
The Assessment of the Phototoxic Action of Chlortetracycline and Doxycycline as a Potential Treatment of Melanotic Melanoma—Biochemical and Molecular Studies on COLO 829 and G-361 Cell Lines


Abstract
1. Introduction
2. Results
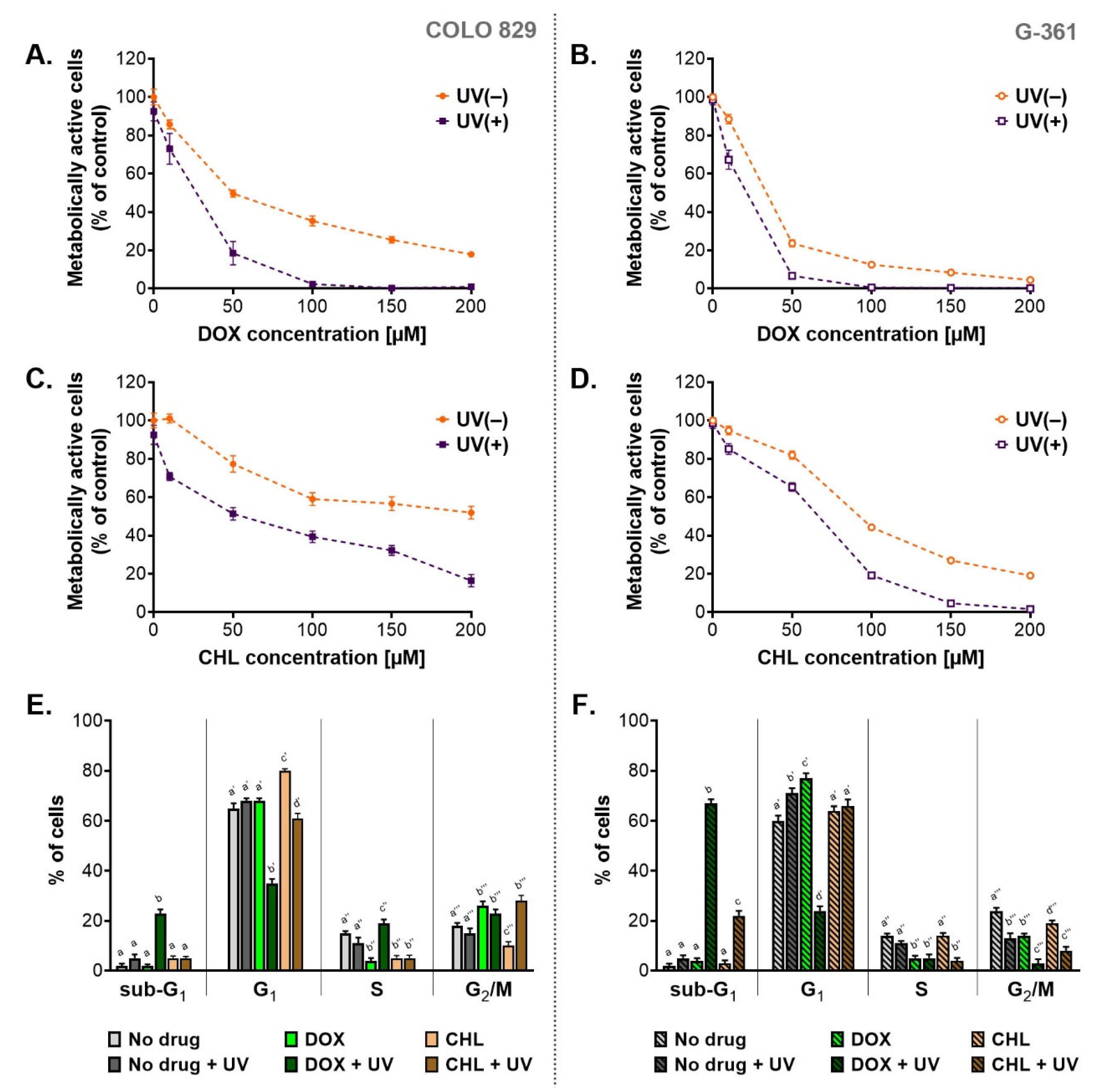
2.1. The Cytotoxic and Phototoxic Actions of Doxycycline and Chlortetracycline Cause a Decrease in the Percentage of Metabolically Active Melanoma Cells and Disturb Cell Cycle
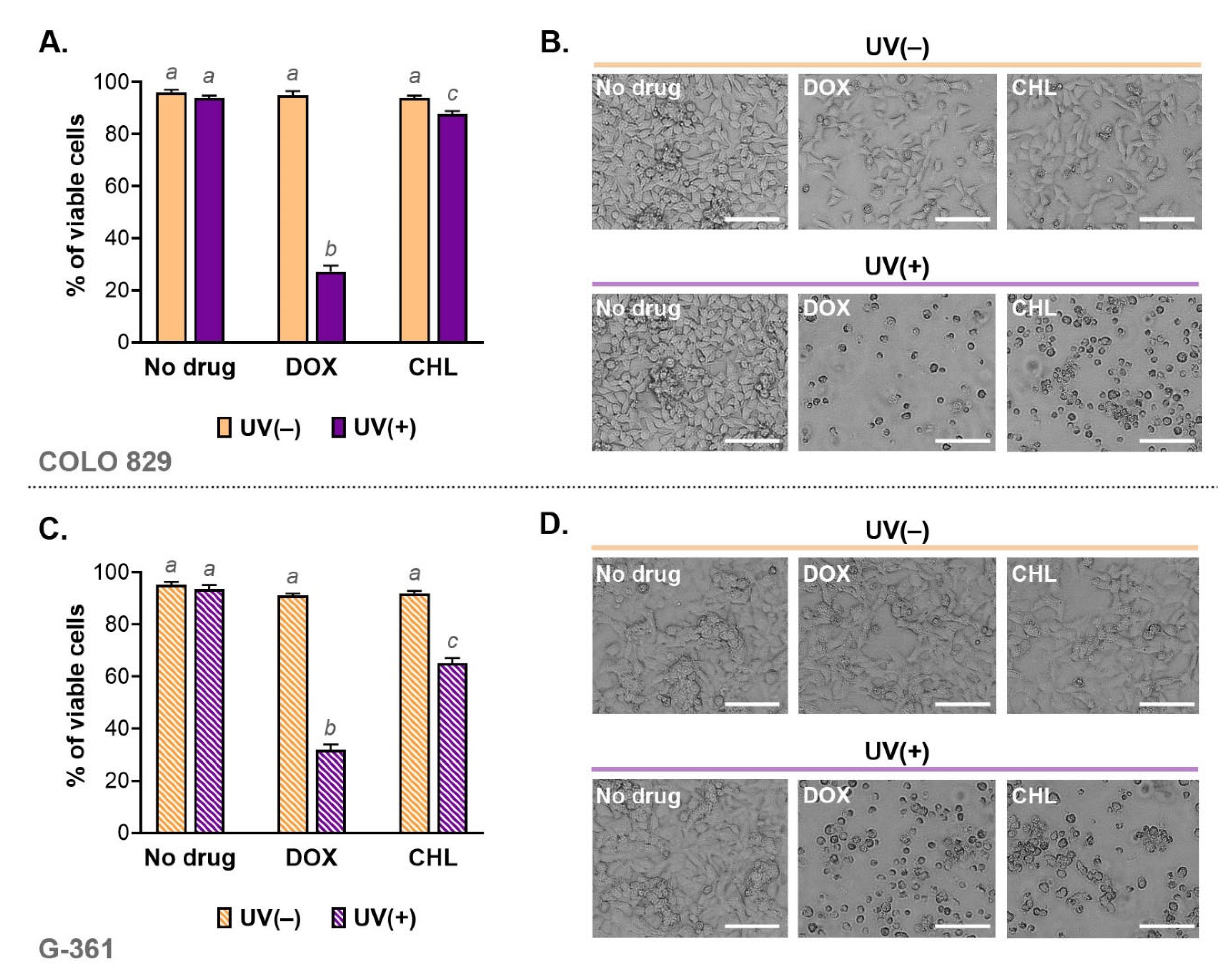
2.2. Phototoxic Action of Doxycycline and Chlortetracycline Causes a Decrease in Melanoma Cell Viability
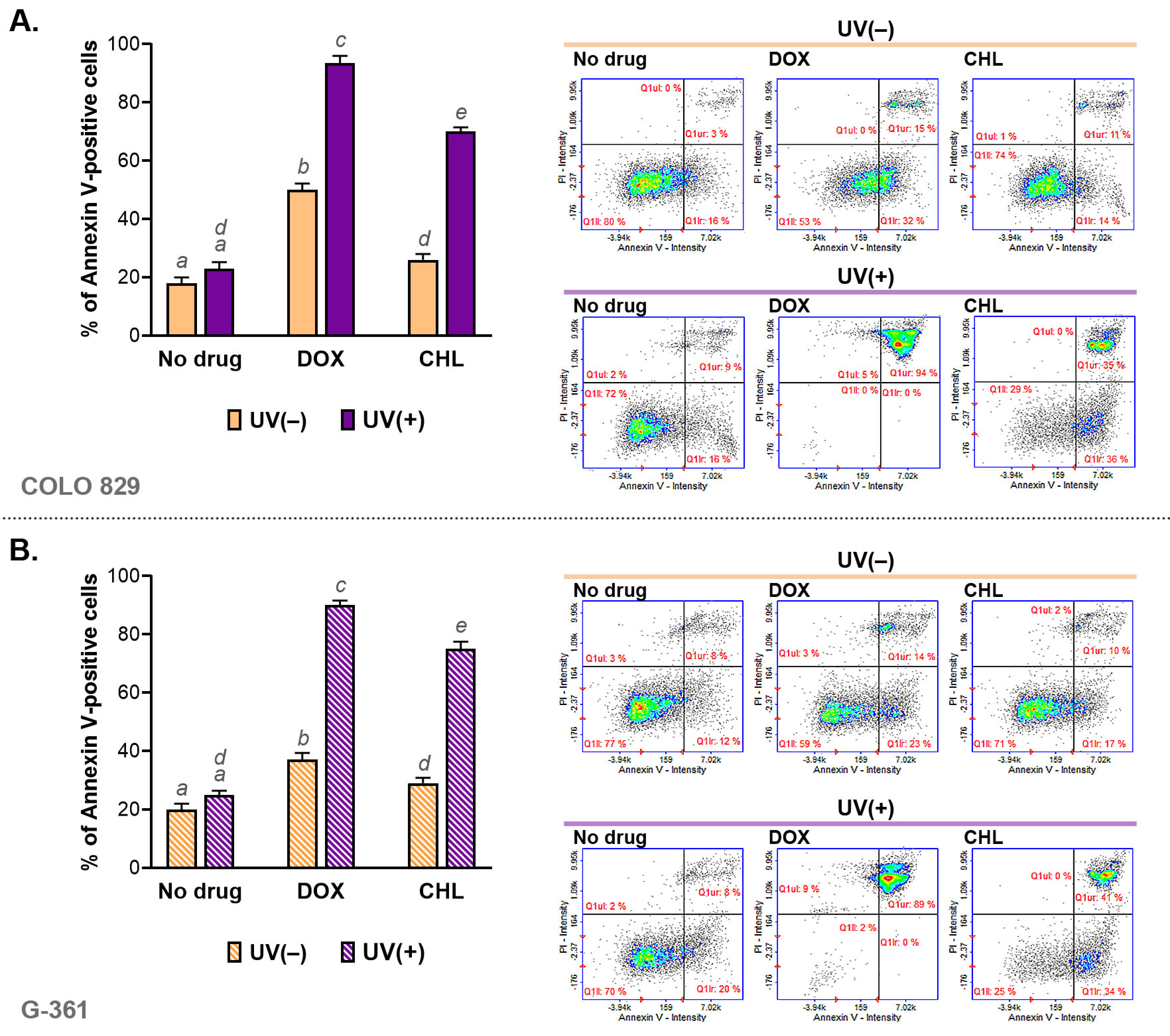
2.3. Analysis of Apoptosis in Melanoma Cells Irradiated with UVA and Treated with Doxycycline and Chlortetracycline
2.4. Analysis of Mitochondrial Membrane Potential (ψm) in Melanoma Cells Treated with Doxycycline or Chlortetracycline and Irradiated with UVA
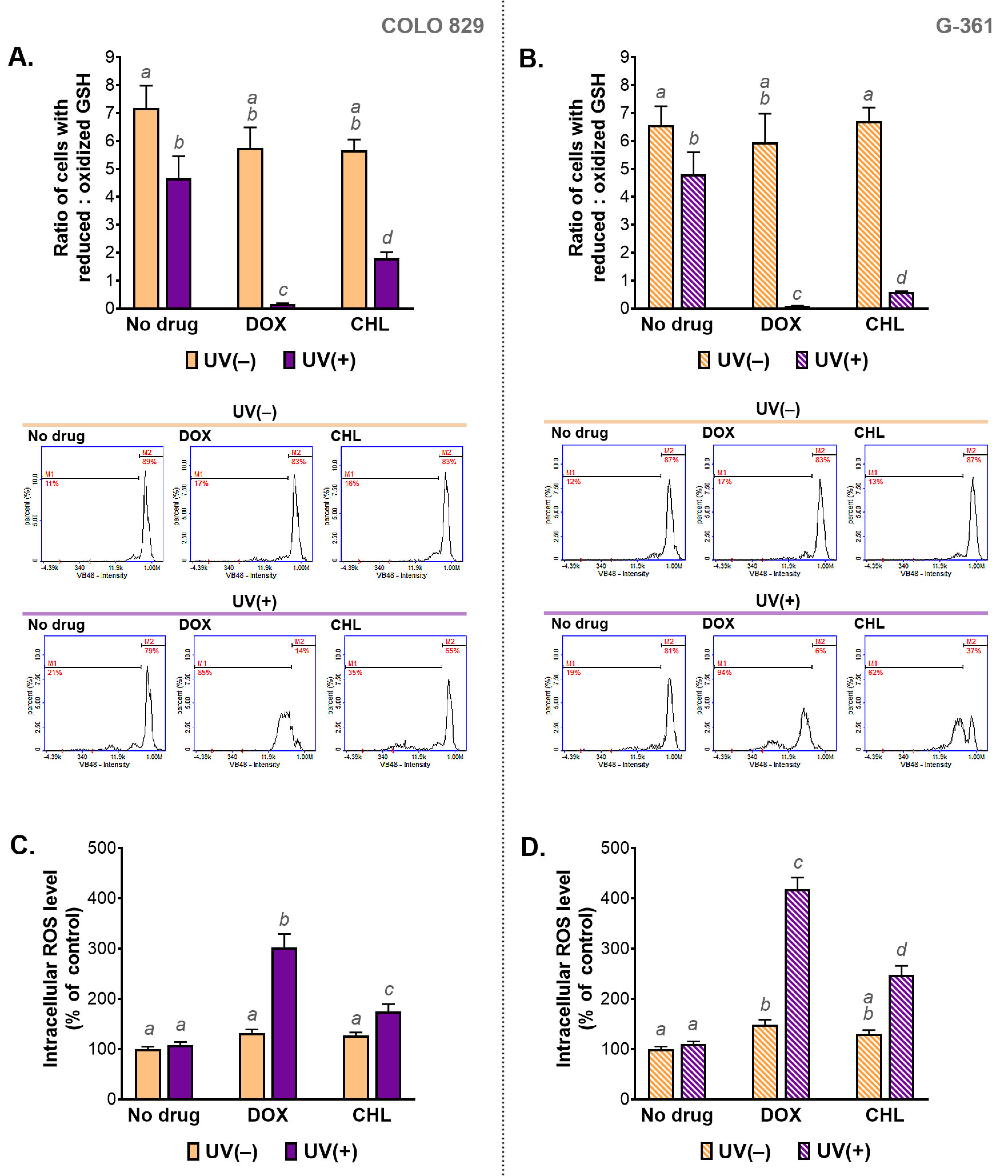
2.5. The Assessment of Redox Homeostasis in Melanoma Cells Treated with Doxycycline and Chlortetracycline and Exposed to UVA Radiation
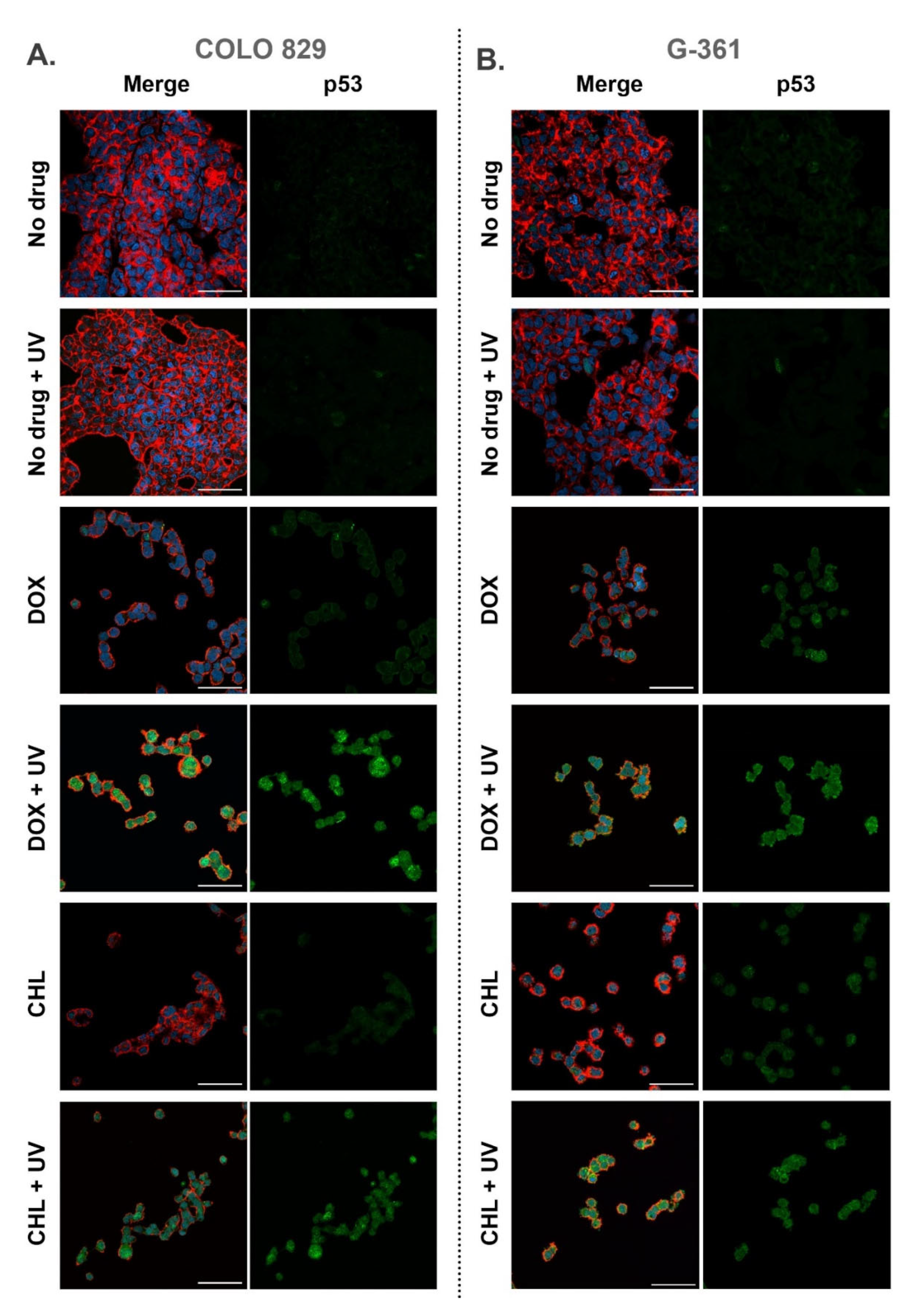
2.6. Confocal Imaging of Melanotic Melanoma Cells Treated with Doxycycline and Chlortetracycline and Exposed to UVA Radiation
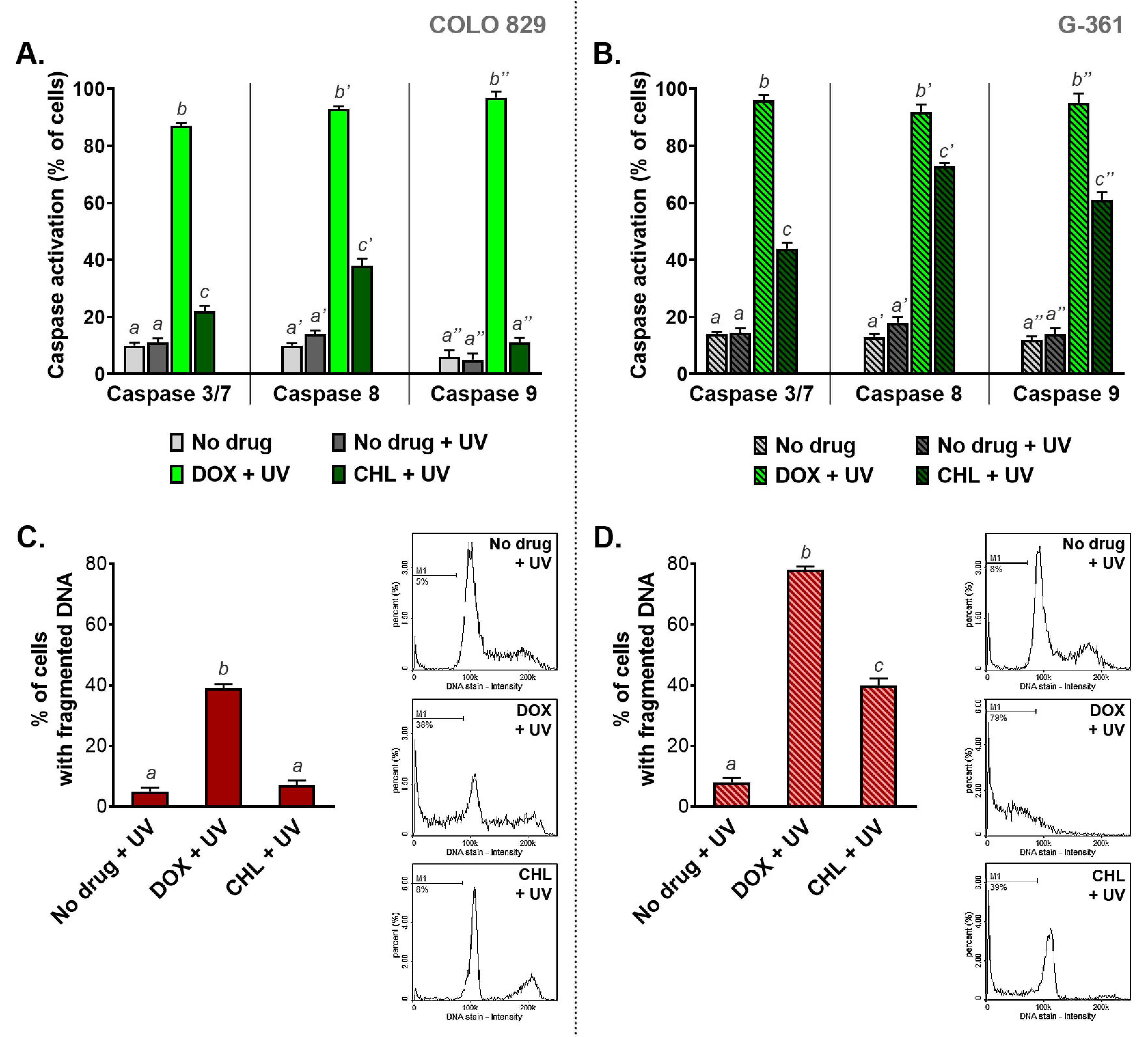
2.7. The Influence of Doxycycline and Chlortetracycline on Caspase Activation and DNA Fragmentation in Melanotic Melanoma Cells Exposed to UVA Radiation
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. Cell Viability Assessment
4.3. WST-1 Assay
4.4. The Quantitation of Cellular Reactive Oxygen Species
4.5. Assessment of the Intracellular Thiol Status
4.6. Annexin V Assay
4.7. Caspase Activity Assay
4.8. DNA Fragmentation Assay
4.9. Cell Cycle Analysis
4.10. Mitochondrial Potential Analysis
4.11. Immunofluorescence Analysis by Confocal Microscopy
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bertrand, J.U.; Steingrimsson, E.; Jouenne, F.; Bressac-de Paillerets, B.; Larue, L. Melanoma risk and melanocyte biology. Acta Derm-Venereol. 2020, 100, adv00139. [Google Scholar] [CrossRef]
- Raimondi, S.; Suppa, M.; Gandini, S. Melanoma epidemiology and sun exposure. Acta Derm-Venereol. 2020, 100, adv00136. [Google Scholar] [CrossRef]
- Respondek, M.; Beberok, A.; Rzepka, Z.; Rok, J.; Wrześniok, D. Mcl-1 inhibitor induces cells death in BRAF-mutant amelanotic melanoma trough GSH depletion, DNA damage and cell cycle changes. Pathol. Oncol. Res. 2020, 26, 1465–1474. [Google Scholar] [CrossRef]
- Respondek, M.; Beberok, A.; Rzepka, Z.; Rok, J.; Wrześniok, D. MIM1 induces COLO829 melanoma cell death through mitochondrial membrane breakdown, GSH depletion, and DNA damage. Fundam. Clin. Pharmacol. 2020, 34, 20–31. [Google Scholar] [CrossRef]
- Elder, D.E.; Bastian, B.C.; Cree, I.A.; Massi, D.; Scolyer, R.A. The 2018 world health organization classification of cutaneous, mucosal, and uveal melanoma: Detailed analysis of 9 district subtypes defined by their evolutionary pathway. Arch. Pathol. Lab. Med. 2020, 44, 500–522. [Google Scholar] [CrossRef]
- Weir, H.K.; Marrett, L.D.; Cokkinides, V.; Barnholtz-Sloan, J.; Patel, P.; Tai, E.; Jemal, A.; Li, J.; Kim, J.; Ekwueme, D.U. Melanoma in adolescents and young adults (ages 15–39 years): United States, 1999–2006. J. Am. Acad. Dermatol. 2011, 65, 38–49. [Google Scholar] [CrossRef]
- Diao, D.Y.; Lee, T.K. Sun-protective behaviors in populations at high risk for skin cancer. Psychol. Res. Behav. Manag. 2013, 7, 9–18. [Google Scholar] [CrossRef]
- Domingues, B.; Lopes, J.M.; Soares, P.; Pópulo, H. Melanoma treatment in review. Immunotargets Ther. 2018, 7, 35–49. [Google Scholar] [CrossRef]
- Lazaroff, J.; Bolotin, D. Targeted therapy and immunotherapy in melanoma. Dermatol. Clin. 2023, 41, 65–77. [Google Scholar] [CrossRef]
- Rok, J.; Karkoszka, M.; Rzepka, Z.; Respondek, M.; Banach, K.; Beberok, A.; Wrześniok, D. Cytotoxic and proapoptotic effect of doxycycline—An in vitro study on the human skin melanoma cells. Toxicol. Vitr. 2020, 65, 104790. [Google Scholar] [CrossRef]
- Nelson, M.L.; Levy, S.B. The history of the tetracyclines. Ann. N. Y. Acad. Sci. 2011, 1241, 17–32. [Google Scholar] [CrossRef]
- Barrenechea, V.; Vargas-Reyes, M.; Quiliano, M.; Milón, P. A complementary mechanism of bacterial mRNA translation inhibition by tetracyclines. Front. Microbiol. 2021, 12, 682682. [Google Scholar] [CrossRef]
- Singh, S.; Khanna, D.; Kalra, S. Minocycline and doxycycline: More than antibiotics. Curr. Mol. Pharmacol. 2021, 14, 1046–1065. [Google Scholar] [CrossRef]
- Barbie, D.A.; Kennedy, B.K. Doxycycline: New tricks for an old drug. Oncotarget 2015, 6, 19336–19337. [Google Scholar] [CrossRef]
- Esposito, S.; Bassetti, M.; Concia, E.; De Simone, G.; De Rosa, F.G.; Grossi, P.; Novelli, A.; Menichetti, F.; Petrosillo, N.; Tinelli, M.; et al. Italian Society of Infectious and Tropical Diseases, Diagnosis and management of skin and soft-tissue infections (SSTI). A literature review and consensus statement: An update. J. Chemother. 2017, 29, 197–214. [Google Scholar] [CrossRef]
- Reinbold, J.B.; Coetzee, J.F.; Gehring, R.; Havel, J.A.; Hollis, L.C.; Olson, K.C.; Apley, M.D. Plasma pharmacokinetics of oral chlortetracycline in group fed, ruminating, Holstein steers in a feedlot setting. J. Vet. Pharmacol. Therapeut. 2010, 33, 76–83. [Google Scholar] [CrossRef]
- Sun, T.; Zhao, N.; Ni, C.S.; Zhao, X.L.; Zhang, W.Z.; Su, X.; Zhang, D.F.; Gu, Q.; Sun, B.C. Doxycycline inhibits the adhesion and migration of melanoma cells by inhibiting the expression and phosphorylation of focal adhesion kinase (FAK). Cancer Lett. 2009, 28, 141–150. [Google Scholar] [CrossRef]
- Cortés, H.; Reyes-Hernández, O.D.; Alcalá-Alcalá, S.; Bernal-Chávez, S.A.; Caballero-Florán, I.H.; González-Torres, M.; SharifiRad, J.; González-Del Carmen, M.; Figueroa-González, G.; Leyva-Gómez, G. Repurposing of drug candidates for treatment of skin cancer. Front. Oncol. 2021, 10, 605714. [Google Scholar] [CrossRef]
- Rok, J.; Rzepka, Z.; Beberok, A.; Pawlik, J.; Wrześniok, D. Cellular and molecular aspects of anti-melanoma effect of minocycline—A study of cytotoxicity and apoptosis on human melanotic melanoma cells. Int. J. Mol. Sci. 2020, 21, 6917. [Google Scholar] [CrossRef]
- Trzcionka, J.; Buszman, E. Oddziaływanie antybiotyków tetracyklinowych z melaniną w aspekcie ich fototoksycznego działania na skórę. Ann. Acad. Med. Siles. 2003, 54–55, 55–63. [Google Scholar]
- Banning, T.P.; Heard, C.M. Binding of doxycycline to keratin, melanin and human epidermal tissue. Int. J. Pharm. 2002, 235, 219–227. [Google Scholar] [CrossRef]
- Bridelli, M.G.; Ciati, A.; Crippa, P.R. Binding of chemicals to melanins re-examined: Adsorption of some drugs to the surface of melanin particles. Biophys. Chem. 2006, 119, 137–145. [Google Scholar] [CrossRef]
- Rok, J.; Rzepka, Z.; Respondek, M.; Beberok, A.; Wrześniok, D. Chlortetracycline and melanin biopolymer—The risk of accumulation and implications for phototoxicity: An in vitro study on normal human melanocytes. Chem. Biol. Interact. 2019, 303, 27–34. [Google Scholar] [CrossRef]
- Rok, J.; Buszman, E.; Beberok, A.; Delijewski, M.; Otręba, M.; Wrześniok, D. Modulation of melanogenesis and antioxidant status of melanocytes in response to phototoxic action of doxycycline. Photochem. Photobiol. 2015, 91, 1429–1434. [Google Scholar] [CrossRef]
- Rok, J.; Buszman, E.; Delijewski, M.; Otrśba, M.; Beberok, A.; Wrześniok, D. Effect of tetracycline and UV radiation on melanization and antioxidant status of melanocytes. J. Photochem. Photobiol. B 2015, 148, 168–173. [Google Scholar] [CrossRef]
- Slominski, R.M.; Sarna, T.; Płonka, P.M.; Raman, C.; Brożyna, A.A.; Slominski, A.T. Melanoma, melanin, and melanogenesis: The yin and yang relationship. Front. Oncol. 2022, 12, 842496. [Google Scholar] [CrossRef]
- Drucker, A.M.; Rosen, C.F. Drug-induced photosensitivity: Culprit drugs, management and prevention. Drug Saf. 2011, 34, 821–837. [Google Scholar] [CrossRef]
- Nelson, M.L. Chemical and biological dynamics of tetracyclines. Adv. Dent. Res. 1998, 12, 5–11. [Google Scholar] [CrossRef]
- Quintero, B.; Miranda, M.A. Mechanisms of photosensitization induced by drugs: A general survey. Ars Pharm. 2000, 41, 27–46. [Google Scholar]
- Dolmans, D.E.; Fukumura, D.; Jain, R.K. Photodynamic therapy for cancer. Nat. Rev. Cancer. 2003, 3, 380–387. [Google Scholar] [CrossRef]
- Gunaydin, G.; Gedik, M.E.; Ayan, S. Photodynamic therapy for the treatment and diagnosis of cancer—A review of the current clinical status. Front. Chem. 2021, 9, 686303. [Google Scholar] [CrossRef]
- dos Santos, A.F.; de Almeida, D.R.Q.; Terra, L.F.; Baptista, M.S.; Labriola, L. Photodynamic therapy in cancer treatment—An update review. J. Cancer Metastasis. Treat. 2019, 5, 25. [Google Scholar] [CrossRef]
- Fuoco, D. Cytotoxicity induced by tetracyclines via protein photooxidation. Adv. Toxicol. 2015, 2015, 787129. [Google Scholar] [CrossRef]
- Sadiq, I.Z. Free radicals and oxidative stress: Signaling mechanisms, redox basis for human diseases, and cell cycle regulation. Curr. Mol. Med. 2023, 23, 13–35. [Google Scholar] [CrossRef]
- Arfin, S.; Jha, N.K.; Jha, S.K.; Kesari, K.K.; Ruokolainen, J.; Roychoudhury, S.; Rathi, B.; Kumar, D. Oxidative stress in cancer cell metabolism. Antioxidants 2021, 10, 642. [Google Scholar] [CrossRef]
- de Almeida, A.J.P.O.; de Oliveira, J.C.P.L.; da Silva Pontes, L.V.; de Souza Júnior, J.F.; Gonçalves, T.A.F.; Dantas, S.H.; de Almeida Feitosa, M.S.; Silva, A.O.; de Medeiros, I.A. ROS: Basic concepts, sources, cellular signaling, and its implications in aging pathways. Oxid. Med. Cell. Longev. 2022, 2022, 1225578. [Google Scholar] [CrossRef]
- Shackelford, R.E.; Kaufmann, W.K.; Paules, R.S. Oxidative stress and cell cycle checkpoint function. Free Radic. Biol. Med. 2000, 28, 1387–1404. [Google Scholar] [CrossRef]
- Kuczler, M.D.; Olseen, A.M.; Pienta, K.J.; Amend, S.R. ROS-induced cell cycle arrest as a mechanism of resistance in polyaneuploid cancer cells (PACCs). Prog. Biophys. Mol. Biol. 2021, 165, 3–7. [Google Scholar] [CrossRef]
- Noda, N.; Wakasugi, H. Cancer and oxidative stress. JMAJ 2001, 44, 535–539. [Google Scholar]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef]
- Hofmann, G.A.; Weber, B. Drug-induced photosensitivity: Culprit drugs, potential mechanisms and clinical consequences. J. Dtsch. Dermatol. Ges. 2021, 19, 19–29. [Google Scholar] [CrossRef]
- Kowalska, J.; Rok, J.; Rzepka, Z.; Wrześniok, D. Drug-induced photosensitivity-From light and chemistry to biological reactions and clinical symptoms. Pharmaceuticals 2021, 14, 723. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Narayanankutty, A.; Job, J.T.; Narayanankutty, V. Glutathione, an antioxidant tripeptide: Dual roles in carcinogenesis and chemoprevention. Curr. Protein Pept. Sci. 2019, 20, 907–917. [Google Scholar] [CrossRef]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsov, A.V.; Margreiter, R.; Ausserlechner, M.J.; Hagenbuchner, J. The complex interplay between mitochondria, ROS and entire cellular metabolism. Antioxidants 2022, 11, 1995. [Google Scholar] [CrossRef] [PubMed]
- Checa, J.; Aran, J.M. Reactive oxygen species: Drivers of physiological and pathological processes. J. Inflamm. Res. 2020, 13, 1057–1073. [Google Scholar] [CrossRef]
- Hao, X.; Bu, W.; Lv, G.; Xu, L.; Hou, D.; Wang, J.; Liu, X.; Yang, T.; Zhang, X.; Liu, Q.; et al. Disrupted mitochondrial homeostasis coupled with mitotic arrest generates antineoplastic oxidative stress. Oncogene 2022, 41, 427–443. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Reutelingsperger, C.P.; van Heerde, W.L. Annexin V, the regulator of phosphatidylserine-catalyzed inflammation and coagulation during apoptosis. Cell. Mol. Life Sci. 1997, 53, 527–532. [Google Scholar] [CrossRef] [PubMed]
- van Engeland, M.; Nieland, L.J.; Ramaekers, F.C.; Schutte, B.; Reutelingsperger, C.P. Annexin V-affinity assay: A review on an apoptosis detection system based on phosphatidylserine exposure. Cytometry 1998, 31, 1–9. [Google Scholar] [CrossRef]
- Berthenet, K.; Castillo Ferrer, C.; Fanfone, D.; Popgeorgiev, N.; Neves, D.; Bertolino, P.; Gibert, B.; Hernandez-Vargas, H.; Ichim, G. Failed apoptosis enhances melanoma cancer cell aggressiveness. Cell Rep. 2020, 31, 107731. [Google Scholar] [CrossRef] [PubMed]
- Jan, R.; Chaudhry, G.E. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Adv. Pharm. Bull. 2019, 9, 205–218. [Google Scholar] [CrossRef]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: Function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef]
- Li, X.Y.; Tan, L.C.; Dong, L.W.; Zhang, W.Q.; Shen, X.X.; Lu, X.; Zheng, H.; Lu, Y.G. Susceptibility and resistance mechanisms during photodynamic therapy of melanoma. Front. Oncol. 2020, 10, 597. [Google Scholar] [CrossRef]
- Yue, J.; Liang, L.; Shen, Y.; Guan, X.; Zhang, J.; Li, Z.; Deng, R.; Xu, S.; Liang, C.; Shi, W.; et al. Investigating dynamic molecular events in melanoma cell nucleus during photodynamic therapy by SERS. Front. Chem. 2019, 6, 665. [Google Scholar] [CrossRef]
- Saczko, J.; Kulbacka, J.; Chwiłkowska, A.; Drag-Zalesińiska, M.; Wysocka, T.; Lugowski, M.; Banaś, T. The influence of photodynamic therapy on apoptosis in human melanoma cell line. Folia Histochem. Cytobiol. 2005, 43, 129–132. [Google Scholar]
- Shi, T.; van Soest, D.M.K.; Polderman, P.E.; Burgering, B.M.T.; Dansen, T.B. DNA damage and oxidant stress activate p53 through differential upstream signaling pathways. Free Radic. Biol. Med. 2021, 172, 298–311. [Google Scholar] [CrossRef]
- Hussein, M.R.; Haemel, A.K.; Wood, G.S. Apoptosis and melanoma: Molecular mechanisms. J. Pathol. 2003, 199, 275–288. [Google Scholar] [CrossRef]
- Yamashita, T.; Tokino, T.; Tonoki, H.; Moriuchi, T.; Jin, H.Y.; Omori, F.; Jimbow, K. Induction of apoptosis in melanoma cell lines by p53 and its related proteins. J. Invest. Dermatol. 2001, 117, 914–919. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Bush, J.A.; Ho, V.C. p53-dependent apoptosis in melanoma cells after treatment with camptothecin. J. Invest. Dermatol. 2000, 114, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Box, N.F.; Vukmer, T.O.; Terzian, T. Targeting p53 in melanoma. Pigment Cell Melanoma Res. 2014, 27, 8–10. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Doxycycline | Chlortetracycline | ||||
|---|---|---|---|---|---|---|
| UV(−) | UV(+) | PIF* | UV(−) | UV(+) | PIF * | |
| COLO 829 | 51.1 µM | 15.0 µM | 3.4 | 185.2 µM | 52.0 µM | 3.6 |
| G-361 | 23.3 µM | 11.5 µM | 2.0 | 85.5 µM | 38.6 µM | 2.2 |
| Cell Line | Coefficient | Control | UV(+) | DOX UV(−) | DOX UV(+) | CHL UV(−) | CHL UV(+) |
|---|---|---|---|---|---|---|---|
| COLO 829 | G1/S | 4.3 | 6.2 | 17.0 | 1.8 | 16 | 12.2 |
| G2-M/S | 1.2 | 1.5 | 6.5 | 1.2 | 2.0 | 5.6 | |
| G-361 | G1/S | 4.3 | 6.5 | 15.4 | 4.0 | 4.6 | 16.5 |
| G2-M/S | 1.7 | 1.2 | 2.8 | 0.5 | 1.4 | 2.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rok, J.; Rzepka, Z.; Banach, K.; Kowalska, J.; Wrześniok, D. The Assessment of the Phototoxic Action of Chlortetracycline and Doxycycline as a Potential Treatment of Melanotic Melanoma—Biochemical and Molecular Studies on COLO 829 and G-361 Cell Lines. Int. J. Mol. Sci. 2023, 24, 2353. https://doi.org/10.3390/ijms24032353
Rok J, Rzepka Z, Banach K, Kowalska J, Wrześniok D. The Assessment of the Phototoxic Action of Chlortetracycline and Doxycycline as a Potential Treatment of Melanotic Melanoma—Biochemical and Molecular Studies on COLO 829 and G-361 Cell Lines. International Journal of Molecular Sciences. 2023; 24(3):2353. https://doi.org/10.3390/ijms24032353
Chicago/Turabian StyleRok, Jakub, Zuzanna Rzepka, Klaudia Banach, Justyna Kowalska, and Dorota Wrześniok. 2023. "The Assessment of the Phototoxic Action of Chlortetracycline and Doxycycline as a Potential Treatment of Melanotic Melanoma—Biochemical and Molecular Studies on COLO 829 and G-361 Cell Lines" International Journal of Molecular Sciences 24, no. 3: 2353. https://doi.org/10.3390/ijms24032353
APA StyleRok, J., Rzepka, Z., Banach, K., Kowalska, J., & Wrześniok, D. (2023). The Assessment of the Phototoxic Action of Chlortetracycline and Doxycycline as a Potential Treatment of Melanotic Melanoma—Biochemical and Molecular Studies on COLO 829 and G-361 Cell Lines. International Journal of Molecular Sciences, 24(3), 2353. https://doi.org/10.3390/ijms24032353