Determinants of Meal-Induced Changes in Circulating FFA Epoxides, Diols, and Diol-to-Epoxide Ratios as Indices of Soluble Epoxide Hydrolase Activity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
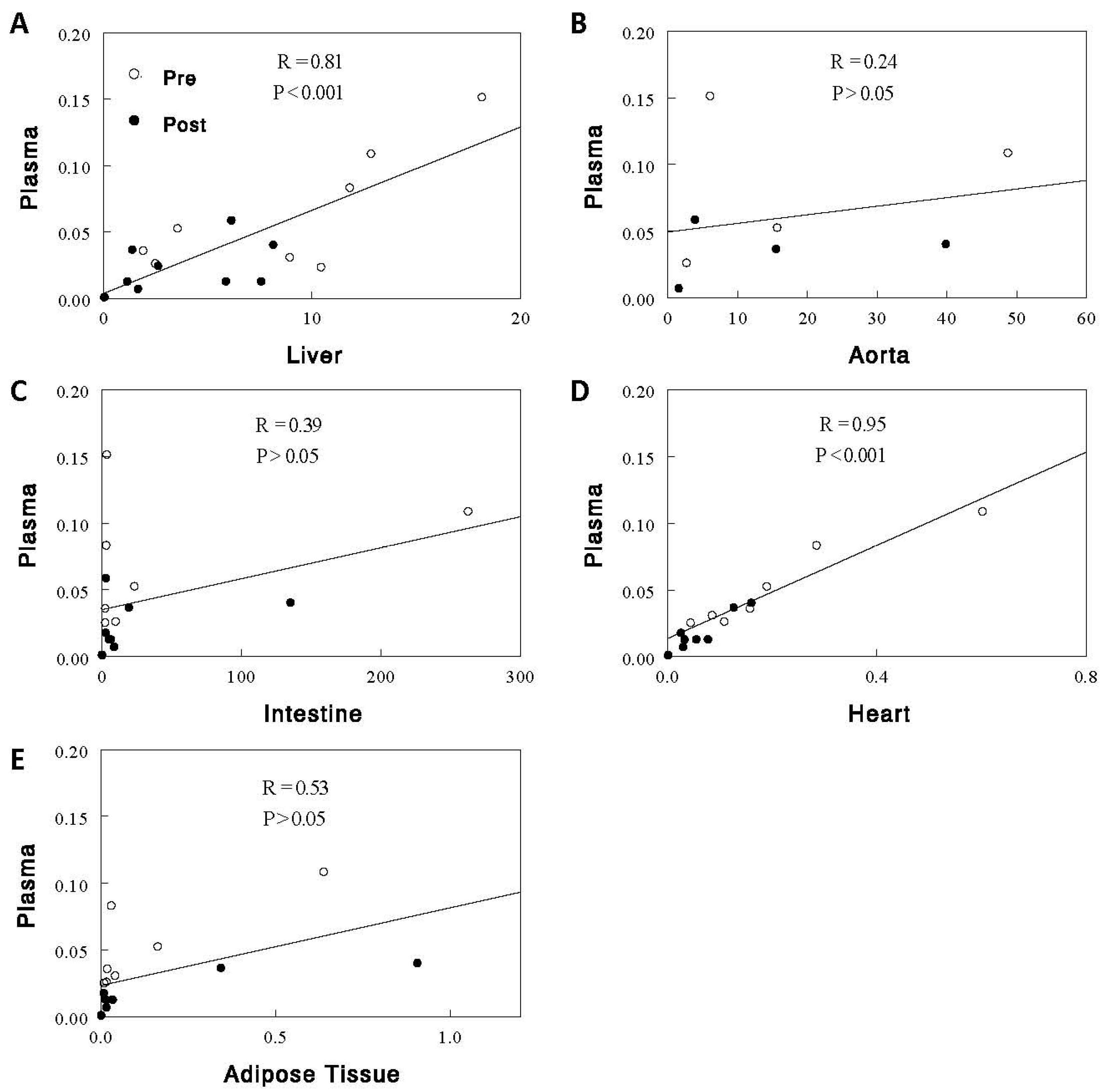
2.1. Effects of Acute Feeding on EpFAs, Diols, and D-to-E Ratios in Plasma and Individual Tissues
2.2. Effects of Acute Feeding on Renal Excretion of EpFAs and Diols
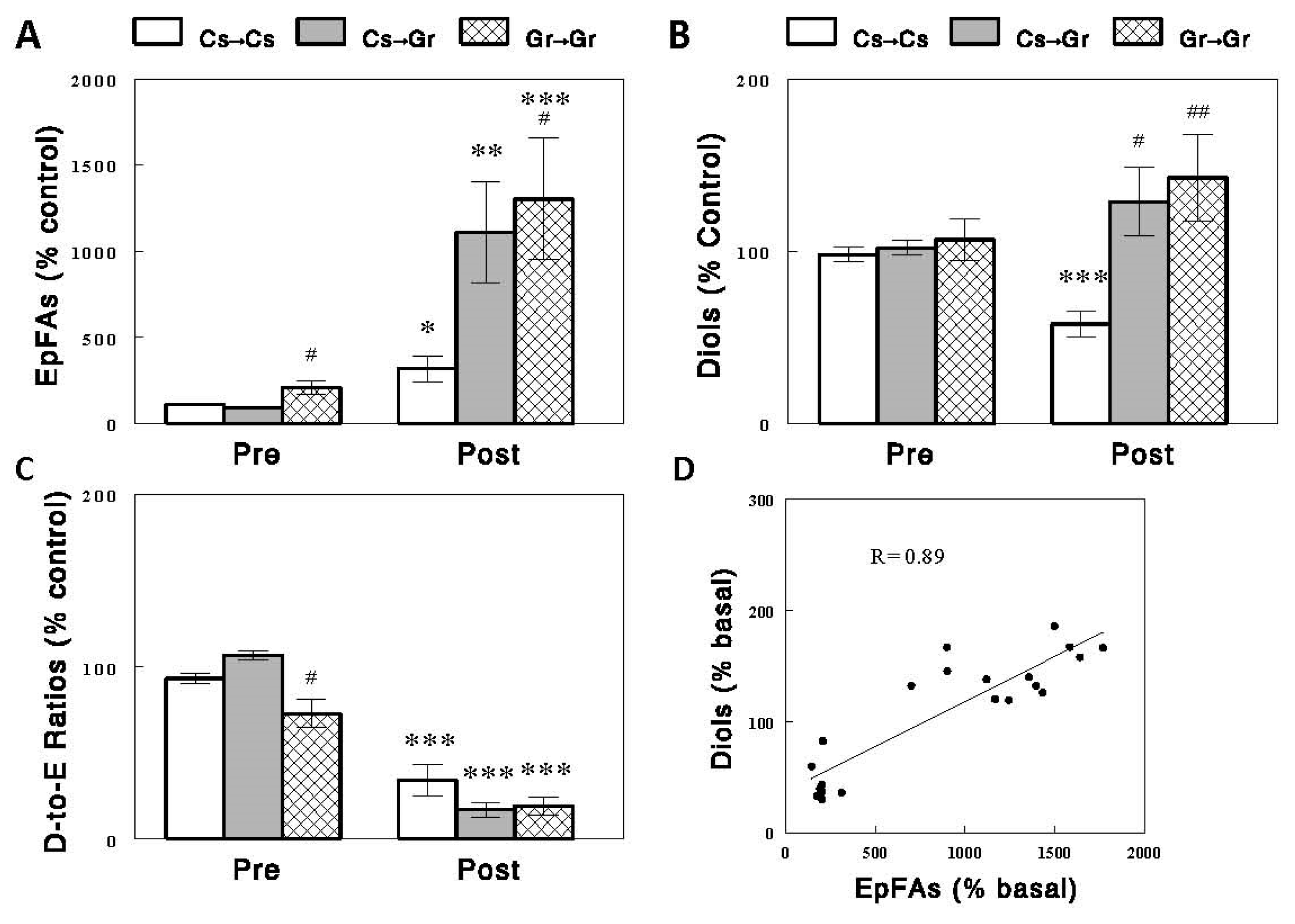
2.3. Effects of Diets on Circulating EpFAs, Diols, and D-to-E Ratios
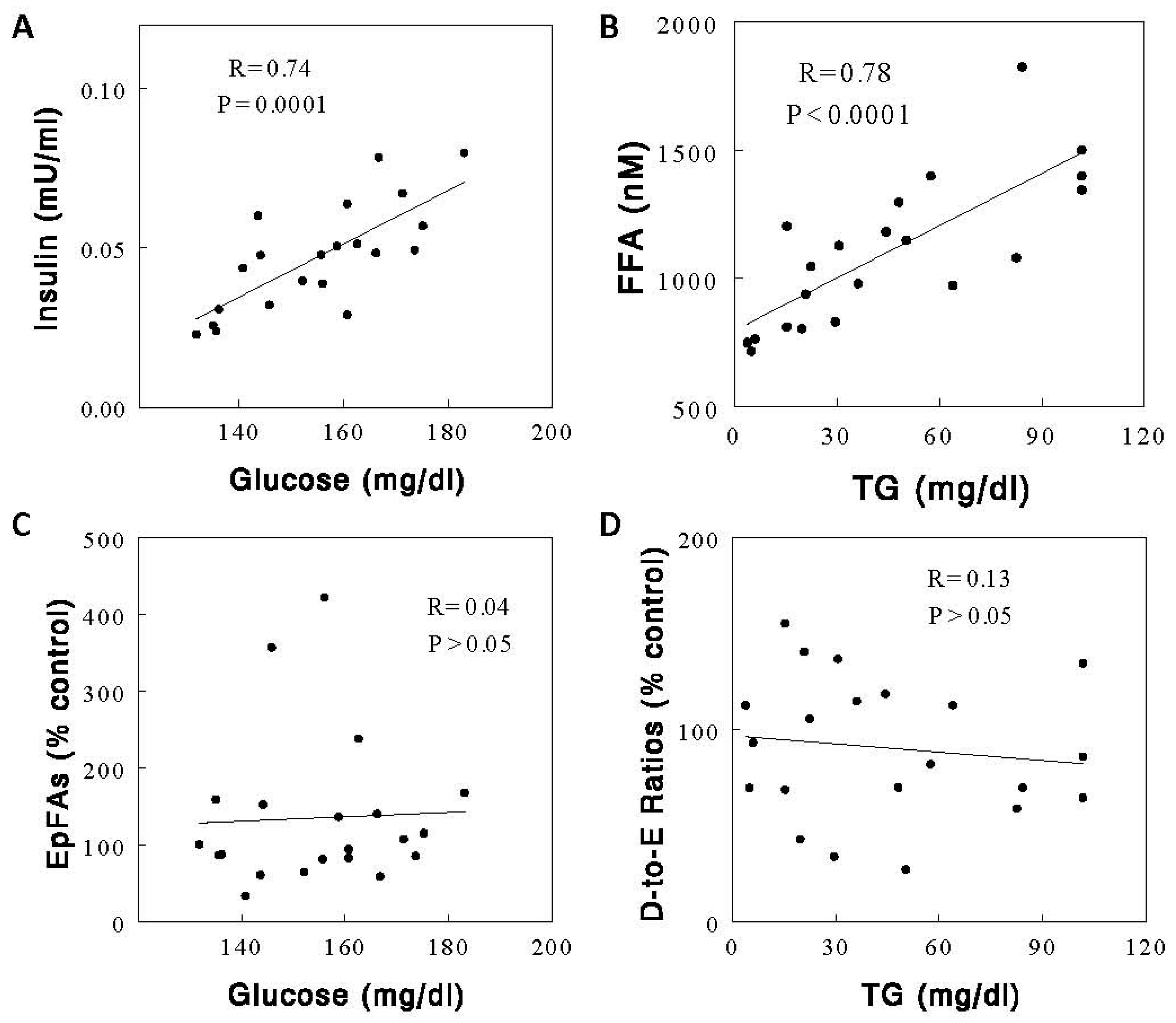
2.4. Effects of Diets on Circulating Metabolic Parameters
3. Discussion
4. Materials and Methods
4.1. Animals and Catheterization
4.2. Animal Experiments
4.3. Assays
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, H.; Weng, J.; Wang, M.H. EETs/sEH in diabetes and obesity-induced cardiovascular diseases. Prostaglandins Other Lipid Mediat 2016, 125, 80–89. [Google Scholar] [CrossRef]
- Edin, M.L.; Duval, C.; Zhang, G.; Zeldin, D.C. Role of linoleic acid-derived oxylipins in cancer. Cancer Metastasis Rev. 2020, 39, 581–582. [Google Scholar] [CrossRef]
- Mustafa, S.; Sharma, V.; McNeill, J.H. Insulin resistance and endothelial dysfunction: Are epoxyeicosatrienoic acids the link? Exp. Clin. Cardiol. 2009, 14, e41–e50. [Google Scholar]
- Xiao, Y.; Pietzner, A.; Rohwer, N.; Jung, A.; Rothe, M.; Weylandt, K.H.; Elbelt, U. Bioactive oxylipins in type 2 diabetes mellitus patients with and without hypertriglyceridemia. Front. Endocrinol. 2023, 14, 1195247. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, R.; Chen, G.; Hoopes, S.L.; Zeldin, D.C.; Wang, D.W. The Role of Cytochrome P450 Epoxygenases, Soluble Epoxide Hydrolase, and Epoxyeicosatrienoic Acids in Metabolic Diseases. Adv. Nutr. 2016, 7, 1122–1128. [Google Scholar] [CrossRef] [PubMed]
- Jurado-Fasoli, L.; Osuna-Prieto, F.J.; Yang, W.; Kohler, I.; Di, X.; Rensen, P.C.; Castillo, M.J.; Martinez-Tellez, B.; Amaro-Gahete, F.J. High omega-6/omega-3 fatty acid and oxylipin ratio in plasma is linked to an adverse cardiometabolic profile in middle-aged adults. J. Nutr. Biochem. 2023, 117, 109331. [Google Scholar] [CrossRef] [PubMed]
- Chiamvimonvat, N.; Ho, C.M.; Tsai, H.J.; Hammock, B.D. The soluble epoxide hydrolase as a pharmaceutical target for hypertension. J. Cardiovasc. Pharmacol. 2007, 50, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.R.; Pretorius, M.; Schuck, R.N.; Burch, L.H.; Bartlett, J.; Williams, S.M.; Zeldin, D.C.; Brown, N.J. Genetic variation in soluble epoxide hydrolase (EPHX2) is associated with forearm vasodilator responses in humans. Hypertension 2011, 57, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Dahl, M.; Grande, P.; Tybjaerg-Hansen, A.; Nordestgaard, B.G. Genetically reduced soluble epoxide hydrolase activity and risk of stroke and other cardiovascular disease. Stroke 2010, 41, 27–33. [Google Scholar] [CrossRef]
- Gschwendtner, A.; Ripke, S.; Freilinger, T.; Lichtner, P.; Müller-Myhsok, B.; Wichmann, H.E.; Meitinger, T.; Dichgans, M. Genetic variation in soluble epoxide hydrolase (EPHX2) is associated with an increased risk of ischemic stroke in white Europeans. Stroke 2008, 39, 1593–1596. [Google Scholar] [CrossRef]
- Fornage, M.; Lee, C.R.; Doris, P.A.; Bray, M.S.; Heiss, G.; Zeldin, D.C.; Boerwinkle, E. The soluble epoxide hydrolase gene harbors sequence variation associated with susceptibility to and protection from incident ischemic stroke. Hum. Mol. Genet. 2005, 14, 2829–2837. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ding, H.; Yan, J.; Hui, R.; Wang, W.; Kissling, G.E.; Zeldin, D.C.; Wang, D.W. Genetic variation in cytochrome P450 2J2 and soluble epoxide hydrolase and risk of ischemic stroke in a Chinese population. Pharmacogenet. Genom. 2008, 18, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Cheriyan, J.; Gutterman, D.D.; Mayer, R.J.; Ament, Z.; Griffin, J.L.; Lazaar, A.L.; Newby, D.E.; Tal-Singer, R.; Wilkinson, I.B. Mechanisms of Vascular Dysfunction in COPD and Effects of a Novel Soluble Epoxide Hydrolase Inhibitor in Smokers. Chest 2017, 151, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Hye Khan, M.A.; Hwang, S.H.; Sharma, A.; Corbett, J.A.; Hammock, B.D.; Imig, J.D. A dual COX-2/sEH inhibitor improves the metabolic profile and reduces kidney injury in Zucker diabetic fatty rat. Prostaglandins Other Lipid Mediat. 2016, 125, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Dreisbach, A.W.; Rice, J.C.; Japa, S.; Newman, J.W.; Sigel, A.; Gill, R.S.; Hess, A.E.; Cemo, A.C.; Fonseca, J.P.; Hammock, B.D.; et al. Salt loading increases urinary excretion of linoleic acid diols and triols in healthy human subjects. Hypertension 2008, 51, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Staprans, I.; Hardman, D.A.; Pan, X.M.; Feingold, K.R. Effect of oxidized lipids in the diet on oxidized lipid levels in postprandial serum chylomicrons of diabetic patients. Diabetes Care 1999, 22, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.P.; Yang, S.H.; Kim, D.K.; Lee, H.; Kim, B.; Cho, J.Y.; Yu, K.S.; Paik, J.H.; Kim, M.; Lim, C.S.; et al. In vivo activity of epoxide hydrolase according to sequence variation affects the progression of human IgA nephropathy. Am. J. Physiol. Renal Physiol. 2011, 300, F1283–F1290. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, D.; Dou, X.; Niu, N.; Huang, W.; Bai, J.; Zhang, G. Elevated 14,15-epoxyeicosatrienoic acid by increasing of cytochrome P450 2C8, 2C9 and 2J2 and decreasing of soluble epoxide hydrolase associated with aggressiveness of human breast cancer. BMC Cancer 2014, 14, 841. [Google Scholar] [CrossRef]
- Theken, K.N.; Schuck, R.N.; Edin, M.L.; Tran, B.; Ellis, K.; Bass, A.; Lih, F.B.; Tomer, K.B.; Poloyac, S.M.; Wu, M.C.; et al. Evaluation of cytochrome P450-derived eicosanoids in humans with stable atherosclerotic cardiovascular disease. Atherosclerosis 2012, 222, 530–536. [Google Scholar] [CrossRef]
- Kondo, K.; Morino, K.; Nishio, Y.; Kondo, M.; Nakao, K.; Nakagawa, F.; Ishikado, A.; Sekine, O.; Yoshizaki, T.; Kashiwagi, A.; et al. A fish-based diet intervention improves endothelial function in postmenopausal women with type 2 diabetes mellitus: A randomized crossover trial. Metabolism 2014, 63, 930–940. [Google Scholar] [CrossRef]
- Yang, J.; Oh, Y.T.; Wan, D.; Watanabe, R.M.; Hammock, B.D.; Youn, J.H. Postprandial effect to decrease soluble epoxide hydrolase activity: Roles of insulin and gut microbiota. J. Nutr. Biochem. 2017, 49, 8–14. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.T.; Yang, J.; Morisseau, C.; He, Q.; Hammock, B.; Youn, J.H. Effects of Individual Circulating FFAs on Plasma and Hepatic FFA Epoxides, Diols, and Epoxide-Diol Ratios as Indices of Soluble Epoxide Hydrolase Activity. Int. J. Mol. Sci. 2023, 24, 10760. [Google Scholar] [CrossRef] [PubMed]
- Braet, F.; Wisse, E. Structural and functional aspects of liver sinusoidal endothelial cell fenestrae: A review. Comp Hepatol. 2002, 1, 1. [Google Scholar] [CrossRef]
- Wu, Y.; Dong, J.H.; Dai, Y.F.; Zhu, M.Z.; Wang, M.Y.; Zhang, Y.; Pan, Y.D.; Yuan, X.R.; Guo, Z.X.; Wang, C.X.; et al. Hepatic soluble epoxide hydrolase activity regulates cerebral Aβ metabolism and the pathogenesis of Alzheimer’s disease in mice. Neuron 2023, 111, 2847–2862. [Google Scholar] [CrossRef]
- Dai, Y.; Dong, J.; Wu, Y.; Zhu, M.; Xiong, W.; Li, H.; Zhao, Y.; Hammock, B.D.; Zhu, X. Enhancement of the liver’s neuroprotective role ameliorates traumatic brain injury pathology. Proc. Natl. Acad. Sci. USA 2023, 120, e2301360120. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.A.; Knowles, R.B.; Kirkby, N.S.; Reed, D.M.; Edin, M.L.; White, W.E.; Chan, M.V.; Longhurst, H.; Yaqoob, M.M.; Milne, G.L.; et al. Kidney transplantation in a patient lacking cytosolic phospholipase A2 proves renal origins of urinary PGI-M and TX-M. Circ. Res. 2018, 122, 555–559. [Google Scholar] [CrossRef]
- Oh, K.S.; Oh, Y.T.; Kim, S.W.; Kita, T.; Kang, I.; Youn, J.H. Gut Sensing of Dietary K+ Intake Increases Renal K+ Excretion. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R421–R429. [Google Scholar] [CrossRef]
- Yang, J.; Schmelzer, K.; Georgi, K.; Hammock, B.D. Quantitative profiling method for oxylipin metabolome by liquid chromatography electrospray ionization tandem mass spectrometry. Anal. Chem. 2009, 81, 8085–8093. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, Y.T.; Yang, J.; Stefanovski, D.; Hammock, B.; Youn, J.H. Determinants of Meal-Induced Changes in Circulating FFA Epoxides, Diols, and Diol-to-Epoxide Ratios as Indices of Soluble Epoxide Hydrolase Activity. Int. J. Mol. Sci. 2023, 24, 17351. https://doi.org/10.3390/ijms242417351
Oh YT, Yang J, Stefanovski D, Hammock B, Youn JH. Determinants of Meal-Induced Changes in Circulating FFA Epoxides, Diols, and Diol-to-Epoxide Ratios as Indices of Soluble Epoxide Hydrolase Activity. International Journal of Molecular Sciences. 2023; 24(24):17351. https://doi.org/10.3390/ijms242417351
Chicago/Turabian StyleOh, Young Taek, Jun Yang, Darko Stefanovski, Bruce Hammock, and Jang H. Youn. 2023. "Determinants of Meal-Induced Changes in Circulating FFA Epoxides, Diols, and Diol-to-Epoxide Ratios as Indices of Soluble Epoxide Hydrolase Activity" International Journal of Molecular Sciences 24, no. 24: 17351. https://doi.org/10.3390/ijms242417351
APA StyleOh, Y. T., Yang, J., Stefanovski, D., Hammock, B., & Youn, J. H. (2023). Determinants of Meal-Induced Changes in Circulating FFA Epoxides, Diols, and Diol-to-Epoxide Ratios as Indices of Soluble Epoxide Hydrolase Activity. International Journal of Molecular Sciences, 24(24), 17351. https://doi.org/10.3390/ijms242417351