
Assessing the Ecotoxicity of Eight Widely Used Antibiotics on River Microbial Communities

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
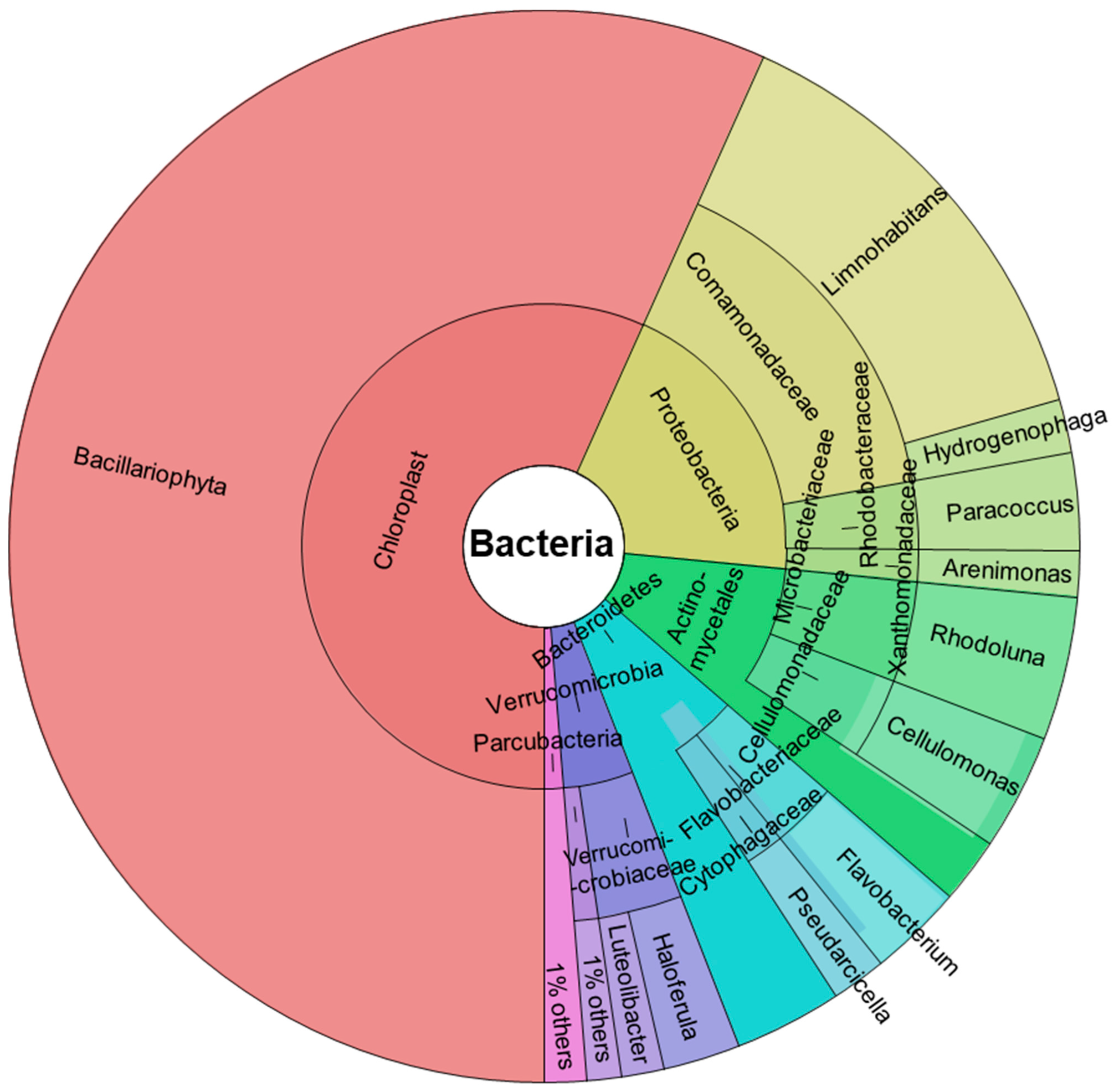
2.1. 16S rRNA Gene Sequencing of River Microbial Communities
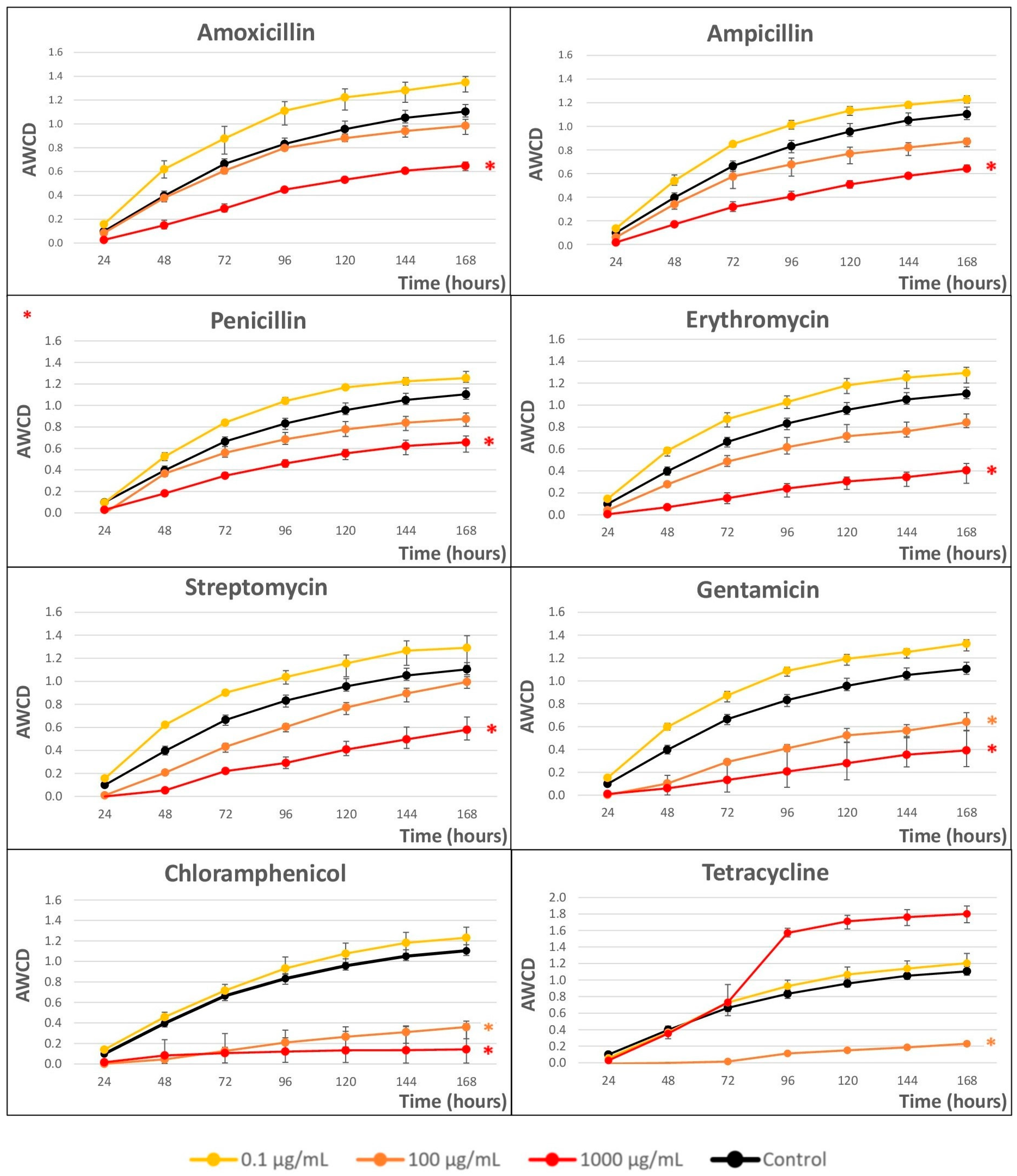
2.2. Impact of Antibiotics on the Growth of River Microbial Communities
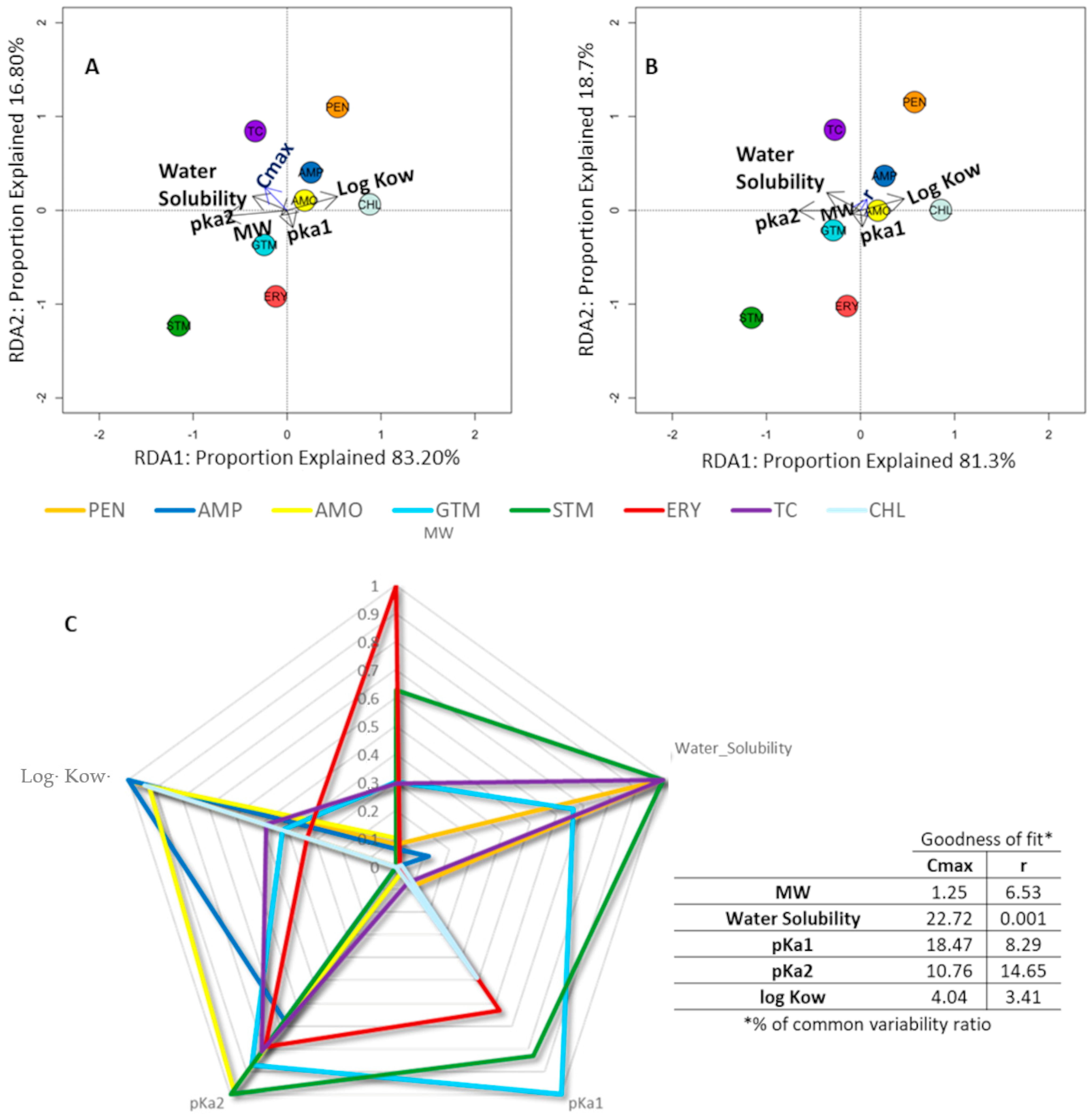
2.3. Relationship between the Physicochemical Properties of Antibiotics and the Impact on Bacterial Growth
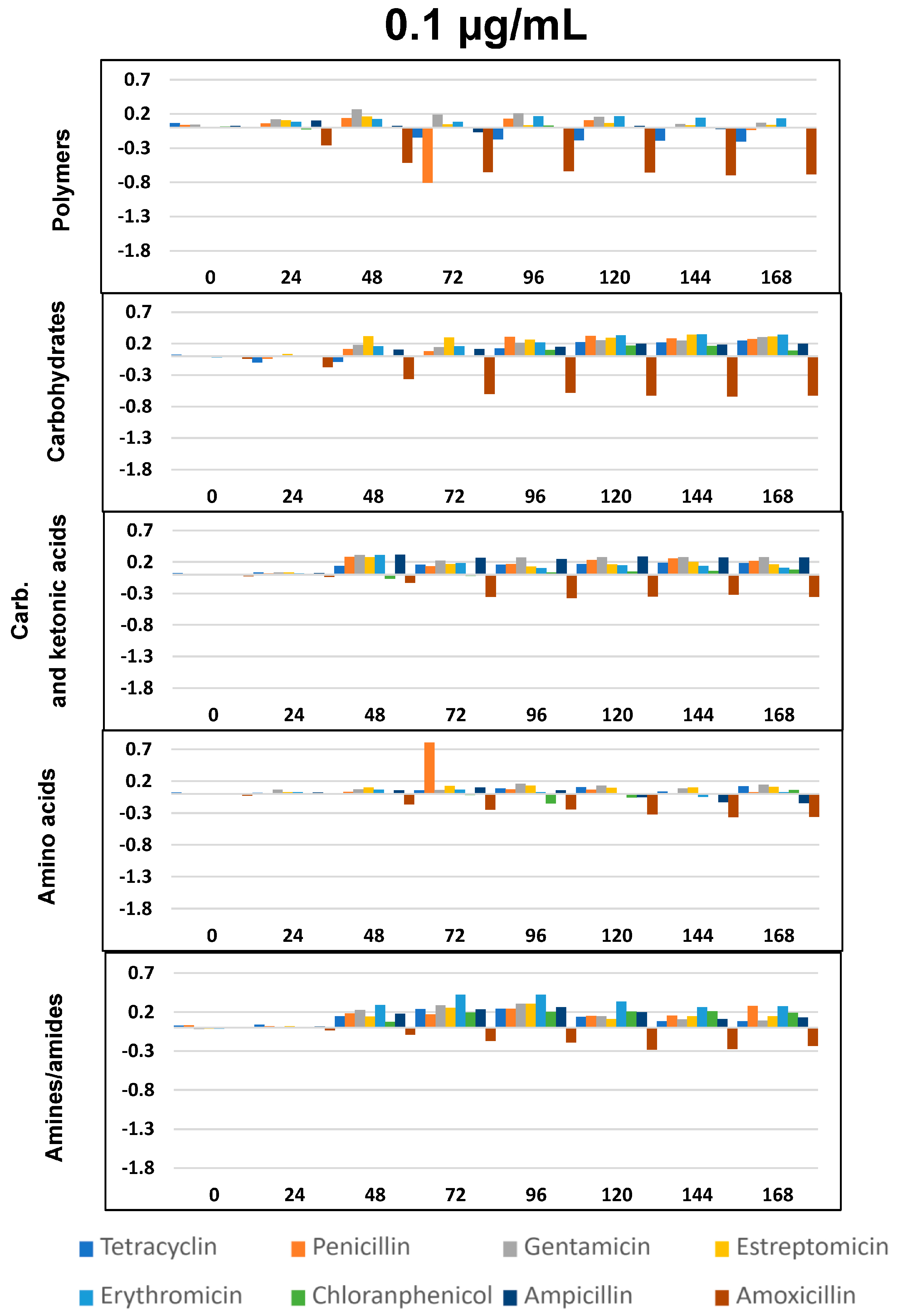
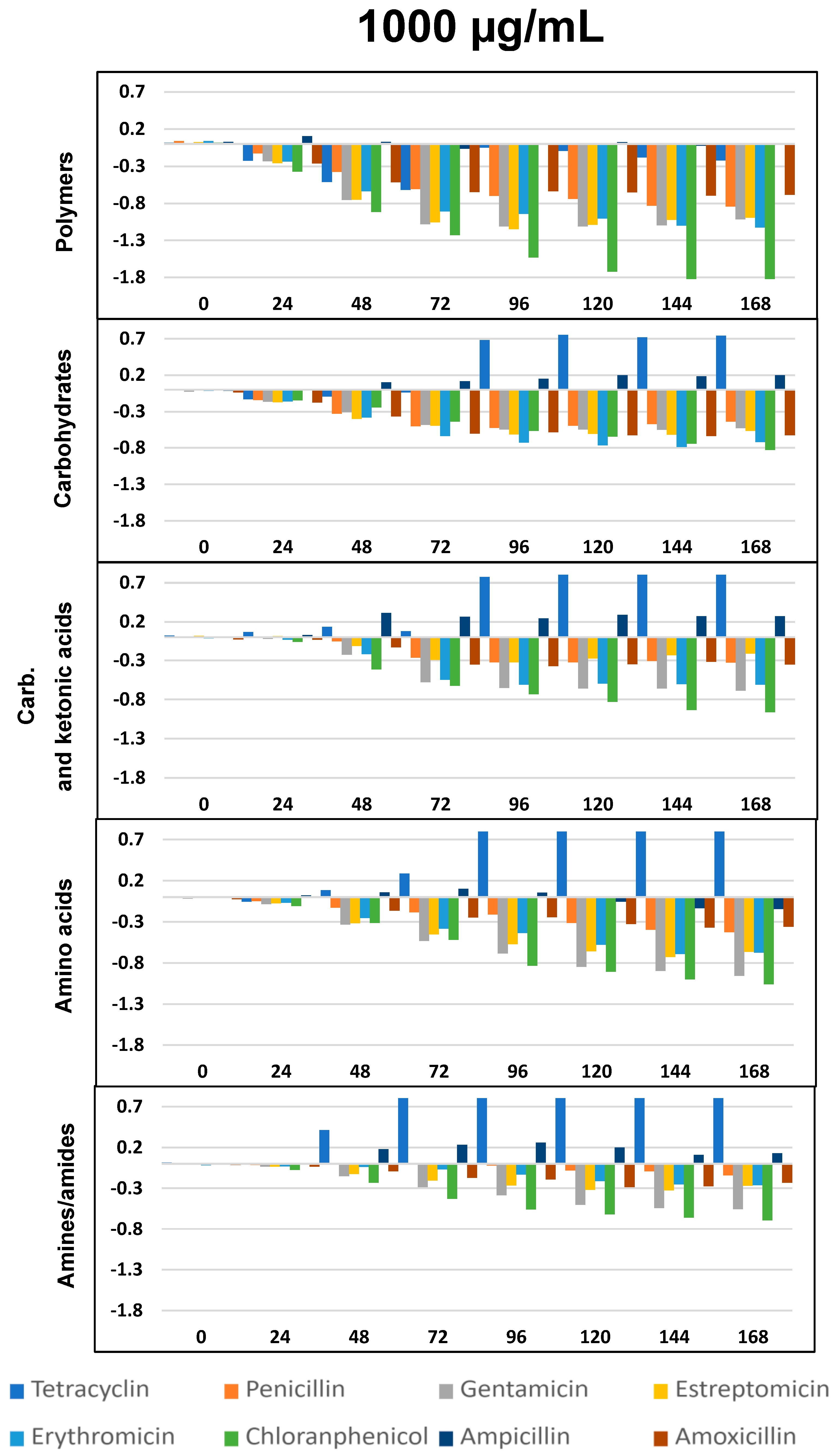
2.4. Impact of Antibiotics on Community-Level Physiological Profiling
3. Discussion
3.1. Chloramphenicol Is the Antibiotic with the Greatest Impact on River Microbiota
3.2. Macrolides and Aminoglycosides Have a Great Impact on Microbial Growth
3.3. Beta-Lactams Have a Minor Effect on Microbial Growth Kinetics
3.4. Tetracycline Has an Anomalous Behavior
3.5. Environmental Relevance
4. Materials and Methods
4.1. Antibiotics
4.2. River Microorganism Sampling
4.3. River Sample Preparation for Genetic Analysis
4.4. River Sample Preparation for Community-Level Physiological Profiling of Freshwater Microbes with Biolog EcoPlates™
4.5. Taxonomical Analysis: 16S rRNA Gene Sequencing
4.6. Bioinformatic Analysis
4.7. Data Representation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cycoń, M.; Mrozik, A.; Piotrowska-Seget, Z. Antibiotics in the Soil Environment—Degradation and Their Impact on Microbial Activity and Diversity. Front. Microbiol. 2019, 10, 338. [Google Scholar] [CrossRef] [PubMed]
- Klein, E.Y.; Van Boeckel, T.P.; Martinez, E.M.; Pant, S.; Gandra, S.; Levin, S.A.; Goossens, H.; Laxminarayan, R. Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc. Natl. Acad. Sci. USA 2018, 115, E3463–E3470. [Google Scholar] [CrossRef] [PubMed]
- Carballa, M.; Omil, F.; Lema, J.M.; Llompart, M.; GarcÍa-Jares, C.; RodrÍguez, I.; Gómez, M.; Ternes, T. Behavior of pharmaceuticals, cosmetics and hormones in a sewage treatment plant. Water Res. 2004, 38, 2918–2926. [Google Scholar] [CrossRef] [PubMed]
- Omuferen, L.O.; Maseko, B.; Olowoyo, J.O. Occurrence of antibiotics in wastewater from hospital and convectional wastewater treatment plants and their impact on the effluent receiving rivers: Current knowledge between 2010 and 2019. Environ. Monit. Assess. 2022, 194, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, Z. Occurrences of pharmaceutical and personal care products as micropollutants in rivers from Romania. Chemosphere 2006, 64, 1808–1817. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, N.E.; Savun-Hekimoğlu, B.; Aksu, A.; Burak, S.; Caglar, N.B. Occurrence, sources and environmental risk assessment of pharmaceuticals in the Sea of Marmara, Turkey. Sci. Total Environ. 2022, 819, 152996. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Ji, M.; Zhai, H.; Guo, Y.; Liu, Y. Occurrence of antibiotics and antibiotic resistance genes in WWTP effluent-receiving water bodies and reclaimed wastewater treatment plants. Sci. Total Environ. 2021, 796, 148919. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Wu, L.; Huang, Y.; Luo, Y.; Christie, P. Total concentrations of heavy metals and occurrence of antibiotics in sewage sludges from cities throughout China. J. Soils Sediments 2014, 14, 1123–1135. [Google Scholar] [CrossRef]
- Chen, C.X.; Aris, A.; Yong, E.L.; Noor, Z.Z. Evaluation of the occurrence of antibiotics at different treatment stages of decentralised and conventional sewage treatment plants. Int. J. Environ. Sci. Technol. 2022, 19, 5547–5562. [Google Scholar] [CrossRef]
- Larsson, D.J.; de Pedro, C.; Paxeus, N. Effluent from drug manufactures contains extremely high levels of pharmaceuticals. J. Hazard. Mater. 2007, 148, 751–755. [Google Scholar] [CrossRef]
- Al-Riyami, I.M.; Ahmed, M.; Al-Busaidi, A.; Choudri, B.S. Antibiotics in wastewaters: A review with focus on Oman. Appl. Water Sci. 2018, 8, 199. [Google Scholar] [CrossRef]
- Bhagat, C.; Kumar, M.; Tyagi, V.K.; Mohapatra, P.K. Proclivities for prevalence and treatment of antibiotics in the ambient water: A review. Npj Clean Water 2020, 3, 42. [Google Scholar] [CrossRef]
- Grenni, P.; Ancona, V.; Caracciolo, A.B. Ecological effects of antibiotics on natural ecosystems: A review. Microchem. J. 2018, 136, 25–39. [Google Scholar] [CrossRef]
- Liu, X.H.; Zhang, G.D.; Liu, Y.; Lu, S.Y.; Qin, P.; Guo, X.C.; Bi, B.; Wang, L.; Xi, B.D.; Wu, F.C.; et al. Occurrence and fate of antibiotics and antibiotic resistance genes in typical urban water of Beijing, China. Environ. Pollut. 2018, 246, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhang, D.; Xiao, S.; Geng, C.; Zhang, X. Occurrence and sources of antibiotics and their metabolites in river water, WWTPs, and swine wastewater in Jiulongjiang River basin, south China. Environ. Sci. Pollut. Res. 2013, 20, 9075–9083. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhu, Z.; Zhao, M.; He, J.; Zhang, X.; Hao, F.; Du, P. Occurrence, removal, emission and environment risk of 32 antibiotics and metabolites in wastewater treatment plants in Wuhu, China. Sci. Total Environ. 2023, 899, 165681. [Google Scholar] [CrossRef] [PubMed]
- Kulik, K.; Lenart-Boroń, A.; Wyrzykowska, K. Impact of Antibiotic Pollution on the Bacterial Population within Surface Water with Special Focus on Mountain Rivers. Water 2023, 15, 975. [Google Scholar] [CrossRef]
- Reis, A.C.; Kolvenbach, B.A.; Nunes, O.C.; Corvini, P.F. Biodegradation of antibiotics: The new resistance determinants–part I. New Biotechnol. 2019, 54, 34–51. [Google Scholar] [CrossRef]
- Yang, Q.; Gao, Y.; Ke, J.; Show, P.L.; Ge, Y.; Liu, Y.; Guo, R.; Chen, J. Antibiotics: An overview on the environmental occurrence, toxicity, degradation, and removal methods. Bioengineered 2021, 12, 7376–7416. [Google Scholar] [CrossRef]
- Nomadolo, N.; Dada, O.E.; Swanepoel, A.; Mokhena, T.; Muniyasamy, S. A Comparative Study on the Aerobic Biodegradation of the Biopolymer Blends of Poly(butylene succinate), Poly(butylene adipate terephthalate) and Poly(lactic acid). Polymers 2022, 14, 1894. [Google Scholar] [CrossRef]
- Olivito, F.; Jagdale, P.; Oza, G. Synthesis and Biodegradation Test of a New Polyether Polyurethane Foam Produced from PEG 400, L-Lysine Ethyl Ester Diisocyanate (L-LDI) and Bis-hydroxymethyl Furan (BHMF). Toxics 2023, 11, 698. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lv, K.; Deng, C.; Yu, Z.; Shi, J.; Johnson, A.C. Persistence and migration of tetracycline, sulfonamide, fluoroquinolone, and macrolide antibiotics in streams using a simulated hydrodynamic system. Environ. Pollut. 2019, 252, 1532–1538. [Google Scholar] [CrossRef] [PubMed]
- Elizalde-Velazquez, A.; Martinez-Rodriguez, H.; Galar-Martinez, M.; Dublan-Garcia, O.; Islas-Flores, H.; Rodriguez-Flores, J.; Castañeda-Peñalvo, G.; Lizcano-Sanz, I.; Gómez-Oliván, L.M. Effect of amoxicillin exposure on brain, gill, liver, and kidney of common carp (Cyprinus carpio): The role of amoxicilloic acid. Environ. Toxicol. 2017, 32, 1102–1120. [Google Scholar] [CrossRef] [PubMed]
- Sendra, M.; Moreno-Garrido, I.; Blasco, J.; Araújo, C.V. Effect of erythromycin and modulating effect of CeO2 NPs on the toxicity exerted by the antibiotic on the microalgae Chlamydomonas reinhardtii and Phaeodactylum tricornutum. Environ. Pollut. 2018, 242, 357–366. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Q.; Zhang, J.; Dong, J.; Ao, Y.; Wang, M.; Wang, X. Long-term exposure to antibiotic mixtures favors microcystin synthesis and release in Microcystis aeruginosa with different morphologies. Chemosphere 2019, 235, 344–353. [Google Scholar] [CrossRef]
- Seoane, M.; Rioboo, C.; Herrero, C.; Cid, A. Toxicity induced by three antibiotics commonly used in aquaculture on the marine microalga Tetraselmis suecica (Kylin) Butch. Mar. Environ. Res. 2014, 101, 1–7. [Google Scholar] [CrossRef]
- Lee, S.; Kim, C.; Liu, X.; Lee, S.; Kho, Y.; Kim, W.-K.; Kim, P.; Choi, K. Ecological Risk Assessment of Amoxicillin, Enrofloxacin, and Neomycin: Are Their Current Levels in the Freshwater Environment Safe? Toxics 2021, 9, 196. [Google Scholar] [CrossRef]
- Ferreira, C.S.G.; Nunes, B.A.; Henriques-Almeida, J.M.d.M.; Guilhermino, L. Acute toxicity of oxytetracycline and florfenicol to the microalgae Tetraselmis chuii and to the crustacean Artemia parthenogenetica. Ecotoxicol. Environ. Saf. 2007, 67, 452–458. [Google Scholar] [CrossRef]
- Isidori, M.; Lavorgna, M.; Nardelli, A.; Pascarella, L.; Parrella, A. Toxic and genotoxic evaluation of six antibiotics on non-target organisms. Sci. Total Environ. 2005, 346, 87–98. [Google Scholar] [CrossRef]
- Yang, C.; Song, G.; Lim, W. A review of the toxicity in fish exposed to antibiotics. Comp. Biochem. Physiol. C-Toxicol. Pharmacol. 2020, 237, 108840. [Google Scholar] [CrossRef]
- Huang, H.; Liu, Z.; Yang, H.; Yin, D. Acute and Chronic Toxic Effects of Chloramphenicol, Erythromycin and Tetracycline on the Early Life Stage of Amphibian (Xenopus tropicalis). Asian J. Chem. 2013, 25, 10293–10295. [Google Scholar] [CrossRef]
- Baumann, M.; Weiss, K.; Maletzki, D.; Schüssler, W.; Schudoma, D.; Kopf, W.; Kühnen, U. Aquatic toxicity of the macrolide antibiotic clarithromycin and its metabolites. Chemosphere 2015, 120, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Minguez, L.; Pedelucq, J.; Farcy, E.; Ballandonne, C.; Budzinski, H.; Halm-Lemeille, M.-P. Toxicities of 48 pharmaceuticals and their freshwater and marine environmental assessment in northwestern France. Environ. Sci. Pollut. Res. 2016, 23, 4992–5001. [Google Scholar] [CrossRef] [PubMed]
- Bielen, A.; Šimatović, A.; Kosić-Vukšić, J.; Senta, I.; Ahel, M.; Babić, S.; Jurina, T.; Plaza, J.J.G.; Milaković, M.; Udiković-Kolić, N. Negative environmental impacts of antibiotic-contaminated effluents from pharmaceutical industries. Water Res. 2017, 126, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Kovalakova, P.; Cizmas, L.; McDonald, T.J.; Marsalek, B.; Feng, M.B.; Sharma, V.K. Occurrence and toxicity of antibiotics in the aquatic environment: A review. Chemosphere 2020, 251, 126351. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, P.; Wang, C.; Chen, J.; Wang, X.; Hu, B.; Yuan, Q. Ecological insights into the disturbances in bacterioplankton communities due to emerging organic pollutants from different anthropogenic activities along an urban river. Sci. Total Environ. 2021, 796, 148973. [Google Scholar] [CrossRef] [PubMed]
- Schoffelen, N.J.; Mohr, W.; Ferdelman, T.G.; Duerschlag, J.; Littmann, S.; Ploug, H.; Kuypers, M.M.M. Phosphate availability affects fixed nitrogen transfer from diazotrophs to their epibionts. Isme J. 2019, 13, 2701–2713. [Google Scholar] [CrossRef]
- Wang, Z.; Han, S.; Cai, M.; Du, P.; Zhang, Z.; Li, X. Environmental behavior of methamphetamine and ketamine in aquatic ecosystem: Degradation, bioaccumulation, distribution, and associated shift in toxicity and bacterial community. Water Res. 2020, 174, 115585. [Google Scholar] [CrossRef]
- Zhang, M.; Sun, Q.; Chen, P.; Wei, X.; Wang, B. How microorganisms tell the truth of potentially toxic elements pollution in environment. J. Hazard. Mater. 2022, 431, 128456. [Google Scholar] [CrossRef]
- Christensen, A.M.; Ingerslev, F.; Baun, A. Ecotoxicity of mixtures of antibiotics used in aquacultures. Environ. Toxicol. Chem. 2006, 25, 2208–2215. [Google Scholar] [CrossRef]
- Kergoat, L.; Besse-Hoggan, P.; Leremboure, M.; Beguet, J.; Devers, M.; Martin-Laurent, F.; Masson, M.; Morin, S.; Roinat, A.; Pesce, S.; et al. Environmental Concentrations of Sulfonamides Can Alter Bacterial Structure and Induce Diatom Deformities in Freshwater Biofilm Communities. Front. Microbiol. 2021, 12, 643719. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Debognies, A.; Zhang, Q.; Zhang, Z.Y.; Zhou, Z.G.; Zhang, J.F.; Sun, L.W.; Lu, T.; Qian, H.F. Effects of ofloxacin on the structure and function of freshwater microbial communities. Aquat. Toxicol. 2022, 244, 106084. [Google Scholar] [CrossRef] [PubMed]
- Maul, J.D.; Schuler, L.J.; Belden, J.B.; Whiles, M.R.; Lydy, M.J. Effects of the antibiotic ciprofloxacin on stream microbial communities and detritivorous macroinvertebrates. Environ. Toxicol. Chem. 2006, 25, 1598–1606. [Google Scholar] [CrossRef] [PubMed]
- Zak, J.C.; Willig, M.R.; Moorhead, D.L.; Wildman, H.G. Functional diversity of microbial communities: A quantitative approach. Soil Biol. Biochem. 1994, 26, 1101–1108. [Google Scholar] [CrossRef]
- Freixa, A.; Romaní, A.M. Shifts in carbon substrate utilization in sediment microbial communities along the Llobregat River. Fundam. Appl. Limnol. 2014, 185, 247–261. [Google Scholar] [CrossRef]
- Gendy, S.; Chauhan, A.; Agarwal, M.; Pathak, A.; Rathore, R.S.; Jaswal, R. Is Long-Term Heavy Metal Exposure Driving Carriage of Antibiotic Resistance in Environmental Opportunistic Pathogens: A Comprehensive Phenomic and Genomic Assessment UsingSerratiasp. SRS-8-S-2018. Front. Microbiol. 2020, 11, 1923. [Google Scholar] [CrossRef] [PubMed]
- Pino-Otín, M.R.; Ballestero, D.; Navarro, E.; González-Coloma, A.; Val, J.; Mainar, A.M. Ecotoxicity of a novel biopesticide from Artemisia absinthium on non-target aquatic organisms. Chemosphere 2019, 216, 131–146. [Google Scholar] [CrossRef]
- Pino-Otín, M.R.; Muñiz, S.; Val, J.; Navarro, E. Effects of 18 pharmaceuticals on the physiological diversity of edaphic microorganisms. Sci. Total Environ. 2017, 595, 441–450. [Google Scholar] [CrossRef]
- Battistuzzi, F.U.; Hedges, S.B. A Major Clade of Prokaryotes with Ancient Adaptations to Life on Land. Mol. Biol. Evol. 2009, 26, 335–343. [Google Scholar] [CrossRef]
- Kasalický, V.; Jezbera, J.; Hahn, M.W.; Šimek, K. The Diversity of the Limnohabitans Genus, an Important Group of Freshwater Bacterioplankton, by Characterization of 35 Isolated Strains. PLoS ONE 2013, 8, e58209. [Google Scholar] [CrossRef]
- Zhang, M.; Zeng, G.; Liang, D.; Xu, Y.; Li, Y.; Huang, X.; Ma, Y.; Wang, F.; Liao, C.; Tang, C.; et al. An Analysis of the Colony Structure of Prokaryotes in the Jialing River Waters in Chongqing. Int. J. Environ. Res. Public Health 2022, 19, 5525. [Google Scholar] [CrossRef] [PubMed]
- Willey, J.M.; Sherwood, L.M.; Woolverton, C.J. Prescott’s Microbiology, 9th ed.; McGraw Hill Higher Education: London, UK, 2013. [Google Scholar]
- Kim, A.-R.; Lee, S.; Han, K.; Ahn, T.-Y. Arenimonas aquaticum sp. nov., a member of the gammaproteobacterium, isolated from a freshwater reservoir. J. Microbiol. 2013, 51, 887. [Google Scholar] [CrossRef]
- Pitt, A.; Schmidt, J.; Koll, U.; Hahn, M.W. Rhodoluna limnophila sp. nov., a bacterium with 1.4 Mbp genome size isolated from freshwater habitats located in Salzburg, Austria. Int. J. Syst. Evol. Microbiol. 2019, 69, 3946–3954. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, H.; Hayashi, T.; Hamada, M.; Kohda, T.; Serisawa, Y.; Matsuyama-Serisawa, K.; Nakagawa, Y.; Otoguro, M.; Yanagida, F.; Tamura, T.; et al. Cellulomonas algicola sp. nov., an actinobacterium isolated from a freshwater alga. Int. J. Syst. Evol. Microbiol. 2019, 69, 2723–2728. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Jeon, C.O. Flavobacterium alvei sp. nov., isolated from a freshwater river. Int. J. Syst. Evol. Microbiol. 2018, 68, 1919–1924. [Google Scholar] [CrossRef] [PubMed]
- Cruaud, P.; Vigneron, A.; Dorea, C.C.; Rodriguez, M.J.; Charette, S.J. Rapid Changes in Microbial Community Structures along a Meandering River. Microorganisms 2020, 8, 1631. [Google Scholar] [CrossRef] [PubMed]
- Fuerst, J.A. Phylum Verrucomicrobia. In Encyclopedia of Microbiology, 4th ed.; Schmidt, T.M., Ed.; Academic Press: Amsterdam, The Netherlands, 2019; pp. 551–563. ISBN 9780128117378. [Google Scholar] [CrossRef]
- Lee, K.-C.; Webb, R.I.; Janssen, P.H.; Sangwan, P.; Romeo, T.; Staley, J.T.; Fuerst, J.A. Phylum Verrucomicrobia representatives share a compartmentalized cell plan with members of bacterial phylum Planctomycetes. BMC Microbiol. 2009, 9, 5. [Google Scholar] [CrossRef]
- Weber, K.P.; Legge, R.L. One-dimensional metric for tracking bacterial community divergence using sole carbon source utilization patterns. J. Microbiol. Methods 2009, 79, 55–61. [Google Scholar] [CrossRef]
- Lehman, R.M.; Colwell, F.S.; Ringelberg, D.B.; White, D.C. Combined microbial community-level analyses for quality assurance of terrestrial subsurface cores. J. Microbiol. Methods 1995, 22, 263–281. [Google Scholar] [CrossRef]
- Wang, X.; Gu, J.; Gao, H.; Qian, X.; Li, H. Abundances of Clinically Relevant Antibiotic Resistance Genes and Bacterial Community Diversity in the Weihe River, China. Int. J. Environ. Res. Public Health 2018, 15, 708. [Google Scholar] [CrossRef]
- Narciso-Da-Rocha, C.; Manaia, C.M. Multidrug resistance phenotypes are widespread over different bacterial taxonomic groups thriving in surface water. Sci. Total Environ. 2016, 563–564, 1–9. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, M.J. (Ed.) The Merck Index—An Encyclopedia of Chemicals, Drugs, and Biologicals; Royal Society of Chemistry: Cambridge, UK, 2013; p. 367. [Google Scholar]
- Alves, J.; Dias, L.; Mateus, J.; Marques, J.; Graças, D.; Ramos, R.; Seldin, L.; Henriques, I.; Silva, A.; Folador, A. Resistome in Lake Bolonha, Brazilian Amazon: Identification of Genes Related to Resistance to Broad-Spectrum Antibiotics. Front. Microbiol. 2020, 11, 67. [Google Scholar] [CrossRef] [PubMed]
- Skopnik, H.; Heimann, G. Pharmacokinetics of antimicrobial drugs in the cerebrospinal fluid. Pediatr. Pharmacol. 1983, 3, 313–320. [Google Scholar]
- Tortora, G.J.; Funke, B.R.; Case, C.L. Introducción a la Microbiología; Médica Panamericana: Buenos Aires, Argentina, 2007; p. 912. [Google Scholar]
- Stow, M.; Starkey, B.J.; Hancock, I.C.; Baddiley, J. Inhibition by Chloramphenicol of Glucose Transfer in Teichoic Acid Biosynthesis. Nat. New Biol. 1971, 229, 56–57. [Google Scholar] [CrossRef] [PubMed]
- Inukai, M.; Isono, F.; Takatsuki, A. Selective inhibition of the bacterial translocase reaction in peptidoglycan synthesis by mureidomycins. Antimicrob. Agents Chemother. 1993, 37, 980–983. [Google Scholar] [CrossRef] [PubMed]
- Delcour, A.H. Outer membrane permeability and antibiotic resistance. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2009, 1794, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Z.; Plésiat, P.; Nikaido, H. The Challenge of Efflux-Mediated Antibiotic Resistance in Gram-Negative Bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef]
- Zhou, Q.; Wang, M.; Zhong, X.; Liu, P.; Xie, X.; Wangxiao, J.; Sun, Y. Dissemination of resistance genes in duck/fish polyculture ponds in Guangdong Province: Correlations between Cu and Zn and antibiotic resistance genes. Environ. Sci. Pollut. Res. 2019, 26, 8182–8193. [Google Scholar] [CrossRef]
- Voolaid, V.; Jõers, A.; Kisand, V.; Tenson, T. Co-occurrence of resistance to different antibiotics among aquatic bacteria. BMC Microbiol. 2012, 12, 225. [Google Scholar] [CrossRef]
- Remenár, M.; Karelová, E.; Harichová, J.; Zámocký, M.; Krčová, K.; Ferianc, P. Actinobacteria occurrence and their metabolic characteristics in the nickel-contaminated soil sample. Biologia 2014, 69, 1453–1463. [Google Scholar] [CrossRef]
- Jelić, D.; Antolović, R. From Erythromycin to Azithromycin and New Potential Ribosome-Binding Antimicrobials. Antibiotics 2016, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Ying, L.; Tang, D. Recent advances in the medicinal chemistry of novel erythromycin-derivatized antibiotics. Curr. Top. Med. Chem. 2010, 10, 1441–1469. [Google Scholar] [CrossRef] [PubMed]
- Cameron, J.C.; Pakrasi, H.B. Glutathione Facilitates Antibiotic Resistance and Photosystem I Stability during Exposure to Gentamicin in Cyanobacteria. Appl. Environ. Microbiol. 2011, 77, 3547–3550. [Google Scholar] [CrossRef] [PubMed]
- Dias, E.; Oliveira, M.; Jones-Dias, D.; Vasconcelos, V.; Ferreira, E.; Manageiro, V.; Caniça, M. Assessing the antibiotic susceptibility of freshwater Cyanobacteria spp. Front. Microbiol. 2015, 6, 799. [Google Scholar] [CrossRef]
- Van Le, V.; Tran, Q.-G.; Ko, S.-R.; Lee, S.-A.; Oh, H.-M.; Kim, H.-S.; Ahn, C.-Y. How do freshwater microalgae and cyanobacteria respond to antibiotics? Crit. Rev. Biotechnol. 2022, 43, 191–211. [Google Scholar] [CrossRef]
- Wu, Y.; Wan, L.; Zhang, W.; Ding, H.; Yang, W. Resistance of cyanobacteria Microcystis aeruginosa to erythromycin with multiple exposure. Chemosphere 2020, 249, 126147. [Google Scholar] [CrossRef]
- Shestakov, S.V.; Mitronova, T.N. Mutants of the blue-green alga Anacystis nidulans resistant to erythromycin and streptomycin. Nauchnye Dokl. Vyss. Shkoly. Biol. Nauk. 1971, 2, 98–102. [Google Scholar]
- Dias, E.; Oliveira, M.; Manageiro, V.; Vasconcelos, V.; Caniça, M. Deciphering the role of cyanobacteria in water resistome: Hypothesis justifying the antibiotic resistance (phenotype and genotype) in Planktothrix genus. Sci. Total Environ. 2019, 652, 447–454. [Google Scholar] [CrossRef]
- Kotra, L.P.; Haddad, J.; Mobashery, S. Aminoglycosides: Perspectives on Mechanisms of Action and Resistance and Strategies to Counter Resistance. Antimicrob. Agents Chemother. 2000, 44, 3249–3256. [Google Scholar] [CrossRef]
- Mingeot-Leclercq, M.-P.; Glupczynski, Y.; Tulkens, P.M. Aminoglycosides: Activity and Resistance. Antimicrob. Agents Chemother. 1999, 43, 727–737. [Google Scholar] [CrossRef]
- Brittain, D.C. Erythromycin. Med. Clin. N. Am. 1987, 71, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Le Page, G.; Gunnarsson, L.; Trznadel, M.; Wedgwood, K.C.; Baudrot, V.; Snape, J.; Tyler, C.R. Variability in cyanobacteria sensitivity to antibiotics and implications for environmental risk assessment. Sci. Total Environ. 2019, 695, 133804. [Google Scholar] [CrossRef] [PubMed]
- Frère, J.-M. Mechanism of action of β-lactam antibiotics at the molecular level. Biochem. Pharmacol. 1977, 26, 2203–2210. [Google Scholar] [CrossRef] [PubMed]
- Aurilio, C.; Sansone, P.; Barbarisi, M.; Pota, V.; Giaccari, L.G.; Coppolino, F.; Barbarisi, A.; Passavanti, M.B.; Pace, M.C. Mechanisms of Action of Carbapenem Resistance. Antibiotics 2022, 11, 421. [Google Scholar] [CrossRef] [PubMed]
- Baselga-Cervera, B.; Cordoba-Diaz, M.; García-Balboa, C.; Costas, E.; López-Rodas, V.; Cordoba-Diaz, D. Assessing the effect of high doses of ampicillin on five marine and freshwater phytoplankton species: A biodegradation perspective. J. Appl. Phycol. 2019, 31, 2999–3010. [Google Scholar] [CrossRef]
- Halling-Sørensen, B. Algal toxicity of antibacterial agents used in intensive farming. Chemosphere 2000, 40, 731–739. [Google Scholar] [CrossRef]
- Du, Y.; Wang, J.; Zhu, F.; Mai, D.; Xiang, Z.; Chen, J.; Guo, R. Comprehensive assessment of three typical antibiotics on cyanobacteria (Microcystis aeruginosa): The impact and recovery capability. Ecotoxicol. Environ. Saf. 2018, 160, 84–93. [Google Scholar] [CrossRef]
- Ke, Y.; Sun, W.; Jing, Z.; Zhao, Z.; Xie, S. Seasonal variations of microbial community and antibiotic resistome in a suburb drinking water distribution system in a northern Chinese city. J. Environ. Sci. 2023, 127, 714–725. [Google Scholar] [CrossRef]
- Zhang, Y.; Shen, G.; Hu, S.; He, Y.; Li, P.; Zhang, B. Deciphering of antibiotic resistance genes (ARGs) and potential abiotic indicators for the emergence of ARGs in an interconnected lake-river-reservoir system. J. Hazard. Mater. 2020, 410, 124552. [Google Scholar] [CrossRef]
- Du, C.; Yang, F.; Li, X.; Liao, H.; Li, Z.; Gao, J.; Zhang, L. Metagenomic analysis of microbial community structure and distribution of resistance genes in Daihai Lake, China. Environ. Pollut. 2022, 302, 119065. [Google Scholar] [CrossRef]
- Dias, M.F.; de Castro, G.M.; de Paiva, M.C.; Reis, M.d.P.; Facchin, S.; Carmo, A.O.D.; Alves, M.S.; Suhadolnik, M.L.; Motta, A.d.M.; Henriques, I.; et al. Exploring antibiotic resistance in environmental integron-cassettes through intI-attC amplicons deep sequencing. Braz. J. Microbiol. 2020, 52, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Shan, A.; Wang, W.; Kang, K.J.; Hou, D.; Luo, J.; Wang, G.; Pan, M.; Feng, Y.; He, Z.; Yang, X. The Removal of Antibiotics in Relation to a Microbial Community in an Integrated Constructed Wetland for Tail Water Decontamination. Wetlands 2020, 40, 993–1004. [Google Scholar] [CrossRef]
- Anderson, J. Evolution of antifungal-drug resistance: Mechanisms and pathogen fitness. Nat. Rev. Microbiol. 2005, 3, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Shang, M.; Xu, X.; Sun, J.; Zang, X. Analysis of morphological change mechanism of linear Arthrospira platensis based on transcriptome results. Gene 2022, 834, 146573. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.M.; da Silva, B.N.M.; Barbosa, G.; Barreiro, E.J. β-lactam antibiotics: An overview from a medicinal chemistry perspective. Eur. J. Med. Chem. 2020, 208, 112829. [Google Scholar] [CrossRef] [PubMed]
- Crea, F.; Cucinotta, D.; De Stefano, C.; Milea, D.; Sammartano, S.; Vianelli, G. Modeling solubility, acid–base properties and activity coefficients of amoxicillin, ampicillin and (+)6-aminopenicillanic acid, in NaCl(aq) at different ionic strengths and temperatures. Eur. J. Pharm. Sci. 2012, 47, 661–677. [Google Scholar] [CrossRef] [PubMed]
- Thiele-Bruhn, S. Pharmaceutical antibiotic compounds in soils–a review. J. Plant Nutr. Soil Sci. 2003, 166, 145–167. [Google Scholar] [CrossRef]
- Scaria, J.; Anupama, K.; Nidheesh, P. Tetracyclines in the environment: An overview on the occurrence, fate, toxicity, detection, removal methods, and sludge management. Sci. Total Environ. 2021, 771, 145291. [Google Scholar] [CrossRef]
- Oka, H.; Ikai, Y.; Kawamura, N.; Yamada, M.; Harada, K.; Ito, S.; Suzuki, M. Photodecomposition products of tetracycline in aqueous solution. J. Agric. Food Chem. 1989, 37, 226–231. [Google Scholar] [CrossRef]
- Addamo, M.; Augugliaro, V.; Di Paola, A.; García-López, E.; Loddo, V.; Marcì, G.; Palmisano, L. Removal of drugs in aqueous systems by photoassisted degradation. J. Appl. Electrochem. 2005, 35, 765–774. [Google Scholar] [CrossRef]
- Ahmad, F.; Zhu, D.; Sun, J. Correction to: Environmental fate of tetracycline antibiotics: Degradation pathway mechanisms, challenges, and perspectives. Environ. Sci. Eur. 2021, 33, 64. [Google Scholar] [CrossRef]
- Shlaes, D.M. An update on tetracyclines. Curr. Opin. Investig. Drugs 2006, 7, 167–171. [Google Scholar] [PubMed]
- Wang, S.; Ji, B.; Zhang, M.; Gu, J.; Ma, Y.; Liu, Y. Tetracycline-induced decoupling of symbiosis in microalgal-bacterial granular sludge. Environ. Res. 2021, 197, 111095. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Tang, Z.; Zhou, F.; Zhang, W.; Song, L. Toxicity studies of tetracycline on Microcystis aeruginosa and Selenastrum capricornutum. Environ. Toxicol. Pharmacol. 2013, 35, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Shang, A.H.; Ye, J.; Chen, D.H.; Lu, X.X.; Lu, H.D.; Liu, C.N.; Wang, L.M. Physiological effects of tetracycline antibiotic pollutants on non-target aquatic Microcystis aeruginosa. J. Environ. Sci. Health Part B-Pestic. Food Contam. Agric. Wastes 2015, 50, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, Q.; Zhang, J.; Guan, T.; Chen, Y.; Shi, W. Critical roles of cyanobacteria as reservoir and source for antibiotic resistance genes. Environ. Int. 2020, 144, 106034. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Song, H.L.; Yang, X.L.; Long, X.Z.; Liu, X.; Chen, T.Q. Behavior of tetracycline and sulfamethoxazole and their corresponding resistance genes in three-dimensional biofilm-electrode reactors with low current. J. Environ. Sci. Health Part A-Toxic/Hazard. Subst. Environ. Eng. 2017, 52, 333–340. [Google Scholar] [CrossRef]
- Ferech, M.; Coenen, S.; Dvorakova, K.; Hendrick, E.; Suetens, C.; Goossens, H.; Grp, E.P. European Surveillance of Antimicrobial Consumption (ESAC): Outpatient penicillin use in Europe. J. Antimicrob. Chemother. 2006, 58, 408–412. [Google Scholar] [CrossRef]
- Bruyndonckx, R.; Adriaenssens, N.; Hens, N.; Versporten, A.; Monnet, D.L.; Molenberghs, G.; Goossens, H.; Weist, K.; Coenen, S.; Grp, E.S.-N.S. Consumption of penicillins in the community, European Union/European Economic Area, 1997–2017. J. Antimicrob. Chemother. 2021, 76, 14–21. [Google Scholar] [CrossRef]
- Seppälä, H.; Klaukka, T.; Lehtonen, R.; Nenonen, E.; Huovinen, P. Outpatient Use of Erythromycin: Link to Increased Erythromycin Resistance in Group A Streptococci. Clin. Infect. Dis. 1995, 21, 1378–1385. [Google Scholar] [CrossRef]
- Zhang, T.; Li, B. Occurrence, Transformation, and Fate of Antibiotics in Municipal Wastewater Treatment Plants. Crit. Rev. Environ. Sci. Technol. 2011, 41, 951–998. [Google Scholar] [CrossRef]
- Ruan, Y.; Wu, R.; Lam, J.C.; Zhang, K.; Lam, P.K. Seasonal occurrence and fate of chiral pharmaceuticals in different sewage treatment systems in Hong Kong: Mass balance, enantiomeric profiling, and risk assessment. Water Res. 2019, 149, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Maiga, A.; Nguessend, N.A.; Dicko, O.A.; Fofana, M.; Maiga, I.I.; Agbo, K.S.M.; Dosso, M. The Hospital Wastewater Antibiotic Residues in Bamako (Mali). Le Mali Med. 2018, 33, 23–28. [Google Scholar] [PubMed]
- Santos, L.H.M.L.M.; Gros, M.; Rodriguez-Mozaz, S.; Delerue-Matos, C.; Pena, A.; Barceló, D.; Montenegro, M.C.B.S.M. Contribution of hospital effluents to the load of pharmaceuticals in urban wastewaters: Identification of ecologically relevant pharmaceuticals. Sci. Total Environ. 2013, 461–462, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Al-Maadheed, S.; Goktepe, I.; Latiff, A.B.A.; Shomar, B. Antibiotics in hospital effluent and domestic wastewater treatment plants in Doha, Qatar. J. Water Process. Eng. 2019, 28, 60–68. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, H.; Xiong, P.; Zhu, Q.; Liao, C.; Jiang, G. Occurrence, fate, and risk assessment of typical tetracycline antibiotics in the aquatic environment: A review. Sci. Total Environ. 2020, 753, 141975. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Guo, C.; Luo, Y.; Lv, J.; Zhang, Y.; Lin, H.; Wang, L.; Xu, J. Occurrence and distribution of antibiotics, antibiotic resistance genes in the urban rivers in Beijing, China. Environ. Pollut. 2016, 213, 833–840. [Google Scholar] [CrossRef]
- Zhang, Q.-Q.; Ying, G.-G.; Pan, C.-G.; Liu, Y.-S.; Zhao, J.-L. Comprehensive Evaluation of Antibiotics Emission and Fate in the River Basins of China: Source Analysis, Multimedia Modeling, and Linkage to Bacterial Resistance. Environ. Sci. Technol. 2015, 49, 6772–6782. [Google Scholar] [CrossRef]
- Borghi, A.A.; Palma, M.S.A. Tetracycline: Production, waste treatment and environmental impact assessment. Braz. J. Pharm. Sci. 2014, 50, 25–40. [Google Scholar] [CrossRef]
- Azanu, D.; Styrishave, B.; Darko, G.; Weisser, J.J.; Abaidoo, R.C. Occurrence and risk assessment of antibiotics in water and lettuce in Ghana. Sci. Total Environ. 2018, 622, 293–305. [Google Scholar] [CrossRef]
- Boger, B.; Surek, M.; Vilhena, R.d.O.; Fachi, M.M.; Junkert, A.M.; Santos, J.M.M.F.; Domingos, E.L.; Cobre, A.d.F.; Momade, D.R.; Pontarolo, R. Occurrence of antibiotics and antibiotic resistant bacteria in subtropical urban rivers in Brazil. J. Hazard. Mater. 2021, 402, 123448. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.C.; Keller, V.; Dumont, E.; Sumpter, J.P. Assessing the concentrations and risks of toxicity from the antibiotics ciprofloxacin, sulfamethoxazole, trimethoprim and erythromycin in European rivers. Sci. Total Environ. 2015, 511, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Lew, S.; Glińska-Lewczuk, K. Environmental controls on the abundance of methanotrophs and methanogens in peat bog lakes. Sci. Total Environ. 2018, 645, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Semedo, M.; Song, B. From Genes to Nitrogen Removal: Determining the Impacts of Poultry Industry Wastewater on Tidal Creek Denitrification. Environ. Sci. Technol. 2019, 54, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; LaVergne, J.M.; Carpenter, C.M.G.; Desai, R.; Zhang, X.; Gray, K.; Helbling, D.E.; Wells, G.F. Exploring co-occurrence patterns between organic micropollutants and bacterial community structure in a mixed-use watershed. Environ. Sci. Process. Impacts 2019, 21, 867–880. [Google Scholar] [CrossRef]
- Hagberg, A.; Gupta, S.; Rzhepishevska, O.; Fick, J.; Burmølle, M.; Ramstedt, M. Do environmental pharmaceuticals affect the composition of bacterial communities in a freshwater stream? A case study of the Knivsta river in the south of Sweden. Sci. Total Environ. 2021, 763, 142991. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Yan, D.; Wang, Q.; Huang, B.; Ren, Z.; Wang, X.; Wang, X.; Li, Y.; Ouyang, C.; Migheli, Q.; et al. Changes in the abundance and community composition of different nitrogen cycling groups in response to fumigation with 1,3-dichloropropene. Sci. Total Environ. 2018, 650, 44–55. [Google Scholar] [CrossRef]
- Guo, X.-P.; Yang, Y.; Niu, Z.-S.; Lu, D.-P.; Zhu, C.-H.; Feng, J.-N.; Wu, J.-Y.; Chen, Y.-R.; Tou, F.-Y.; Liu, M.; et al. Characteristics of microbial community indicate anthropogenic impact on the sediments along the Yangtze Estuary and its coastal area, China. Sci. Total Environ. 2019, 648, 306–314. [Google Scholar] [CrossRef]
- Madrova, P.; Vetrovsky, T.; Omelka, M.; Grunt, M.; Smutna, Y.; Rapoport, D.; Vach, M.; Baldrian, P.; Kopecky, J.; Sagova-Mareckova, M. A Short-Term Response of Soil Microbial Communities to Cadmium and Organic Substrate Amendment in Long-Term Contaminated Soil by Toxic Elements. Front. Microbiol. 2018, 9, 2807. [Google Scholar] [CrossRef]
- Xin, X.; Huang, G.; Zhang, B. Review of aquatic toxicity of pharmaceuticals and personal care products to algae. J. Hazard. Mater. 2020, 410, 124619. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, G.; Zheng, Q.; Tang, J.; Chen, Y.; Xu, W.; Zou, Y.; Chen, X. Occurrence and risks of antibiotics in the Laizhou Bay, China: Impacts of river discharge. Ecotoxicol. Environ. Saf. 2012, 80, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Yin, Z.; Yang, Y.; Liang, Y.; Shi, H.; Xu, X. Investigation of the microbial community structure and diversity in the environment surrounding a veterinary antibiotic production factory. RSC Adv. 2022, 12, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhang, Z.; Feng, L.; Zhang, J.; Li, Y.; Lu, T.; Qian, H. Adverse effects of levofloxacin and oxytetracycline on aquatic microbial communities. Sci. Total Environ. 2020, 734, 139499. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Jiang, Y.; Liu, Y.; Zhang, J. Antibiotic-accelerated cyanobacterial growth and aquatic community succession towards the formation of cyanobacterial bloom in eutrophic lake water. Environ. Pollut. 2021, 290, 118057. [Google Scholar] [CrossRef] [PubMed]
- Goodman, J.J.; Matrishin, M.; Backus, E.J. The effect of anhydrochlortetracycline on the growth of actinomycetes. J. Bacteriol. 1955, 69, 70–72. [Google Scholar] [CrossRef]
- Lu, T.; Zhu, Y.; Ke, M.; Peijnenburg, W.; Zhang, M.; Wang, T.; Chen, J.; Qian, H. Evaluation of the taxonomic and functional variation of freshwater plankton communities induced by trace amounts of the antibiotic ciprofloxacin. Environ. Int. 2019, 126, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Grenni, P.; Patrolecco, L.; Rauseo, J.; Spataro, F.; Di Lenola, M.; Aimola, G.; Zacchini, M.; Pietrini, F.; Di Baccio, D.; Stanton, I.C.; et al. Sulfamethoxazole persistence in a river water ecosystem and its effects on the natural microbial community and Lemna minor plant. Microchem. J. 2019, 149, 103999. [Google Scholar] [CrossRef]
- Cunha, B.A. Antibiotic Resistance: A Historical Perspective. Semin. Respir. Crit. Care Med. 2000, 21, 3–8. [Google Scholar] [CrossRef]
- Lin, Z.; Yuan, T.; Zhou, L.; Cheng, S.; Qu, X.; Lu, P.; Feng, Q. Impact factors of the accumulation, migration and spread of antibiotic resistance in the environment. Environ. Geochem. Health 2021, 43, 1741–1758. [Google Scholar] [CrossRef]
- World Health Organization. Monitoring and Evaluation of the Global Action Plan on Antimicrobial Resistance: Framework and Recommended Indicators; 2019; p. 32, ISBN: 9789241515665. Available online: https://www.who.int/publications/i/item/monitoring-and-evaluation-of-the-global-action-plan-on-antimicrobial-resistance (accessed on 30 January 2023).
- Grenni, P. Antimicrobial Resistance in Rivers: A Review of the Genes Detected and New Challenges. Environ. Toxicol. Chem. 2022, 41, 687–714. [Google Scholar] [CrossRef]
- Pino-Otín, M.R.; Ferrando, N.; Ballestero, D.; Langa, E.; Roig, F.J.; Terrado, E.M. Impact of eight widely consumed antibiotics on the growth and physiological profile of natural soil microbial communities. Chemosphere 2022, 305, 135473. [Google Scholar] [CrossRef]
- Garland, J.L. Analysis and interpretation of community-level physiological profiles in microbial ecology. Fems Microbiol. Ecol. 1997, 24, 289–300. [Google Scholar] [CrossRef]
- Gu, M.Y.; Zhang, Z.D.; Wang, W.; Tang, Q.Y.; Song, S.Q.; Zhu, J.; Xie, Y.Q.; Zhang, L.J. The Effects of Radiation Pollution on the Population Diversities and Metabolic Characteristics of Soil Microorganisms. Water Air Soil Pollut. 2014, 225, 2133. [Google Scholar] [CrossRef]
- Garland, J.L.; Mills, A.L. Classification and Characterization of Heterotrophic Microbial Communities on the Basis of Patterns of Community-Level Sole-Carbon-Source Utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.T.; Davis-Richardson, A.G.; Giongo, A.; Gano, K.A.; Crabb, D.B.; Mukherjee, N.; Casella, G.; Drew, J.C.; Ilonen, J.; Knip, M.; et al. Gut Microbiome Metagenomics Analysis Suggests a Functional Model for the Development of Autoimmunity for Type 1 Diabetes. PLoS ONE 2011, 6, e25792. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef]
- Peleg, M.; Corradini, M.G.; Normand, M.D. The logistic (Verhulst) model for sigmoid microbial growth curves revisited. Food Res. Int. 2007, 40, 808–818. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic Name | Abbr. | Family | CAS Number | MW (g/mol) | Water Solubility (mg/mL) | pKa | Log Ko/w | 1000 µg/mL | ||
|---|---|---|---|---|---|---|---|---|---|---|
| pKa1 | pKa2 | Cmax (1) | r (2) | |||||||
| Chloramphenicol | CHL | Amphenicols | 56-75-7 | 323.1 | 2.5 | 7.5 | −2.8 | 1.0 | 0.133 | 0.067 |
| Tetracycline | TC | Tetracyclines | 64-75-5 | 444.4 | 75.5 | 3.3 | 9.2 | −2.0 | 1.801 | 0.070 |
| Erythromycin | ERY | Macrolides | 114-07-8 | 733.9 | 2.0 | 8.9 | 8.9 | −3.0 | 0.645 | 0.043 |
| Streptomycin sulphate | STM | Aminoglycosides | 3810-74-0 | 581.6 | 75.0 | 10.9 | 12.0 | −5.2 | 0.602 | 0.037 |
| Gentamycin sulphate | GTM | 1405-41-0 | 447.6 | 50.0 | 12.6 | 10.1 | −2.4 | 0.420 | 0.035 | |
| Amoxicillin | AMO | Beta-lactams | 26787-78-0 | 356.4 | 1.0 | 2.9 | 11.7 | 0.9 | 0.645 | 0.043 |
| Ampicillin | AMP | 69-53-4 | 349.4 | 10.0 | 2.6 | 7.2 | 1.4 | 0.640 | 0.037 | |
| Penicillin G sodium salt | PEN | 69-57-8 | 356.4 | 75.0 | 3.5 | −2.8 | 1.2 | 0.650 | 0.042 | |
| Control | 1.081 | 0.043 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pino-Otín, M.R.; Lorca, G.; Langa, E.; Roig, F.; Terrado, E.M.; Ballestero, D. Assessing the Ecotoxicity of Eight Widely Used Antibiotics on River Microbial Communities. Int. J. Mol. Sci. 2023, 24, 16960. https://doi.org/10.3390/ijms242316960
Pino-Otín MR, Lorca G, Langa E, Roig F, Terrado EM, Ballestero D. Assessing the Ecotoxicity of Eight Widely Used Antibiotics on River Microbial Communities. International Journal of Molecular Sciences. 2023; 24(23):16960. https://doi.org/10.3390/ijms242316960
Chicago/Turabian StylePino-Otín, María Rosa, Guillermo Lorca, Elisa Langa, Francisco Roig, Eva M. Terrado, and Diego Ballestero. 2023. "Assessing the Ecotoxicity of Eight Widely Used Antibiotics on River Microbial Communities" International Journal of Molecular Sciences 24, no. 23: 16960. https://doi.org/10.3390/ijms242316960
APA StylePino-Otín, M. R., Lorca, G., Langa, E., Roig, F., Terrado, E. M., & Ballestero, D. (2023). Assessing the Ecotoxicity of Eight Widely Used Antibiotics on River Microbial Communities. International Journal of Molecular Sciences, 24(23), 16960. https://doi.org/10.3390/ijms242316960