1. Nature 2010, 468, 696–700. [Google Scholar] [CrossRef]
- Heo, J.Y.; Nam, M.-H.; Yoon, H.H.; Kim, J.; Hwang, Y.J.; Won, W.; Woo, D.H.; Lee, J.A.; Park, H.-J.; Jo, S.; et al. Aberrant Tonic Inhibition of Dopaminergic Neuronal Activity Causes Motor Symptoms in Animal Models of Parkinson’s Disease. Curr. Biol. 2020, 30, 276–291.e9. [Google Scholar] [CrossRef]
- An, H.; Heo, J.Y.; Lee, C.J.; Nam, C.J.L.A.M.-H. The Pathological Role of Astrocytic MAOB in Parkinsonism Revealed by Genetic Ablation and Over-expression of MAOB. Exp. Neurobiol. 2021, 30, 113–119. [Google Scholar] [CrossRef]
- Nam, M.-H.; Park, J.H.; Song, H.J.; Choi, J.W.; Kim, S.; Jang, B.K.; Yoon, H.H.; Heo, J.Y.; Lee, H.; An, H.; et al. KDS2010, a Newly Developed Reversible MAO-B Inhibitor, as an Effective Therapeutic Candidate for Parkinson’s Disease. Neurotherapeutics 2021, 18, 1729–1747. [Google Scholar] [CrossRef]
- Nam, M.H.; Sa, M.; Ju, Y.H.; Park, M.G.; Lee, C.J. Revisiting the Role of Astrocytic MAOB in Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23, 4453. [Google Scholar] [CrossRef]
- Brown, B.D.; Gentner, B.; Cantore, A.; Colleoni, S.; Amendola, M.; Zingale, A.; Baccarini, A.; Lazzari, G.; Galli, C.; Naldini, L. Endogenous microRNA can be broadly exploited to regulate transgene expression according to tissue, lineage and differentiation state. Nat. Biotechnol. 2007, 25, 1457–1467. [Google Scholar] [CrossRef]
- Mátrai, J.; Cantore, A.; Bartholomae, C.C.; Annoni, A.; Wang, W.; Acosta-Sanchez, A.; Samara-Kuko, E.; De Waele, L.; Ma, L.; Genovese, P.; et al. Hepatocyte-targeted expression by integrase-defective lentiviral vectors induces antigen-specific tolerance in mice with low genotoxic risk. Hepatology 2011, 53, 1696–1707. [Google Scholar] [CrossRef]
- Alexander, G.M.; Rogan, S.C.; Abbas, A.I.; Armbruster, B.N.; Pei, Y.; Allen, J.A.; Nonneman, R.J.; Hartmann, J.; Moy, S.S.; Nicolelis, M.A.; et al. Remote control of neuronal activity in transgenic mice expressing evolved G protein-coupled receptors. Neuron 2009, 63, 27–39. [Google Scholar] [CrossRef]
- Sato, H.; Arawaka, S.; Hara, S.; Fukushima, S.; Koga, K.; Koyama, S.; Kato, T. Authentically phosphorylated α-synuclein at Ser129 accelerates neurodegeneration in a rat model of familial Parkinson’s disease. J. Neurosci. 2011, 31, 16884–16894. [Google Scholar] [CrossRef]
- Niwa, H.; Yamamura, K.; Miyazaki, J. Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene 1991, 108, 193–199. [Google Scholar]
- Yoon, H.H.; Park, J.H.; Kim, Y.H.; Min, J.; Hwang, E.; Lee, C.J.; Suh, J.-K.F.; Hwang, O.; Jeon, S.R. Optogenetic inactivation of the subthalamic nucleus improves forelimb akinesia in a rat model of parkinson disease. Neurosurgery 2014, 74, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Miyanishi, K.; Choudhury, M.E.; Watanabe, M.; Kubo, M.; Nomoto, M.; Yano, H.; Tanaka, J. Behavioral tests predicting striatal dopamine level in a rat hemi-Parkinson’s disease model. Neurochem. Int. 2019, 122, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Gordián-Vélez, W.J.; Chouhan, D.; España, R.A.; Chen, H.I.; Burdick, J.A.; Duda, J.E.; Cullen, D.K. Restoring lost nigrostriatal fibers in Parkinson’s disease based on clinically-inspired design criteria. Brain Res. Bull. 2021, 175, 168–185. [Google Scholar] [CrossRef] [PubMed]
- Joh, T.H.; Park, D.H.; Reis, D.J. Direct phosphorylation of brain tyrosine hydroxylase by cyclic AMP-dependent protein kinase: Mechanism of enzyme activation. Proc. Natl. Acad. Sci. USA 1978, 75, 4744–4748. [Google Scholar] [CrossRef] [PubMed]
- Kilbourne, E.J.; Nankova, B.B.; Lewis, E.J.; McMahon, A.; Osaka, H.; Sabban, D.B.; Sabban, E.L. Regulated expression of the tyrosine hydroxylase gene by membrane depolarization. Identification of the responsive element and possible second messengers. J. Biol. Chem. 1992, 267, 7563–7569. [Google Scholar] [CrossRef]
- Aumann, T.D.; Egan, K.; Lim, J.; Boon, W.C.; Bye, C.R.; Chua, H.K.; Baban, N.; Parish, C.L.; Bobrovskaya, L.; Dickson, P.; et al. Neuronal activity regulates expression of tyrosine hydroxylase in adult mouse substantia nigra pars compacta neurons. J. Neurochem. 2011, 116, 646–658. [Google Scholar] [CrossRef]
- Fougère, M.; Van Der Zouwen, C.I.; Boutin, J.; Neszvecsko, K.; Sarret, P.; Ryczko, D. Optogenetic stimulation of glutamatergic neurons in the cuneiform nucleus controls locomotion in a mouse model of Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2021, 118, e2110934118. [Google Scholar] [CrossRef]
- Yoon, H.H.; Nam, M.H.; Choi, I.; Min, J.; Jeon, S.R. Optogenetic inactivation of the entopeduncular nucleus improves forelimb akinesia in a Parkinson’s disease model. Behav. Brain Res. 2020, 386, 112551. [Google Scholar] [CrossRef]
- Kravitz, A.V.; Freeze, B.S.; Parker, P.R.L.; Kay, K.; Thwin, M.T.; Deisseroth, K.; Kreitzer, A.C. Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry. Nature 2010, 466, 622–626. [Google Scholar] [CrossRef]
- Ztaou, S.; Maurice, N.; Camon, J.; Guiraudie-Capraz, G.; Kerkerian-Le Goff, L.; Beurrier, C.; Liberge, M.; Amalric, M. Involvement of Striatal Cholinergic Interneurons and M1 and M4 Muscarinic Receptors in Motor Symptoms of Parkinson’s Disease. J. Neurosci. 2016, 36, 9161. [Google Scholar] [CrossRef]
- Tanimura, A.; Du, Y.; Kondapalli, J.; Wokosin, D.L.; Surmeier, D.J. Cholinergic interneurons amplify thalamostriatal excitation of striatal indirect pathway neurons in Parkinson’s disease models. Neuron 2019, 101, 444–458. [Google Scholar] [CrossRef]
- Sharma, P.K.; Wells, L.; Rizzo, G.; Elson, J.L.; Passchier, J.; Rabiner, E.A.; Gunn, R.N.; Dexter, D.T.; Pienaar, I.S. DREADD activation of pedunculopontine cholinergic neurons reverses motor deficits and restores striatal dopamine signaling in parkinsonian rats. Neurotherapeutics 2020, 17, 1120–1141. [Google Scholar] [CrossRef]
- Chen, K.; Yang, G.; So, K.-F.; Zhang, L. Activation of cortical somatostatin interneurons rescues synapse loss and motor deficits after acute MPTP infusion. Iscience 2019, 17, 230–241. [Google Scholar] [CrossRef]
- McIver, E.L.; Atherton, J.F.; Chu, H.-Y.; Cosgrove, K.E.; Kondapalli, J.; Wokosin, D.; Surmeier, D.J.; Bevan, M.D. Maladaptive downregulation of autonomous subthalamic nucleus activity following the loss of midbrain dopamine neurons. Cell Rep. 2019, 28, 992–1002. [Google Scholar] [CrossRef]
- Avila, C.; Kucinski, A.; Sarter, M. Complex movement control in a rat model of Parkinsonian falls: Bidirectional control by striatal cholinergic interneurons. J. Neurosci. 2020, 40, 6049–6067. [Google Scholar] [CrossRef]
- Alcacer, C.; Andreoli, L.; Sebastianutto, I.; Jakobsson, J.; Fieblinger, T.; Cenci, M.A. Chemogenetic stimulation of striatal projection neurons modulates responses to Parkinson’s disease therapy. J. Clin. Investig. 2017, 127, 720–734. [Google Scholar] [CrossRef]
- Lee, E.J.; Yoon, H.H.; Park, E.S.; Min, J.; Jeon, S.R. A Novel Animal Model of Parkinson’s Disease Using Optogenetics: Representation of Various Disease Stages by Modulating the Illumination Parameter. Stereotact. Funct. Neurosurg. 2018, 96, 22–32. [Google Scholar] [CrossRef]
- Yu, C.; Cassar, I.R.; Sambangi, J.; Grill, W.M. Frequency-Specific Optogenetic Deep Brain Stimulation of Subthalamic Nucleus Improves Parkinsonian Motor Behaviors. J. Neurosci. 2020, 40, 4323–4334. [Google Scholar] [CrossRef]
- Lu, Y.; Driscoll, N.; Ozden, I.; Yu, Z.; Nurmikko, A.V. Modulating dopamine release by optogenetics in transgenic mice reveals terminal dopaminergic dynamics. Neurophotonics 2015, 2, 031207. [Google Scholar] [CrossRef]
- De Almeida, L.P.; Zala, D.; Aebischer, P.; Deglon, N. Neuroprotective effect of a CNTF-expressing lentiviral vector in the quinolinic acid rat model of Huntington’s disease. Neurobiol. Dis. 2001, 8, 433–446. [Google Scholar] [CrossRef]
- Stone, D. Novel viral vector systems for gene therapy. Viruses 2010, 2, 1002–1007. [Google Scholar] [CrossRef] [PubMed]
- Nayerossadat, N.; Maedeh, T.; Ali, P.A. Viral and nonviral delivery systems for gene delivery. Adv. Biomed. Res. 2012, 1, 27. [Google Scholar] [CrossRef] [PubMed]
- Hermens, W.T.; Verhaagen, J. Viral vectors, tools for gene transfer in the nervous system. Prog. Neurobiol. 1998, 55, 399–432. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A. AAV Vectors: Are They Safe? Hum. Gene Ther. 2020, 31, 697–699. [Google Scholar] [CrossRef]
- US Food & Drug Association Hompage. Approved Cellular and Gene Therapy Products. 2022. Available online: https://www.fda.gov/vaccines-blood-biologics/cellular-gene-therapy-products/approved-cellular-and-gene-therapy-products (accessed on 17 January 2023).
- Ingusci, S.; Verlengia, G.; Soukupova, M.; Zucchini, S.; Simonato, M. Gene Therapy Tools for Brain Diseases. Gene Therapy Tools for Brain Diseases. Front. Pharmacol. 2019, 10, 724. [Google Scholar] [CrossRef]
- Roth, B.L. DREADDs for Neuroscientists. Neuron 2016, 89, 683–694. [Google Scholar] [CrossRef]
- Park, J.; Throop, A.L.; LaBaer, J. Site-specific recombinational cloning using gateway and in-fusion cloning schemes. Curr. Protoc. Mol. Biol. 2015, 110, 3–20. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, D.-C.; Ju, Y.H.; Seo, J.-J.; Oh, S.-J.; Lee, C.J.; Lee, S.E.; Nam, M.-H. DDC-Promoter-Driven Chemogenetic Activation of SNpc Dopaminergic Neurons Alleviates Parkinsonian Motor Symptoms. Int. J. Mol. Sci. 2023, 24, 2491. https://doi.org/10.3390/ijms24032491
Seo D-C, Ju YH, Seo J-J, Oh S-J, Lee CJ, Lee SE, Nam M-H. DDC-Promoter-Driven Chemogenetic Activation of SNpc Dopaminergic Neurons Alleviates Parkinsonian Motor Symptoms. International Journal of Molecular Sciences. 2023; 24(3):2491. https://doi.org/10.3390/ijms24032491
Chicago/Turabian StyleSeo, Dong-Chan, Yeon Ha Ju, Jin-Ju Seo, Soo-Jin Oh, C. Justin Lee, Seung Eun Lee, and Min-Ho Nam. 2023. "DDC-Promoter-Driven Chemogenetic Activation of SNpc Dopaminergic Neurons Alleviates Parkinsonian Motor Symptoms" International Journal of Molecular Sciences 24, no. 3: 2491. https://doi.org/10.3390/ijms24032491
APA StyleSeo, D.-C., Ju, Y. H., Seo, J.-J., Oh, S.-J., Lee, C. J., Lee, S. E., & Nam, M.-H. (2023). DDC-Promoter-Driven Chemogenetic Activation of SNpc Dopaminergic Neurons Alleviates Parkinsonian Motor Symptoms. International Journal of Molecular Sciences, 24(3), 2491. https://doi.org/10.3390/ijms24032491