

CXCR2-Blocking Has Context-Sensitive Effects on Rat Glioblastoma Cell Line Outgrowth (S635) in an Organotypic Rat Brain Slice Culture Depending on Microglia-Depletion (PLX5622) and Dexamethasone Treatment
 , , , ,
, , , ,  and
and 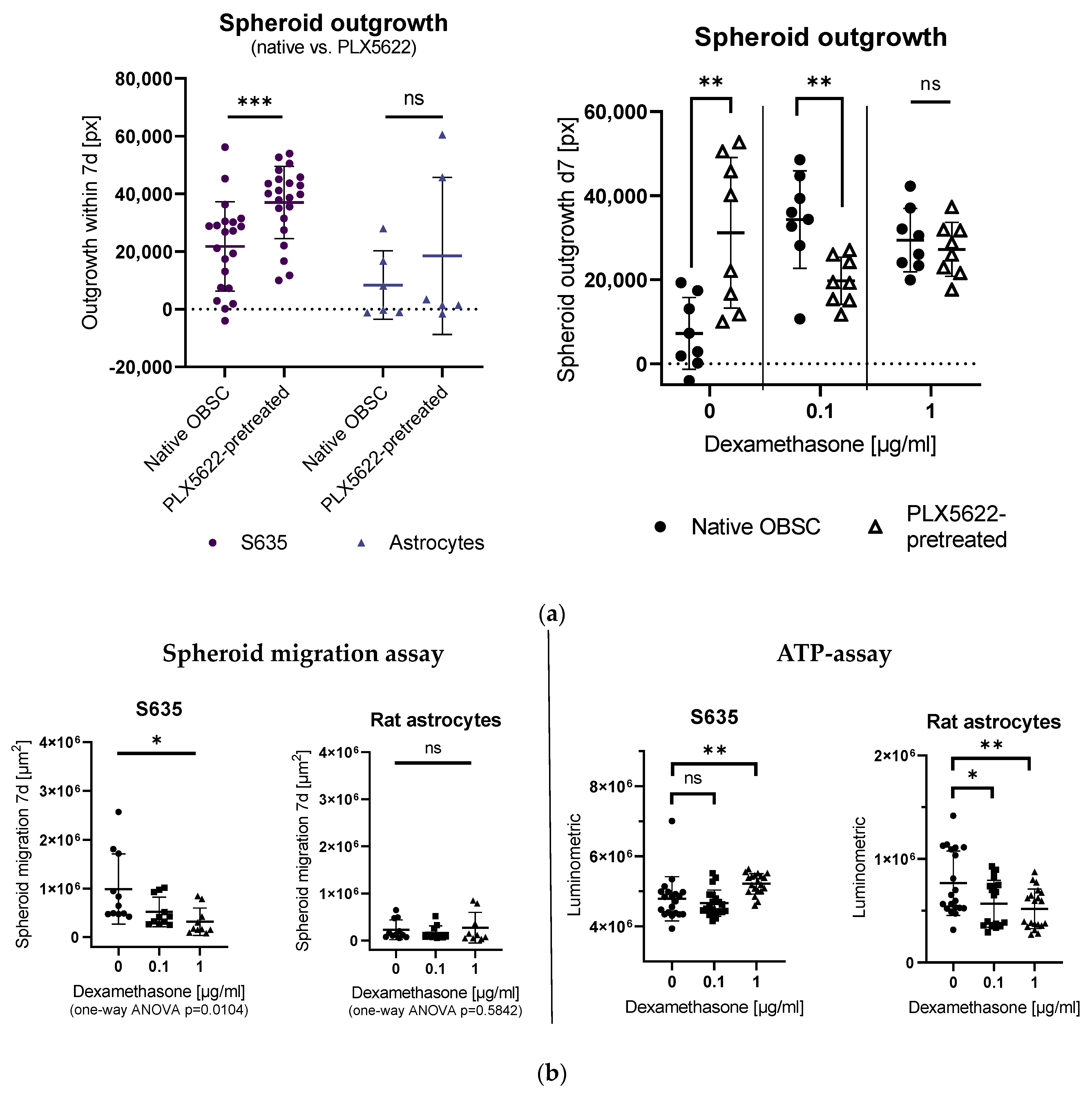{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
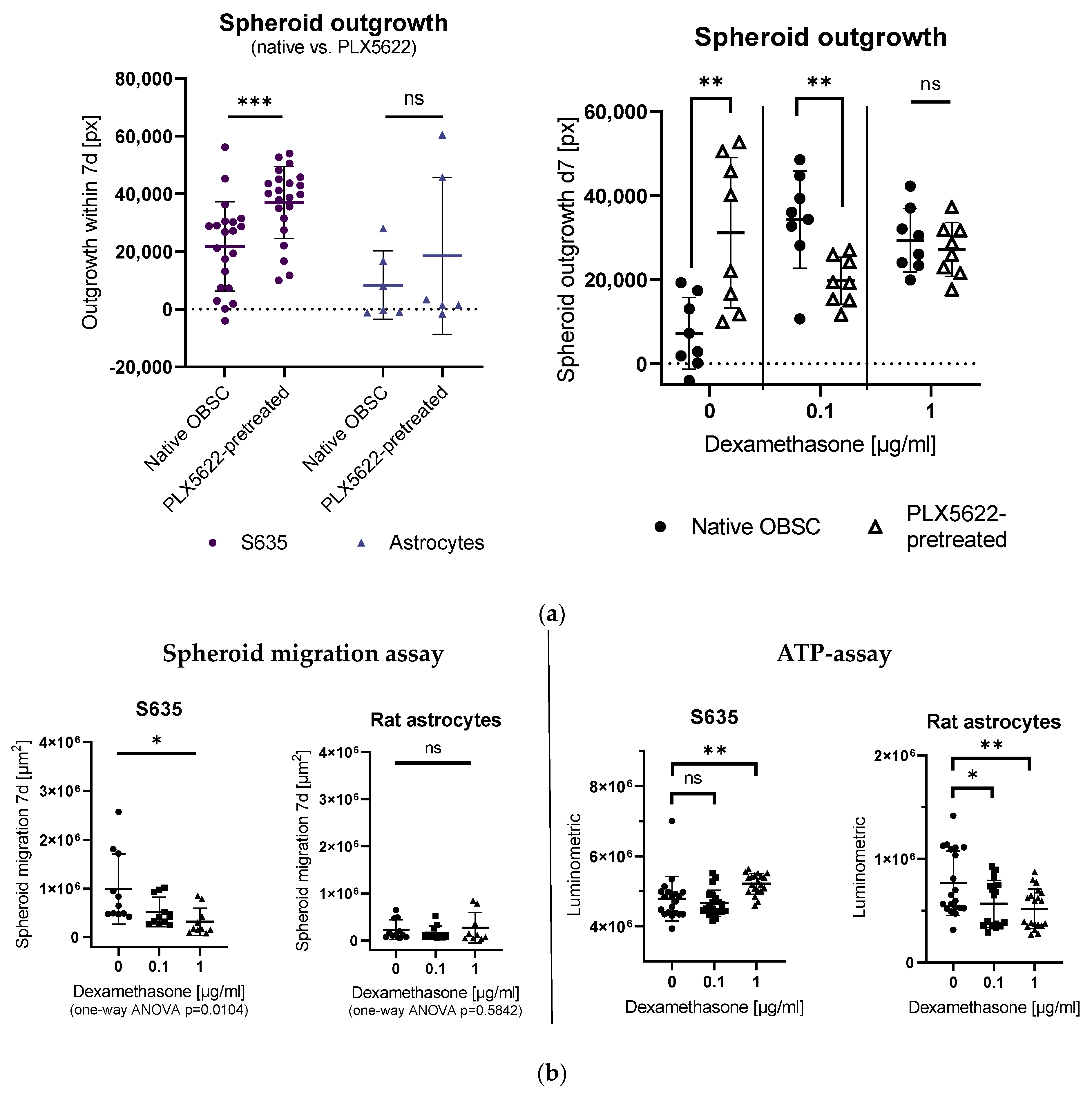
2.1. Effects of PLX5622-Pretreatment and Dexamethasone Treatment on S635 GBM Rat Cell Line Outgrowth
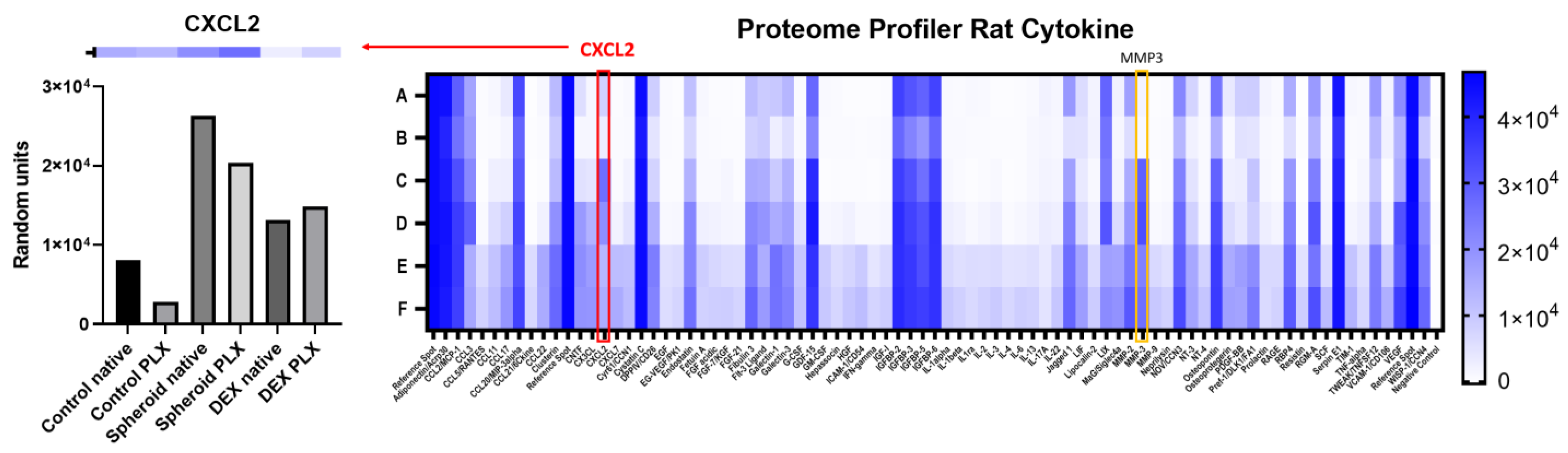
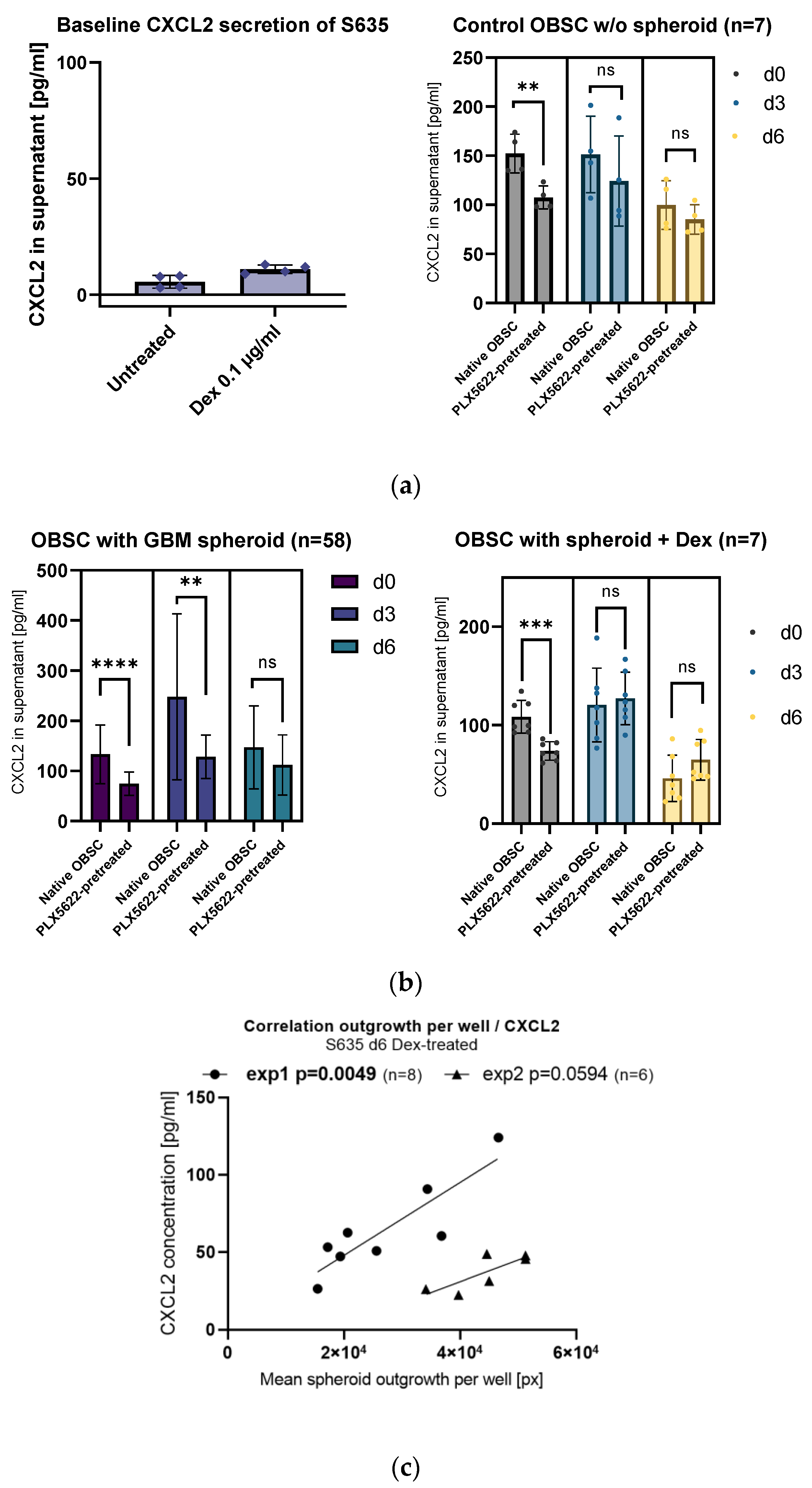
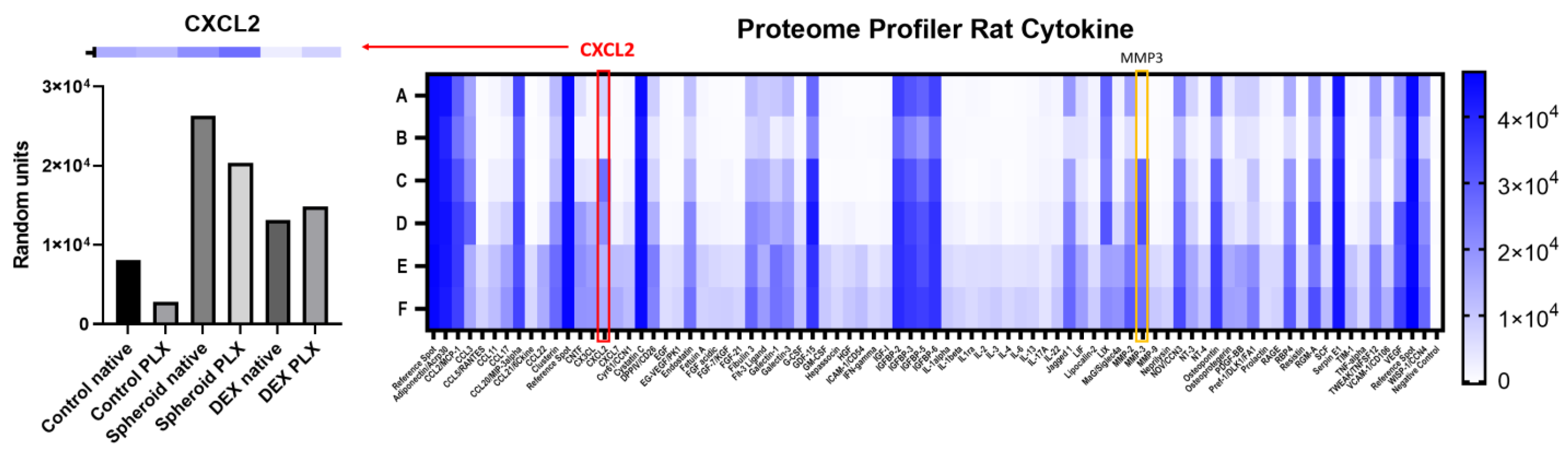
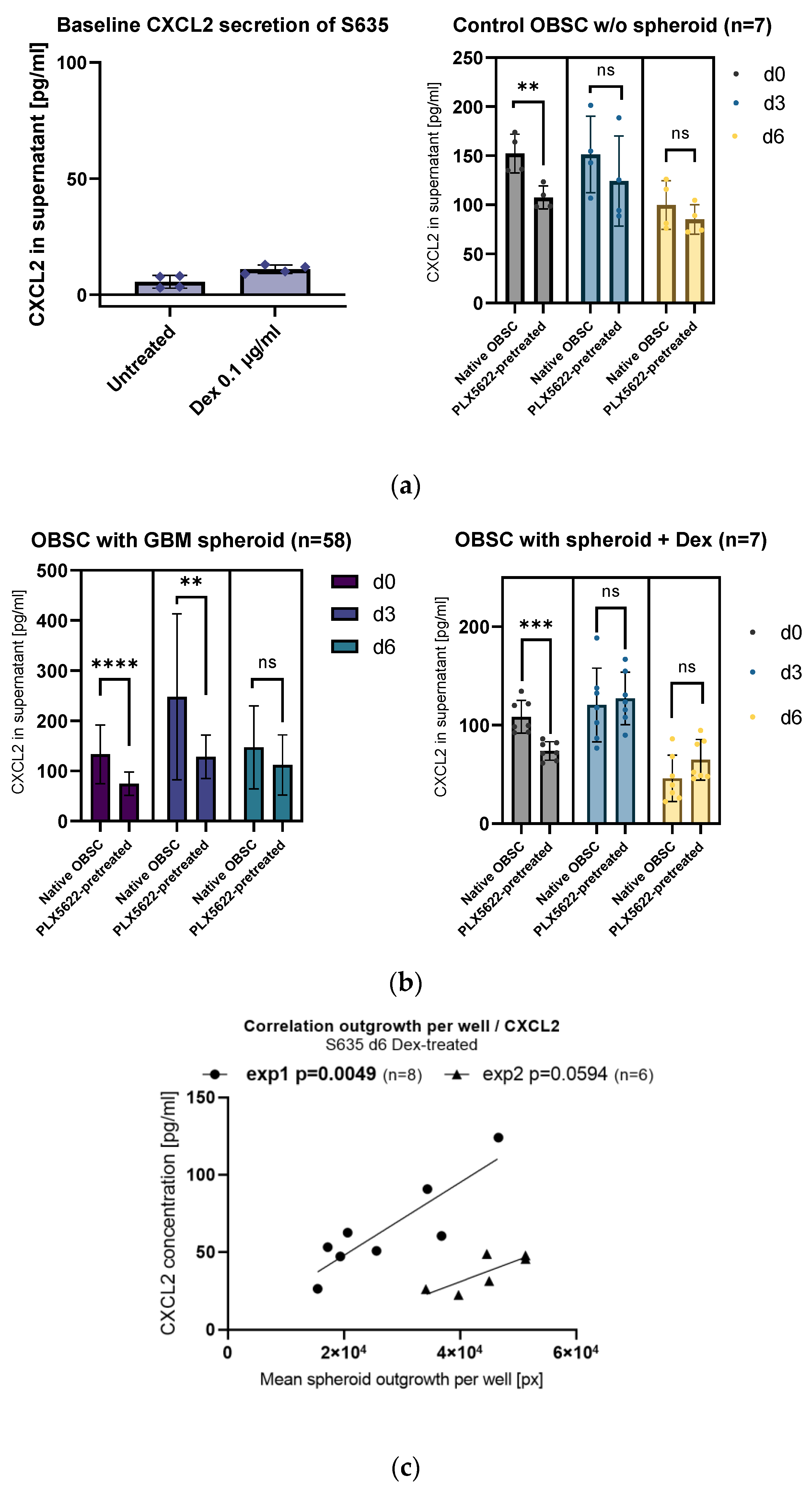
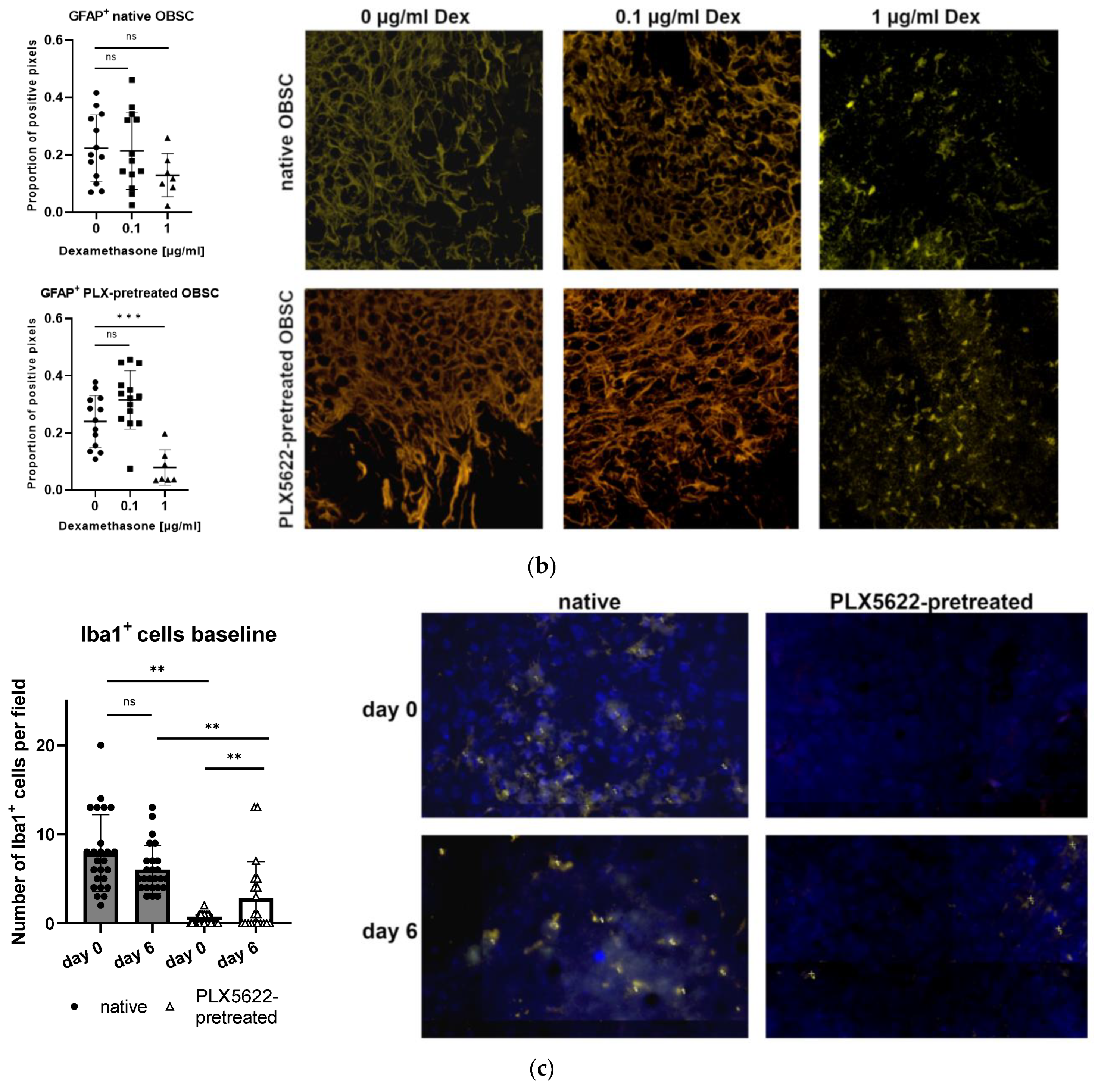
2.2. Influence of PLX5622-Pretreatment and Dexamethasone on CXCL2 Secretion
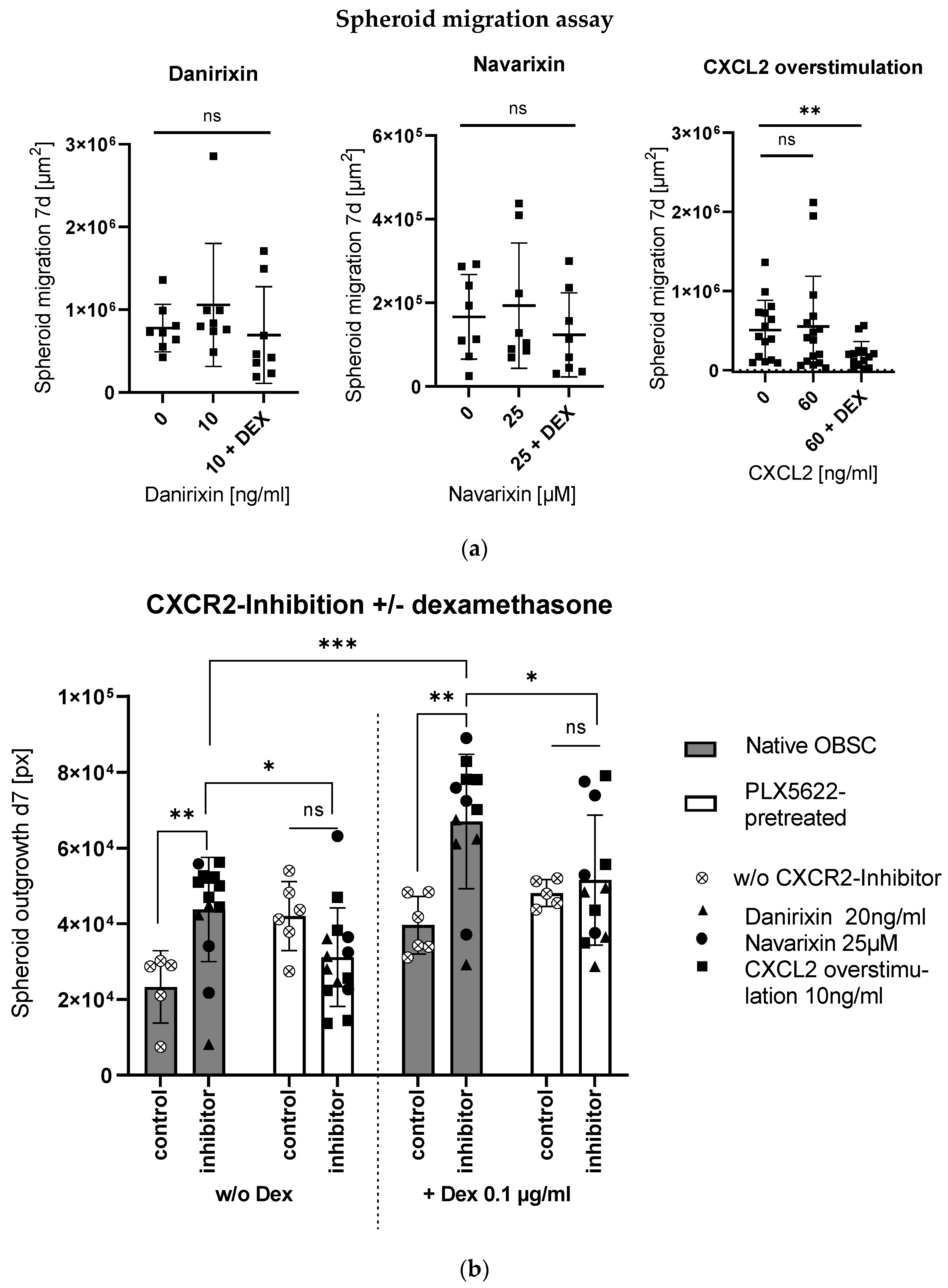
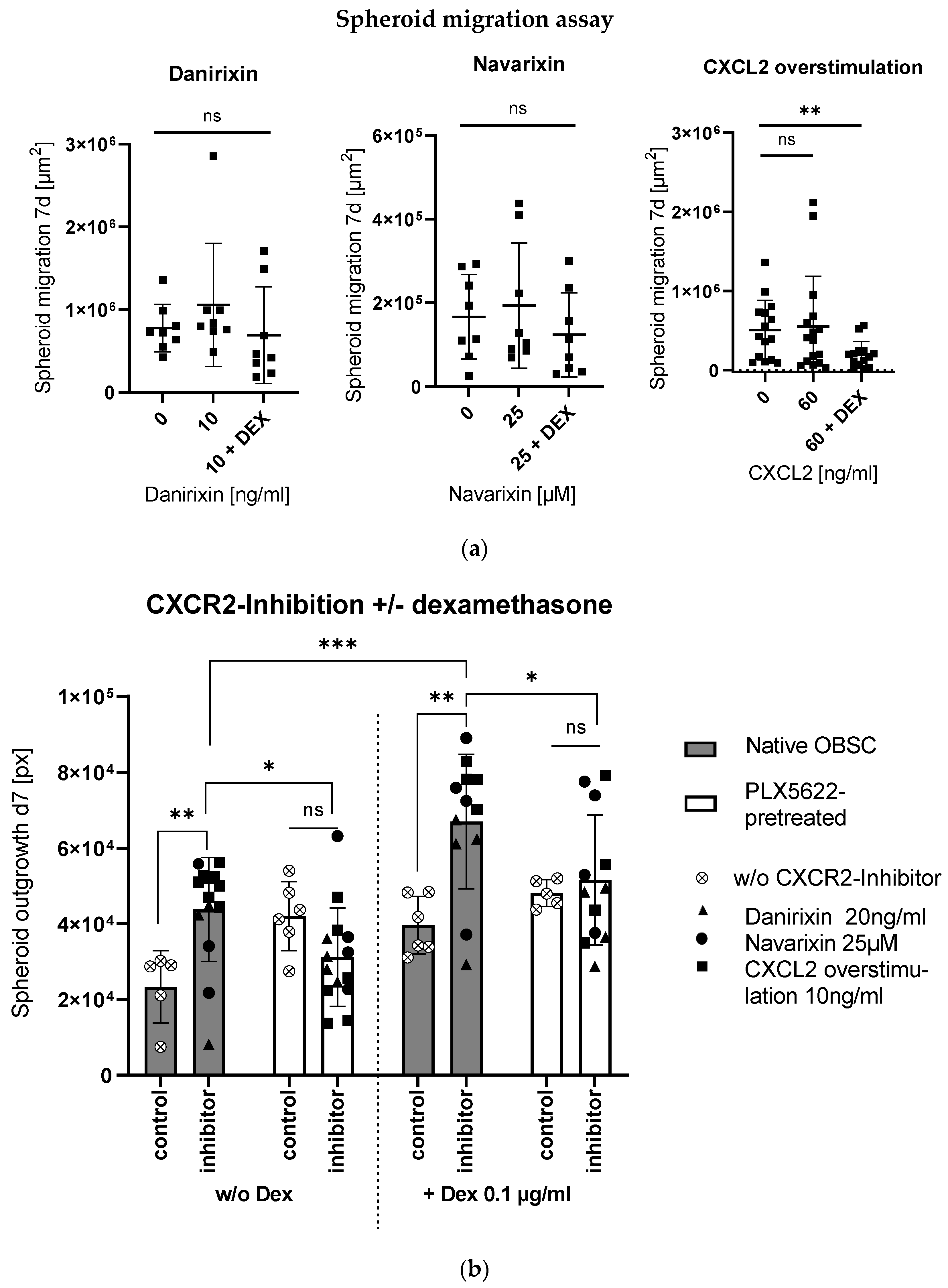
2.3. Context-Sensitive Impact of CXCL2-Receptor (CXCR2)-Blocking on Spheroid Outgrowth
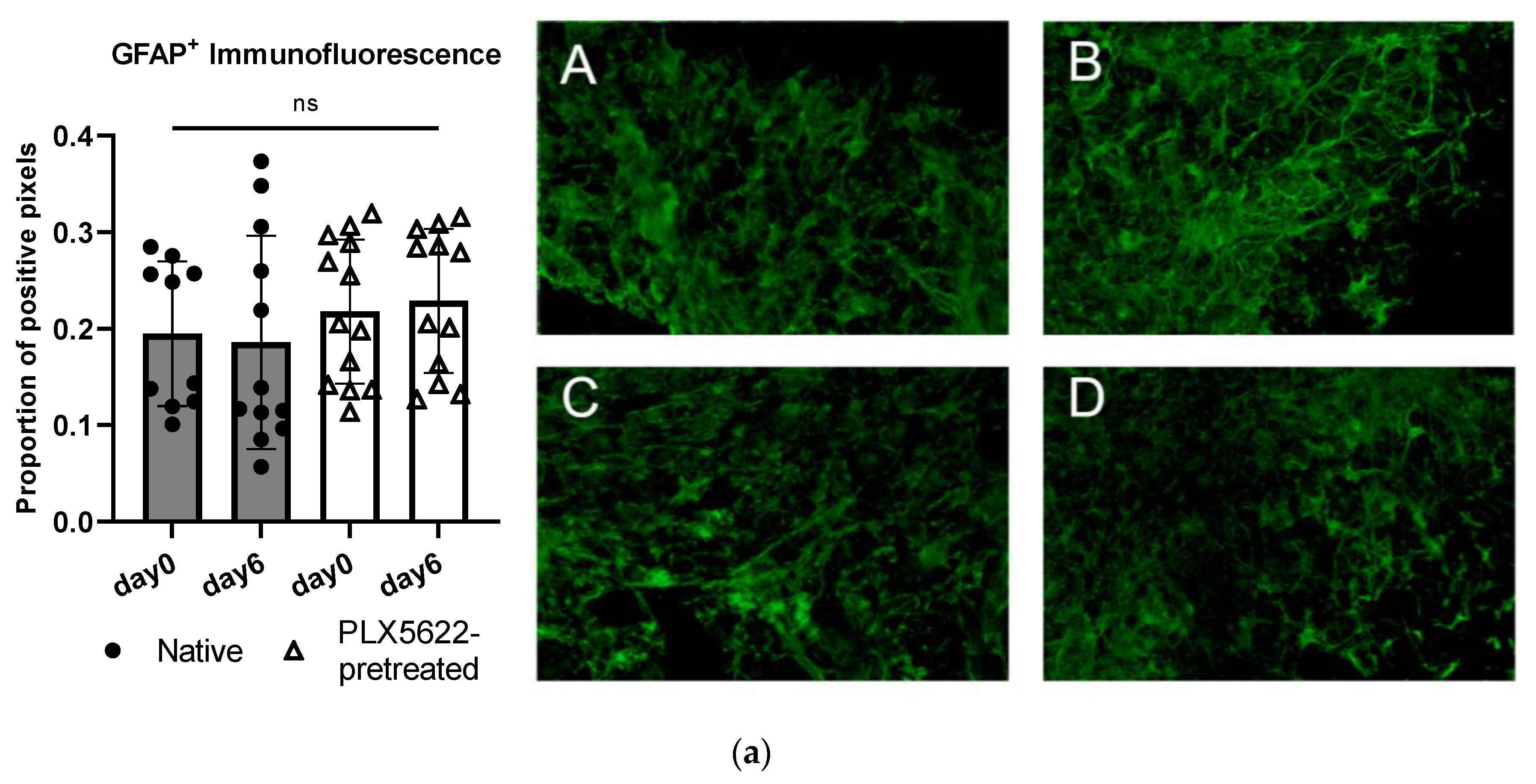
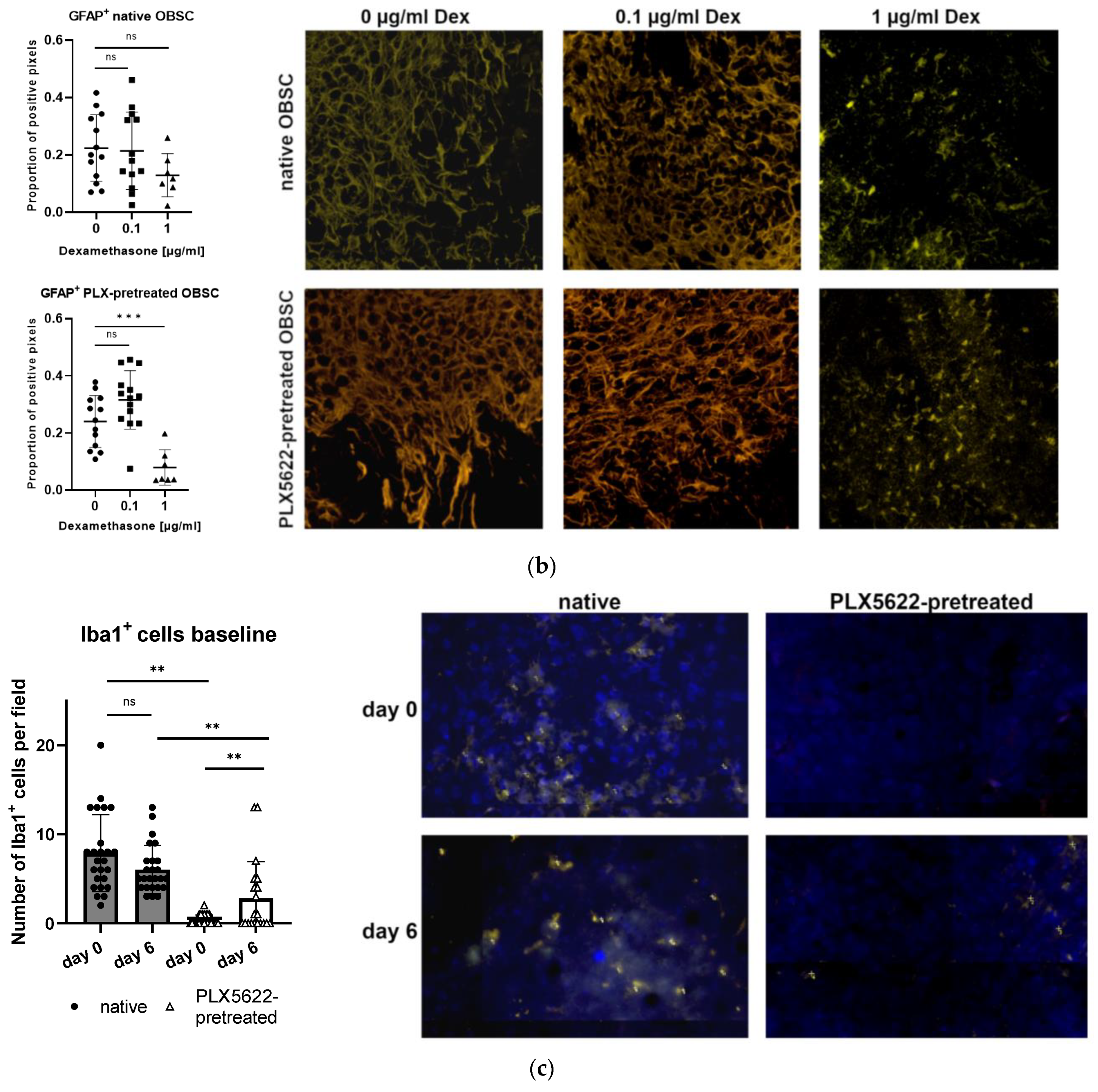
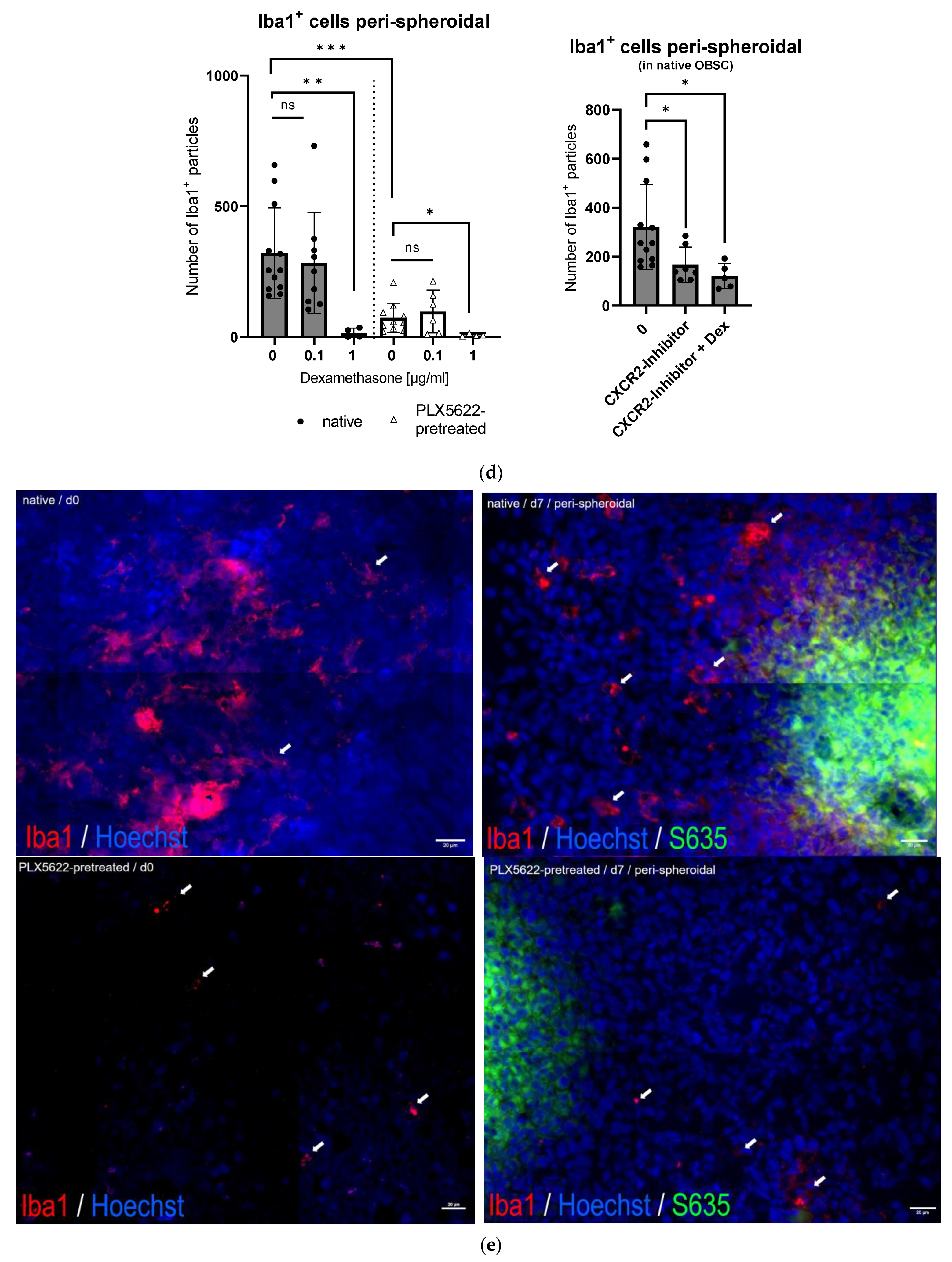
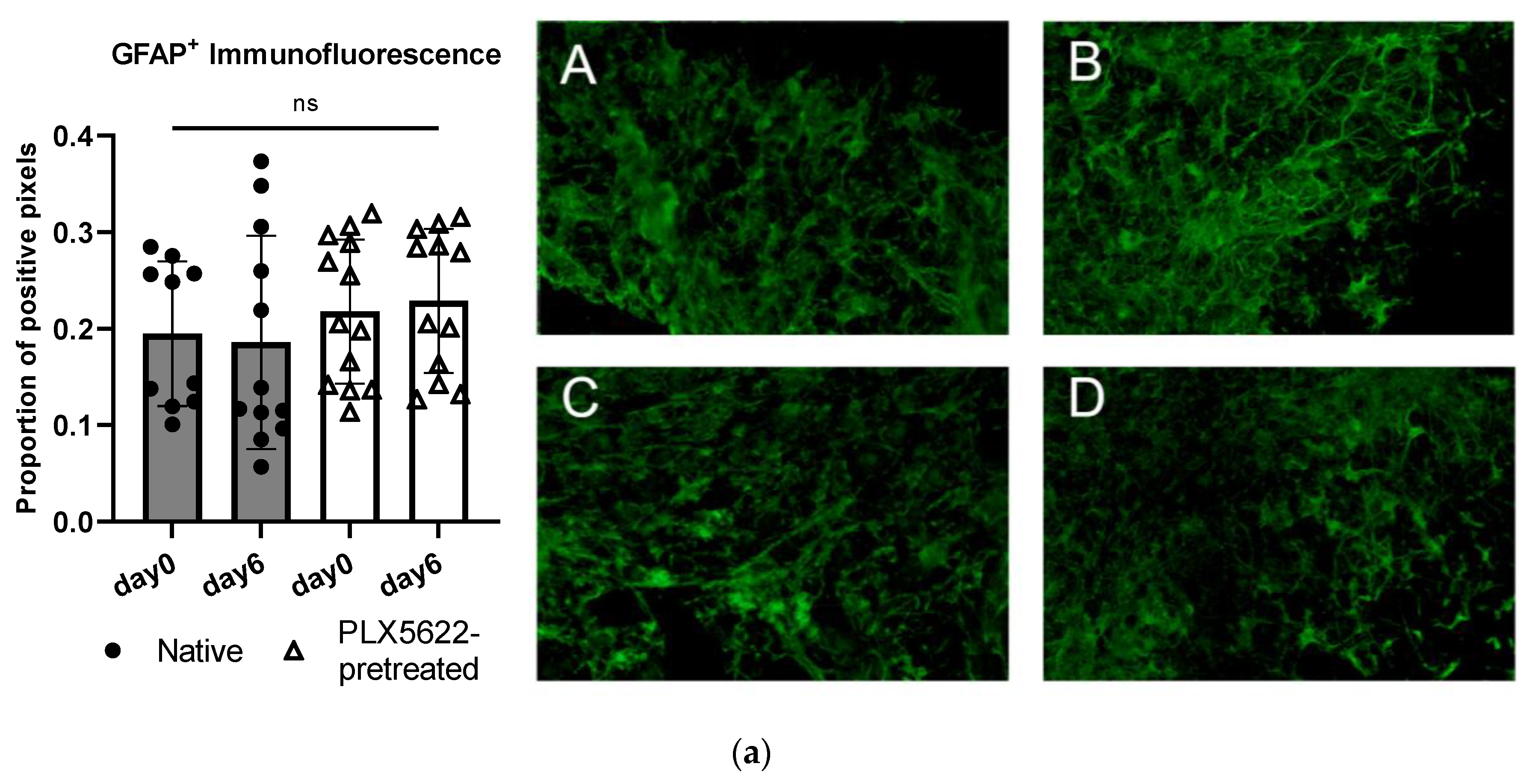
2.4. Cellular Correlate (Microglia [Iba1+] and Astrocytes [GFAP+]) within the Rat OBSC
3. Discussion
- External stimuli that can induce general microglia- and tumor microenvironment-modulating effects;
- Sustained unspecific effects of the CSF1-R inhibitor PLX5622 on various cell types of the brain slice co-culture, which might generally minimize the validity of the conclusions from the model;
- Our evaluations of the cytokine signaling only allow for analyzing the general humoral fingerprint/general CXCL2 secretion—the exact secreting cells remain elusive;
- CXCL1-3 and CXCL5-8 contain an ELR motif showing high binding affinity for CXCR2; CXCL3 or the other above-mentioned possible ligands were not included in the proteome profiler, nor did we check their secretion in our model;
- Our exploratory study was conducted only on a single cell line—control for cell line dependencies is not given and, therefore, generalizability is very limited.
4. Materials and Methods
4.1. Cells
4.2. Fluorescent Cell Labeling and Spheroid Formation
4.3. Spheroid Migration Assay
4.4. ATP-Assay
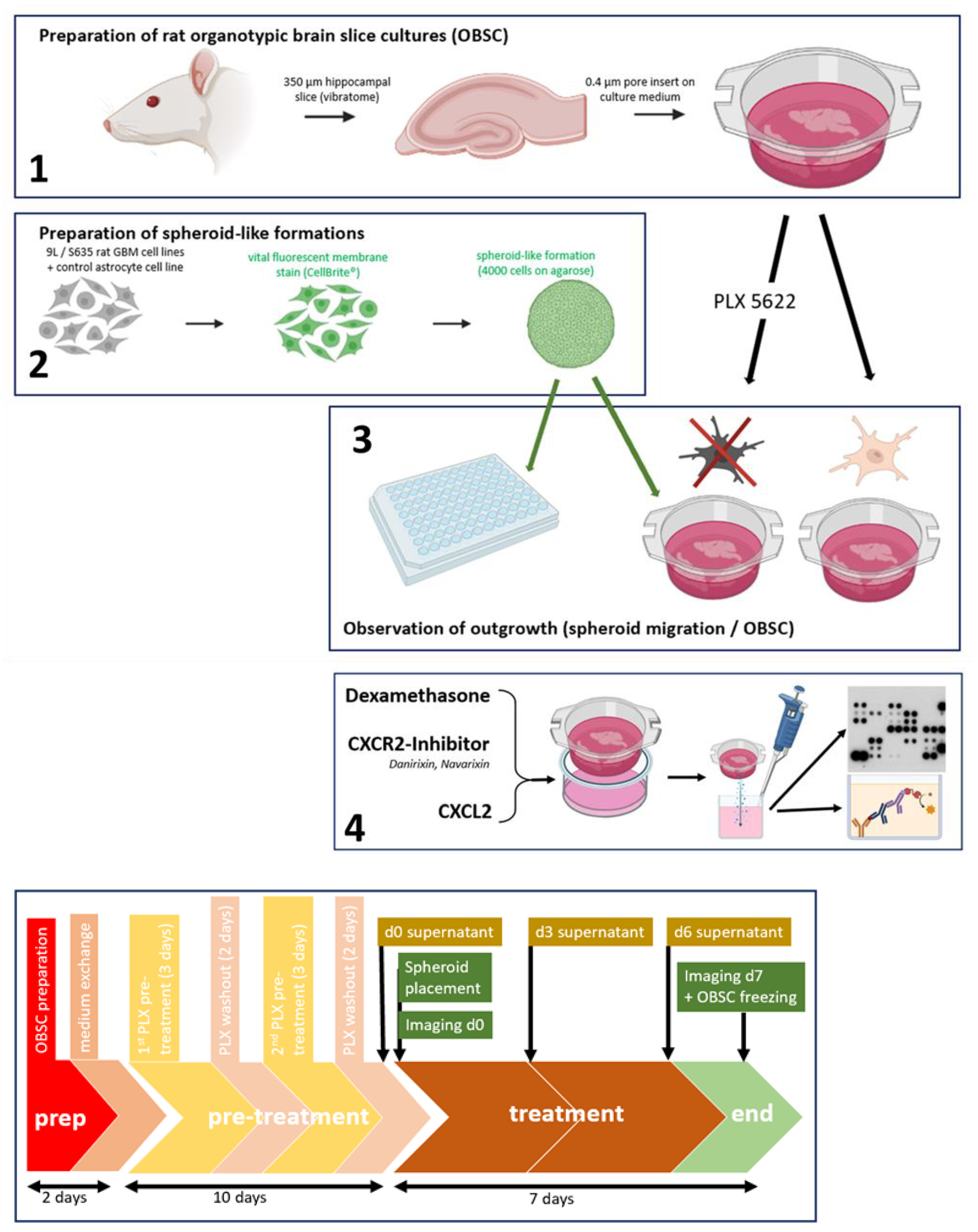
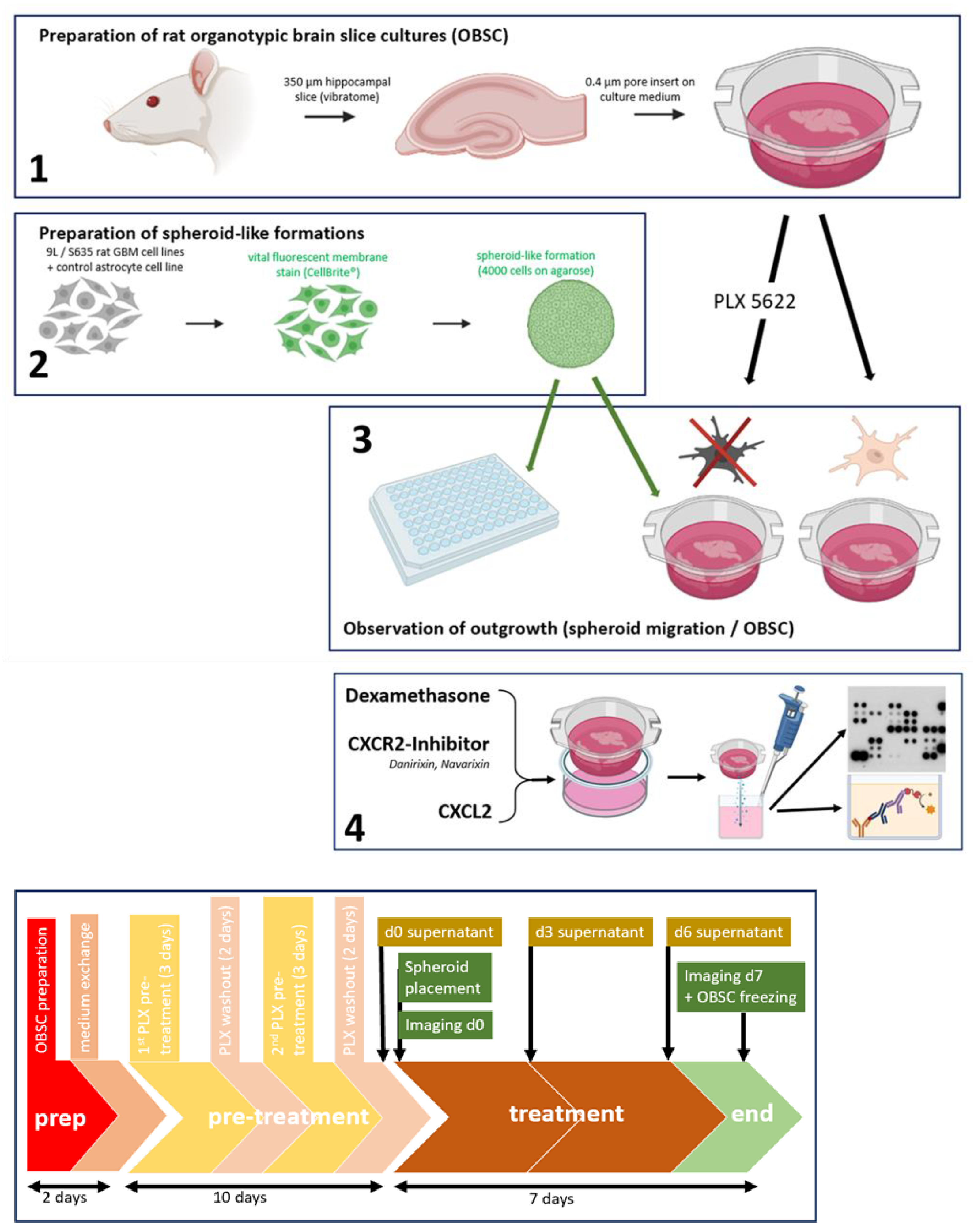
4.5. Ex Vivo Rat Brain Slice Assay—Organotypic Rat Brain Slice Culture (OBSC)
OBSC Preparation
4.6. PLX5622-Pretreatment of OBSC/Dexamethasone Treatment
4.7. Cytokine Screening Proteome Profiler
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. CXCL2/CXCR-Inhibitors
4.10. Immunofluorescence Staining (GFAP, Iba1)
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Habib, A.; Pease, M.; Kodavali, C.V.; Amankulor, N.; Zinn, P.O. A contemporary update on glioblastoma: Molecular biology, current management, and a vision towards bio-adaptable personalized care. J. Neurooncol. 2021, 151, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Kostaras, X.; Cusano, F.; Kline, G.A.; Roa, W.; Easaw, J. Use of dexamethasone in patients with high-grade glioma: A clinical practice guideline. Curr. Oncol. 2014, 21, e493–e503. [Google Scholar] [CrossRef] [PubMed]
- Kaup, B.; Schindler, I.; Knüpfer, H.; Schlenzka, A.; Preiss, R.; Knüpfer, M.M. Time-dependent inhibition of glioblastoma cell proliferation by dexamethasone. J. Neurooncol. 2001, 51, 105–110. [Google Scholar] [CrossRef]
- Piette, C.; Deprez, M.; Roger, T.; Noël, A.; Foidart, J.-M.; Munaut, C. The dexamethasone-induced inhibition of proliferation, migration, and invasion in glioma cell lines is antagonized by macrophage migration inhibitory factor (MIF) and can be enhanced by specific MIF inhibitors. J. Biol. Chem. 2009, 284, 32483–32492. [Google Scholar] [CrossRef]
- Villeneuve, J.; Galarneau, H.; Beaudet, M.-J.; Tremblay, P.; Chernomoretz, A.; Vallières, L. Reduced glioma growth following dexamethasone or anti-angiopoietin 2 treatment. Brain Pathol. 2008, 18, 401–414. [Google Scholar] [CrossRef]
- Petrelli, F.; de Stefani, A.; Ghidini, A.; Bruschieri, L.; Riboldi, V.; Dottorini, L.; Iaculli, A.; Zaniboni, A.; Trevisan, F. Steroids use and survival in patients with glioblastoma multiforme: A pooled analysis. J. Neurol. 2021, 268, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Shen, Y.; Huang, T.; Sun, Y.; Alolga, R.N.; Zhang, G.; Ge, Y. The Prognostic Effect of Dexamethasone on Patients with Glioblastoma: A Systematic Review and Meta-Analysis. Front. Pharmacol. 2021, 12, 727707. [Google Scholar] [CrossRef]
- Wong, E.T.; Lok, E.; Gautam, S.; Swanson, K.D. Dexamethasone exerts profound immunologic interference on treatment efficacy for recurrent glioblastoma. Br. J. Cancer 2015, 113, 232–241. [Google Scholar] [CrossRef]
- Shields, L.B.E.; Shelton, B.J.; Shearer, A.J.; Chen, L.; Sun, D.A.; Parsons, S.; Bourne, T.D.; LaRocca, R.; Spalding, A.C. Dexamethasone administration during definitive radiation and temozolomide renders a poor prognosis in a retrospective analysis of newly diagnosed glioblastoma patients. Radiat. Oncol. 2015, 10, 222. [Google Scholar] [CrossRef]
- Pitter, K.L.; Tamagno, I.; Alikhanyan, K.; Hosni-Ahmed, A.; Pattwell, S.S.; Donnola, S.; Dai, C.; Ozawa, T.; Chang, M.; Chan, T.A.; et al. Corticosteroids compromise survival in glioblastoma. Brain 2016, 139, 1458–1471. [Google Scholar] [CrossRef]
- Dubinski, D.; Won, S.-Y.; Gessler, F.; Quick-Weller, J.; Behmanesh, B.; Bernatz, S.; Forster, M.-T.; Franz, K.; Plate, K.-H.; Seifert, V.; et al. Dexamethasone-induced leukocytosis is associated with poor survival in newly diagnosed glioblastoma. J. Neurooncol. 2018, 137, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.S.; Zdioruk, M.; Nowicki, M.O.; Griffith, A.M.; Aguilar, E.; Aguilar, L.K.; Guzik, B.W.; Barone, F.; Tak, P.P.; Tabatabai, G.; et al. Systemic high-dose dexamethasone treatment may modulate the efficacy of intratumoral viral oncolytic immunotherapy in glioblastoma models. J. Immunother. Cancer 2022, 10, e003368. [Google Scholar] [CrossRef] [PubMed]
- Iorgulescu, J.B.; Gokhale, P.C.; Speranza, M.C.; Eschle, B.K.; Poitras, M.J.; Wilkens, M.K.; Soroko, K.M.; Chhoeu, C.; Knott, A.; Gao, Y.; et al. Concurrent Dexamethasone Limits the Clinical Benefit of Immune Checkpoint Blockade in Glioblastoma. Clin. Cancer Res. 2021, 27, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Giles, A.J.; Hutchinson, M.-K.N.D.; Sonnemann, H.M.; Jung, J.; Fecci, P.E.; Ratnam, N.M.; Zhang, W.; Song, H.; Bailey, R.; Davis, D.; et al. Dexamethasone-induced immunosuppression: Mechanisms and implications for immunotherapy. J. Immunother. Cancer 2018, 6, 51. [Google Scholar] [CrossRef] [PubMed]
- Keskin, D.B.; Anandappa, A.J.; Sun, J.; Tirosh, I.; Mathewson, N.D.; Li, S.; Oliveira, G.; Giobbie-Hurder, A.; Felt, K.; Gjini, E.; et al. Neoantigen vaccine generates intratumoral T cell responses in phase Ib glioblastoma trial. Nature 2019, 565, 234–239. [Google Scholar] [CrossRef]
- Wong, E.T.; Swanson, K.D. Dexamethasone-Friend or Foe for Patients with Glioblastoma? JAMA Neurol. 2019, 76, 247–248. [Google Scholar] [CrossRef]
- Cenciarini, M.; Valentino, M.; Belia, S.; Sforna, L.; Rosa, P.; Ronchetti, S.; D’Adamo, M.C.; Pessia, M. Dexamethasone in Glioblastoma Multiforme Therapy: Mechanisms and Controversies. Front. Mol. Neurosci. 2019, 12, 65. [Google Scholar] [CrossRef]
- Upadhyayula, P.S.; Higgins, D.M.; Argenziano, M.G.; Spinazzi, E.F.; Wu, C.-C.; Canoll, P.; Bruce, J.N. The Sledgehammer in Precision Medicine: Dexamethasone and Immunotherapeutic Treatment of Glioma. Cancer Investig. 2022, 40, 554–566. [Google Scholar] [CrossRef]
- Swildens, K.X.; Sillevis Smitt, P.A.E.; van den Bent, M.J.; French, P.J.; Geurts, M. The effect of dexamethasone on the microenvironment and efficacy of checkpoint inhibitors in glioblastoma: A systematic review. Neuro-Oncol. Adv. 2022, 4, vdac087. [Google Scholar] [CrossRef]
- González-Tablas Pimenta, M.; Otero, Á.; Arandia Guzman, D.A.; Pascual-Argente, D.; Ruíz Martín, L.; Sousa-Casasnovas, P.; García-Martin, A.; Roa Montes de Oca, J.C.; Villaseñor-Ledezma, J.; Torres Carretero, L.; et al. Tumor cell and immune cell profiles in primary human glioblastoma: Impact on patient outcome. Brain Pathol. 2021, 31, 365–380. [Google Scholar] [CrossRef]
- Khan, F.; Pang, L.; Dunterman, M.; Lesniak, M.S.; Heimberger, A.B.; Chen, P. Macrophages and microglia in glioblastoma: Heterogeneity, plasticity, and therapy. J. Clin. Investig. 2023, 133, e163446. [Google Scholar] [CrossRef]
- Butowski, N.; Colman, H.; de Groot, J.F.; Omuro, A.M.; Nayak, L.; Wen, P.Y.; Cloughesy, T.F.; Marimuthu, A.; Haidar, S.; Perry, A.; et al. Orally administered colony stimulating factor 1 receptor inhibitor PLX3397 in recurrent glioblastoma: An Ivy Foundation Early Phase Clinical Trials Consortium phase II study. Neuro-Oncology 2016, 18, 557–564. [Google Scholar] [CrossRef]
- Woolf, Z.; Swanson, M.E.V.; Smyth, L.C.; Mee, E.W.; Schweder, P.; Heppner, P.; Kim, B.J.H.; Turner, C.; Oldfield, R.L.; Curtis, M.A.; et al. Single-cell image analysis reveals a protective role for microglia in glioblastoma. Neuro-Oncol. Adv. 2021, 3, vdab031. [Google Scholar] [CrossRef]
- Ahmed, R.; Zaman, T.; Chowdhury, F.; Mraiche, F.; Tariq, M.; Ahmad, I.S.; Hasan, A. Single-Cell RNA Sequencing with Spatial Transcriptomics of Cancer Tissues. Int. J. Mol. Sci. 2022, 23, 3042. [Google Scholar] [CrossRef]
- Xuan, W.; Lesniak, M.S.; James, C.D.; Heimberger, A.B.; Chen, P. Context-Dependent Glioblastoma-Macrophage/Microglia Symbiosis and Associated Mechanisms. Trends Immunol. 2021, 42, 280–292. [Google Scholar] [CrossRef]
- Codrici, E.; Popescu, I.-D.; Tanase, C.; Enciu, A.-M. Friends with Benefits: Chemokines, Glioblastoma-Associated Microglia/Macrophages, and Tumor Microenvironment. Int. J. Mol. Sci. 2022, 23, 2509. [Google Scholar] [CrossRef]
- Pombo Antunes, A.R.; Scheyltjens, I.; Lodi, F.; Messiaen, J.; Antoranz, A.; Duerinck, J.; Kancheva, D.; Martens, L.; Vlaminck, K.; de van Hove, H.; et al. Single-cell profiling of myeloid cells in glioblastoma across species and disease stage reveals macrophage competition and specialization. Nat. Neurosci. 2021, 24, 595–610. [Google Scholar] [CrossRef]
- Andersen, J.K.; Miletic, H.; Hossain, J.A. Tumor-Associated Macrophages in Gliomas-Basic Insights and Treatment Opportunities. Cancers 2022, 14, 1319. [Google Scholar] [CrossRef]
- Acker, G.; Zollfrank, J.; Jelgersma, C.; Nieminen-Kelhä, M.; Kremenetskaia, I.; Mueller, S.; Ghori, A.; Vajkoczy, P.; Brandenburg, S. The CXCR2/CXCL2 signalling pathway—An alternative therapeutic approach in high-grade glioma. Eur. J. Cancer 2020, 126, 106–115. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, J.; Yao, X.; Wu, S.; Tian, W.; Gan, C.; Wan, X.; You, C.; Hu, F.; Zhang, S.; et al. Programmed Cell Death 10 Mediated CXCL2-CXCR2 Signaling in Regulating Tumor-Associated Microglia/Macrophages Recruitment in Glioblastoma. Front. Immunol. 2021, 12, 637053. [Google Scholar] [CrossRef]
- Urbantat, R.M.; Jelgersma, C.; Brandenburg, S.; Nieminen-Kelhä, M.; Kremenetskaia, I.; Zollfrank, J.; Mueller, S.; Rubarth, K.; Koch, A.; Vajkoczy, P.; et al. Tumor-Associated Microglia/Macrophages as a Predictor for Survival in Glioblastoma and Temozolomide-Induced Changes in CXCR2 Signaling with New Resistance Overcoming Strategy by Combination Therapy. Int. J. Mol. Sci. 2021, 22, 11180. [Google Scholar] [CrossRef] [PubMed]
- Urbantat, R.M.; Blank, A.; Kremenetskaia, I.; Vajkoczy, P.; Acker, G.; Brandenburg, S. The CXCL2/IL8/CXCR2 Pathway Is Relevant for Brain Tumor Malignancy and Endothelial Cell Function. Int. J. Mol. Sci. 2021, 22, 2634. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhao, Q.; Kong, L.-Y.; Wang, J.; Yan, J.; Xia, X.; Jia, Z.; Heimberger, A.B.; Li, S. Regulation of tumor immune suppression and cancer cell survival by CXCL1/2 elevation in glioblastoma multiforme. Sci. Adv. 2021, 7, eabc2511. [Google Scholar] [CrossRef] [PubMed]
- Urbantat, R.M.; Jelgersma, C.; Vajkoczy, P.; Brandenburg, S.; Acker, G. Combining TMZ and SB225002 induces changes of CXCR2 and VEGFR signalling in primary human endothelial cells in vitro. Oncol. Rep. 2022, 48, 158. [Google Scholar] [CrossRef] [PubMed]
- Urbantat, R.M.; Vajkoczy, P.; Brandenburg, S. Advances in Chemokine Signaling Pathways as Therapeutic Targets in Glioblastoma. Cancers 2021, 13, 2983. [Google Scholar] [CrossRef]
- Liu, Y.; Given, K.S.; Dickson, E.L.; Owens, G.P.; Macklin, W.B.; Bennett, J.L. Concentration-dependent effects of CSF1R inhibitors on oligodendrocyte progenitor cells ex vivo and in vivo. Exp. Neurol. 2019, 318, 32–41. [Google Scholar] [CrossRef]
- Coleman, L.G.; Zou, J.; Crews, F.T. Microglial depletion and repopulation in brain slice culture normalizes sensitized proinflammatory signaling. J. Neuroinflamm. 2020, 17, 27. [Google Scholar] [CrossRef]
- Sakuma, R.; Kawahara, M.; Nakano-Doi, A.; Takahashi, A.; Tanaka, Y.; Narita, A.; Kuwahara-Otani, S.; Hayakawa, T.; Yagi, H.; Matsuyama, T.; et al. Brain pericytes serve as microglia-generating multipotent vascular stem cells following ischemic stroke. J. Neuroinflamm. 2016, 13, 57. [Google Scholar] [CrossRef]
- Hutter, G.; Theruvath, J.; Graef, C.M.; Zhang, M.; Schoen, M.K.; Manz, E.M.; Bennett, M.L.; Olson, A.; Azad, T.D.; Sinha, R.; et al. Microglia are effector cells of CD47-SIRPα antiphagocytic axis disruption against glioblastoma. Proc. Natl. Acad. Sci. USA 2019, 116, 997–1006. [Google Scholar] [CrossRef]
- D’Alessio, A.; Proietti, G.; Sica, G.; Scicchitano, B.M. Pathological and Molecular Features of Glioblastoma and Its Peritumoral Tissue. Cancers 2019, 11, 469. [Google Scholar] [CrossRef]
- Böttcher, C.; Schlickeiser, S.; Sneeboer, M.A.M.; Kunkel, D.; Knop, A.; Paza, E.; Fidzinski, P.; Kraus, L.; Snijders, G.J.L.; Kahn, R.S.; et al. Human microglia regional heterogeneity and phenotypes determined by multiplexed single-cell mass cytometry. Nat. Neurosci. 2019, 22, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Afshari, A.R.; Sanati, M.; Aminyavari, S.; Shakeri, F.; Bibak, B.; Keshavarzi, Z.; Soukhtanloo, M.; Jalili-Nik, M.; Sadeghi, M.M.; Mollazadeh, H.; et al. Advantages and drawbacks of dexamethasone in glioblastoma multiforme. Crit. Rev. Oncol. Hematol. 2022, 172, 103625. [Google Scholar] [CrossRef] [PubMed]
- Dubinski, D.; Hattingen, E.; Senft, C.; Seifert, V.; Peters, K.G.; Reiss, Y.; Devraj, K.; Plate, K.H. Controversial roles for dexamethasone in glioblastoma—Opportunities for novel vascular targeting therapies. J. Cereb. Blood Flow Metab. 2019, 39, 1460–1468. [Google Scholar] [CrossRef] [PubMed]
- Park, M.-J.; Park, H.-S.; You, M.-J.; Yoo, J.; Kim, S.H.; Kwon, M.-S. Dexamethasone Induces a Specific Form of Ramified Dysfunctional Microglia. Mol. Neurobiol. 2019, 56, 1421–1436. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Xu, S.; Xin, Y.; Yu, S.-C.; Ping, Y.-F.; Chen, L.; Xiao, H.; Wang, B.; Yi, L.; Wang, Q.; et al. Tumor-associated microglia/macrophages enhance the invasion of glioma stem-like cells via TGF-β1 signaling pathway. J. Immunol. 2012, 189, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-W.E.; Lumibao, J.; Leary, S.; Sarkaria, J.N.; Steelman, A.J.; Gaskins, H.R.; Harley, B.A.C. Crosstalk between microglia and patient-derived glioblastoma cells inhibit invasion in a three-dimensional gelatin hydrogel model. J. Neuroinflamm. 2020, 17, 346. [Google Scholar] [CrossRef]
- Lanza, M.; Casili, G.; Campolo, M.; Paterniti, I.; Colarossi, C.; Mare, M.; Giuffrida, R.; Caffo, M.; Esposito, E.; Cuzzocrea, S. Immunomodulatory Effect of Microglia-Released Cytokines in Gliomas. Brain Sci. 2021, 11, 466. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Korbecki, J.; Grochans, S.; Gutowska, I.; Barczak, K.; Baranowska-Bosiacka, I. CC Chemokines in a Tumor: A Review of Pro-Cancer and Anti-Cancer Properties of Receptors CCR5, CCR6, CCR7, CCR8, CCR9, and CCR10 Ligands. Int. J. Mol. Sci. 2020, 21, 7619. [Google Scholar] [CrossRef]
- Martins, T.A.; Schmassmann, P.; Shekarian, T.; Boulay, J.-L.; Ritz, M.-F.; Zanganeh, S.; Vom Berg, J.; Hutter, G. Microglia-Centered Combinatorial Strategies Against Glioblastoma. Front. Immunol. 2020, 11, 571951. [Google Scholar] [CrossRef]
- Proescholdt, M.A.; Merrill, M.J.; Ikejiri, B.; Walbridge, S.; Akbasak, A.; Jacobson, S.; Oldfield, E.H. Site-specific immune response to implanted gliomas. J. Neurosurg. 2001, 95, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Stoppini, L.; Buchs, P.-A.; Muller, D. A simple method for organotypic cultures of nervous tissue. J. Neurosci. Methods 1991, 37, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Humpel, C. Organotypic brain slice cultures: A review. Neuroscience 2015, 305, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Spiteri, A.G.; King, N.J.C. Putting PLX5622 into perspective: Microglia in central nervous system viral infection. Neural Regen. Res. 2023, 18, 1269–1270. [Google Scholar]
- Czock, D.; Keller, F.; Rasche, F.M.; Häussler, U. Pharmacokinetics and pharmacodynamics of systemically administered glucocorticoids. Clin. Pharmacokinet. 2005, 44, 61–98. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falter, J.; Lohmeier, A.; Eberl, P.; Stoerr, E.-M.; Koskimäki, J.; Falter, L.; Rossmann, J.; Mederer, T.; Schmidt, N.O.; Proescholdt, M. CXCR2-Blocking Has Context-Sensitive Effects on Rat Glioblastoma Cell Line Outgrowth (S635) in an Organotypic Rat Brain Slice Culture Depending on Microglia-Depletion (PLX5622) and Dexamethasone Treatment. Int. J. Mol. Sci. 2023, 24, 16803. https://doi.org/10.3390/ijms242316803
Falter J, Lohmeier A, Eberl P, Stoerr E-M, Koskimäki J, Falter L, Rossmann J, Mederer T, Schmidt NO, Proescholdt M. CXCR2-Blocking Has Context-Sensitive Effects on Rat Glioblastoma Cell Line Outgrowth (S635) in an Organotypic Rat Brain Slice Culture Depending on Microglia-Depletion (PLX5622) and Dexamethasone Treatment. International Journal of Molecular Sciences. 2023; 24(23):16803. https://doi.org/10.3390/ijms242316803
Chicago/Turabian StyleFalter, Johannes, Annette Lohmeier, Petra Eberl, Eva-Maria Stoerr, Janne Koskimäki, Lena Falter, Jakob Rossmann, Tobias Mederer, Nils Ole Schmidt, and Martin Proescholdt. 2023. "CXCR2-Blocking Has Context-Sensitive Effects on Rat Glioblastoma Cell Line Outgrowth (S635) in an Organotypic Rat Brain Slice Culture Depending on Microglia-Depletion (PLX5622) and Dexamethasone Treatment" International Journal of Molecular Sciences 24, no. 23: 16803. https://doi.org/10.3390/ijms242316803
APA StyleFalter, J., Lohmeier, A., Eberl, P., Stoerr, E.-M., Koskimäki, J., Falter, L., Rossmann, J., Mederer, T., Schmidt, N. O., & Proescholdt, M. (2023). CXCR2-Blocking Has Context-Sensitive Effects on Rat Glioblastoma Cell Line Outgrowth (S635) in an Organotypic Rat Brain Slice Culture Depending on Microglia-Depletion (PLX5622) and Dexamethasone Treatment. International Journal of Molecular Sciences, 24(23), 16803. https://doi.org/10.3390/ijms242316803