

FARSB Facilitates Hepatocellular Carcinoma Progression by Activating the mTORC1 Signaling Pathway


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Result
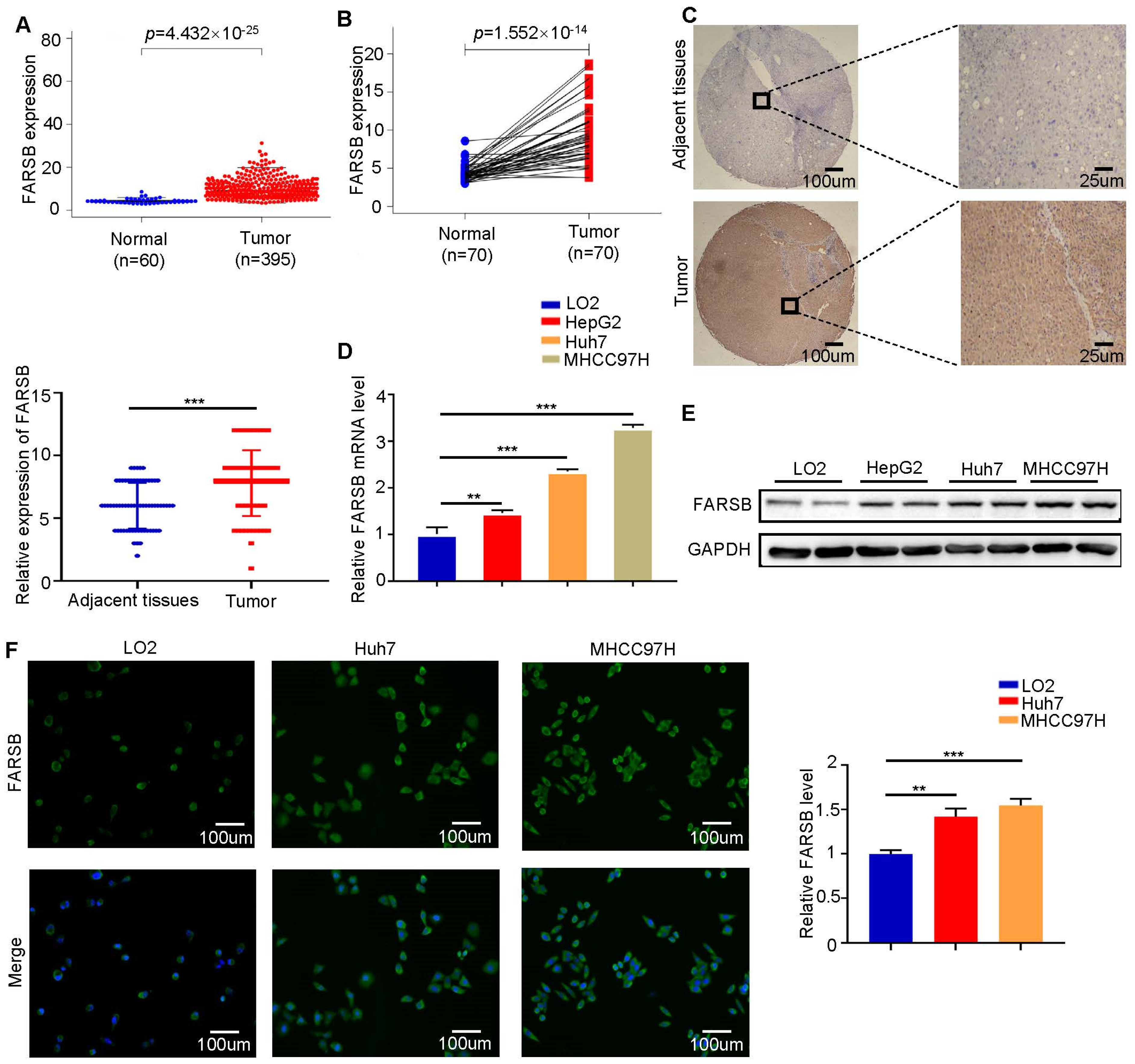
2.1. FARSB Is Highly Expressed in Hepatocellular Carcinoma
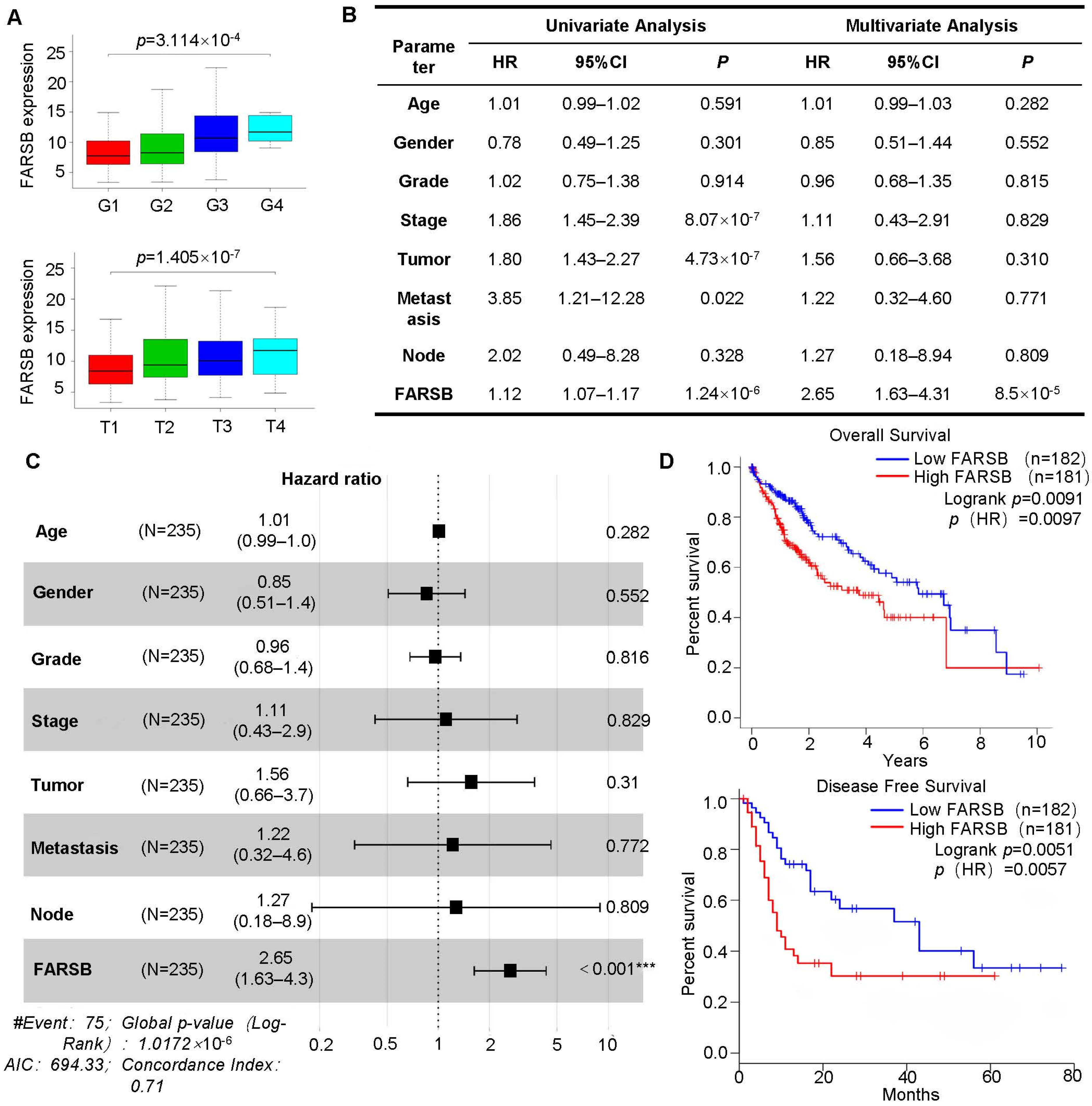
2.2. High Expression of FARSB Is Associated with Poor Prognosis of Patients
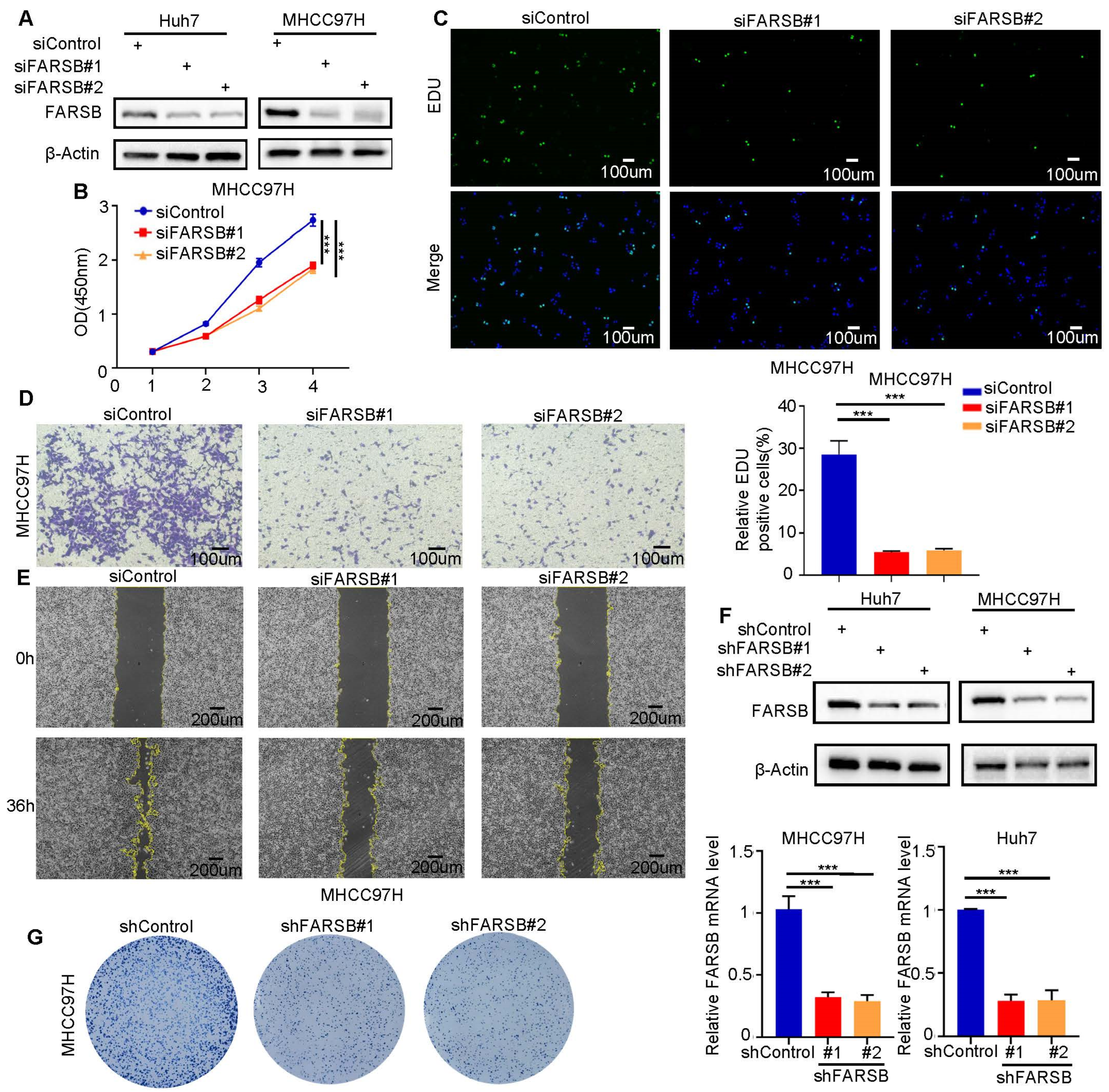
2.3. FARSB Knockdown Suppresses the Proliferation and Migration of HCC Cells
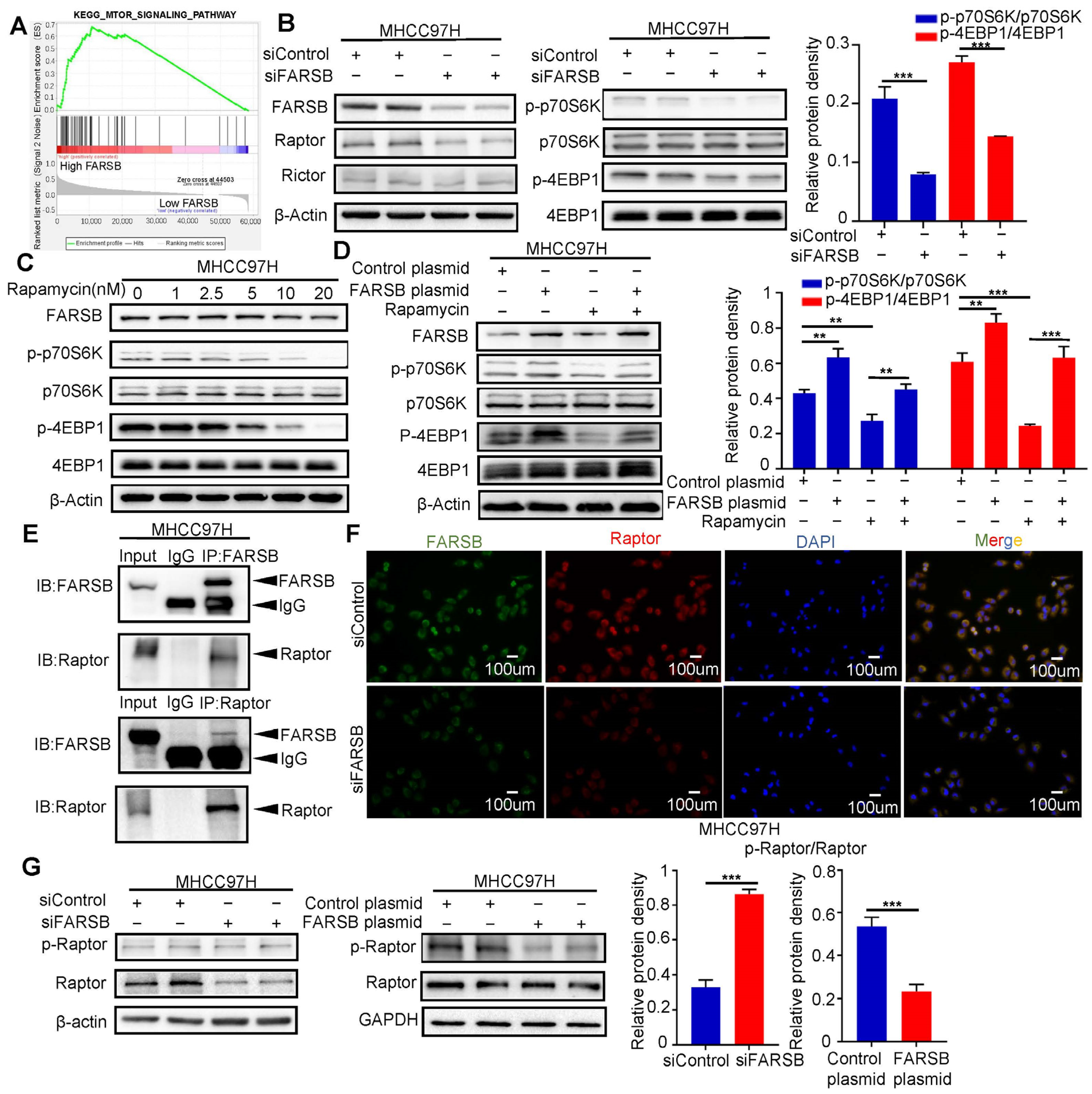
2.4. FARSB Activates the mTORC1 Signaling Pathway by Suppressing Raptor Phosphorylation
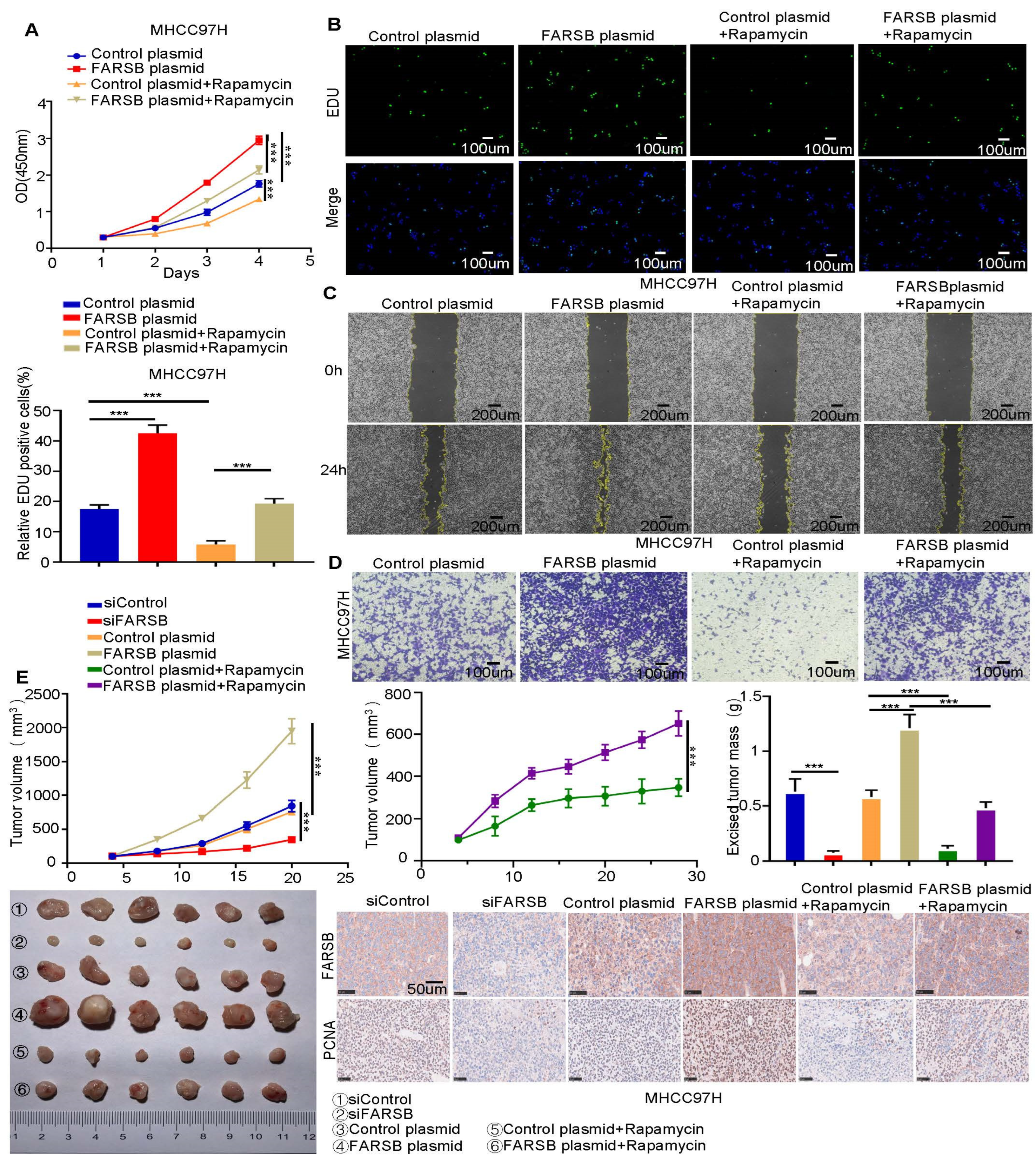
2.5. FARSB Promotes the Proliferation and Migration of HCC Cells by Activating the mTORC1 Signaling Pathway
2.6. FARSB Suppresses Ferroptosis in HCC Cells by Activating mTORC1 Expression
3. Discussion
4. Material and Methods
4.1. Bioinformatics Analysis
4.2. Patients and Tissue Specimens
4.3. Immunohistochemistry
4.4. Cell Culture
4.5. Immunofluorescence
4.6. Cell Transfection and Lentivirus Infection
4.7. Reverse Transcription and RT-PCR
4.8. Western Blot
4.9. Cell Proliferation Assay
4.10. Wound-Healing Assay
4.11. Transwell Assay
4.12. Colony Formation Assay
4.13. Co-IP
4.14. Xenograft Tumor Model in Nude Mice
4.15. ROS Detection
4.16. MDA Detection
4.17. Mitochondrial Electron Microscopy
4.18. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
| FARSB | Phenylalanyl-tRNA synthetase subunit beta |
| ARSs | aminoacyl-tRNA synthases |
| CysRS | cysteyl-trna synthetase |
| SerRS | seryl-tRNA synthetase |
| IleRS2 | isoleucyl-tRNA synthetase |
| HCC | hepatocellular carcinoma |
| GSEA | gene set enrichment analysis |
| Co-IP | Co-immunoprecipitation |
| CCK8 | Cell counting kit-8 |
| DMEM | Dulbecco’s modified eagle medium |
| RT-PCR | Quantitative real-time polymerase chain reaction |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| EdU | 5-Ethynyl-2′-deoxyuridine |
| 4EBP1 | 4E binding protein 1 |
| Fer-1 | Ferrostatin-1 |
| GPX4 | Glutathione peroxidase 4 |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| MDA | Malondialdehyde |
| mTOR | Mammalian target of rapamycin |
| mTORC1 | Mammalian target of rapamycin complex 1 |
| P70S6K | Phosphoprotein 70 ribosomal protein S6 kinase |
| PheRS | Phenylalanyl-tRNA synthetase |
| p-Raptor | Phosphorylated regulatory associated protein of mTOR |
| p-P70S6K | Phosphorylated phosphoprotein 70 ribosomal protein S6 kinase |
| p-4EBP1 | Phosphorylated 4E binding protein 1 |
| Raptor | Regulatory associated protein of mTOR |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Afra, F.; Mahboobipour, A.A.; Salehi Farid, A.; Ala, M. Recent progress in the immunotherapy of hepatocellular carcinoma: Non-coding RNA-based immunotherapy may improve the outcome. Biomed. Pharmacother. 2023, 165, 115104. [Google Scholar] [CrossRef] [PubMed]
- Vogel, A.; Meyer, T.; Sapisochin, G.; Salem, R.; Saborowski, A. Hepatocellular carcinoma. Lancet 2022, 400, 1345–1362. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.; Yoon, I.; Han, J.M.; Kim, S. Functional and pathologic association of aminoacyl-tRNA synthetases with cancer. Exp. Mol. Med. 2022, 54, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Feria, A.S.; Notarnicola, A.; Lundberg, I.E.; Horuluoglu, B. Aminoacyl-tRNA Synthetases: On Anti-Synthetase Syndrome and Beyond. Front. Immunol. 2022, 13, 866087. [Google Scholar] [CrossRef]
- Turvey, A.K.; Horvath, G.A.; Cavalcanti, A.R.O. Aminoacyl-tRNA synthetases in human health and disease. Front. Physiol. 2022, 13, 1029218. [Google Scholar] [CrossRef]
- Zhou, Z.; Sun, B.; Nie, A.; Yu, D.; Bian, M. Roles of Aminoacyl-tRNA Synthetases in Cancer. Front. Cell Dev. Biol. 2020, 8, 599765. [Google Scholar] [CrossRef]
- Ho, M.T.; Lu, J.; Brunßen, D.; Suter, B. A translation-independent function of PheRS activates growth and proliferation in Drosophila. Dis. Model Mech. 2021, 14, dmm048132. [Google Scholar] [CrossRef]
- Fan, W.; Jin, X.; Xu, M.; Xi, Y.; Lu, W.; Yang, X.; Guan, M.X.; Ge, W. FARS2 deficiency in Drosophila reveals the developmental delay and seizure manifested by aberrant mitochondrial tRNA metabolism. Nucleic Acids Res. 2021, 49, 13108–13121. [Google Scholar] [CrossRef]
- Zadjali, F.; Al-Yahyaee, A.; Al-Nabhani, M.; Al-Mubaihsi, S.; Gujjar, A.; Raniga, S.; Al-Maawali, A. Homozygosity for FARSB mutation leads to Phe-tRNA synthetase-related disease of growth restriction, brain calcification, and interstitial lung disease. Hum. Mutat. 2018, 39, 1355–1359. [Google Scholar] [CrossRef]
- Karimzadeh, P.; Rezakhani, S.; Miryounesi, M.; Alijanpour, S. Neurodegenerative disorder and diffuse brain calcifications due to FARSB mutation in two siblings. Clin. Case Rep. 2022, 10, e6195. [Google Scholar] [CrossRef]
- Xu, Z.; Lo, W.S.; Beck, D.B.; Schuch, L.A.; Oláhová, M.; Kopajtich, R.; Chong, Y.E.; Alston, C.L.; Seidl, E.; Zhai, L.; et al. Bi-allelic Mutations in Phe-tRNA Synthetase Associated with a Multi-system Pulmonary Disease Support Non-translational Function. Am. J. Hum. Genet. 2018, 103, 100–114. [Google Scholar] [CrossRef]
- Ruan, W.; Wang, Y.; Ma, Y.; Xing, X.; Lin, J.; Cui, J.; Lai, M. HSP60, a protein downregulated by IGFBP7 in colorectal carcinoma. J. Exp. Clin. Cancer Res. 2010, 29, 41. [Google Scholar] [CrossRef]
- Dong, Y.M.; Li, M.; He, Q.E.; Tong, Y.F.; Gao, H.Z.; Zhang, Y.Z.; Wu, Y.M.; Hu, J.; Zhang, N.; Song, K. Epigenome-Wide Tobacco-Related Methylation Signature Identification and Their Multilevel Regulatory Network Inference for Lung Adenocarcinoma. BioMed Res. Int. 2020, 2020, 2471915. [Google Scholar] [CrossRef]
- Zhen, J.; Pan, J.; Zhou, X.; Yu, Z.; Jiang, Y.; Gong, Y.; Ding, Y.; Liu, Y.; Guo, L. FARSB serves as a novel hypomethylated and immune cell infiltration related prognostic biomarker in hepatocellular carcinoma. Aging 2023, 15, 2937–2969. [Google Scholar] [CrossRef] [PubMed]
- Szwed, A.; Kim, E.; Jacinto, E. Regulation and metabolic functions of mTORC1 and mTORC2. Physiol. Rev. 2021, 101, 1371–1426. [Google Scholar] [CrossRef] [PubMed]
- Hua, H.; Kong, Q.; Zhang, H.; Wang, J.; Luo, T.; Jiang, Y. Targeting mTOR for cancer therapy. J. Hematol. Oncol. 2019, 12, 71. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.; Sonenberg, N.; Gores, G.J. The mTOR pathway in hepatic malignancies. Hepatology 2013, 58, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Paliogiannis, P.; Calvisi, D.F.; Chen, X. Role of the Mammalian Target of Rapamycin Pathway in Liver Cancer: From Molecular Genetics to Targeted Therapies. Hepatology 2021, 73, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.T.; White, R.R.; Liu, J.X.; Liu, H.Y. Seryl-tRNA synthetase-mediated essential amino acids regulate β-casein synthesis via cell proliferation and mammalian target of rapamycin (mTOR) signaling pathway in bovine mammary epithelial cells. J. Dairy Sci. 2018, 101, 10456–10468. [Google Scholar] [CrossRef]
- Di, X.; Jin, X.; Ma, H.; Wang, R.; Cong, S.; Tian, C.; Liu, J.; Zhao, M.; Li, R.; Wang, K. The Oncogene IARS2 Promotes Non-small Cell Lung Cancer Tumorigenesis by Activating the AKT/MTOR Pathway. Front. Oncol. 2019, 9, 393. [Google Scholar] [CrossRef]
- Chen, X.; Kang, R.; Kroemer, G.; Tang, D. Broadening horizons: The role of ferroptosis in cancer. Nat. Rev. Clin. Oncol. 2021, 18, 280–296. [Google Scholar] [CrossRef]
- Chen, X.; Li, J.; Kang, R.; Klionsky, D.J.; Tang, D. Ferroptosis: Machinery and regulation. Autophagy 2021, 17, 2054–2081. [Google Scholar] [CrossRef]
- Hayano, M.; Yang, W.S.; Corn, C.K.; Pagano, N.C.; Stockwell, B.R. Loss of cysteinyl-tRNA synthetase (CARS) induces the transsulfuration pathway and inhibits ferroptosis induced by cystine deprivation. Cell Death Differ. 2016, 23, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lin, X.; Chen, S. Cysteinyl-tRNA Synthetase 1 Promotes Ferroptosis-Induced Cell Death via Regulating GPX4 Expression. J. Oncol. 2022, 2022, 4849174. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Zhu, J.; Wu, J.; Thompson, C.B.; Jiang, X. Oncogenic activation of PI3K-AKT-mTOR signaling suppresses ferroptosis via SREBP-mediated lipogenesis. Proc. Natl. Acad. Sci. USA 2020, 117, 31189–31197. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jung, K.; Lee, C.; Song, D.; Kim, K.; Yoo, H.C.; Park, S.J.; Kang, J.S.; Lee, K.R.; Kim, S.; et al. Structure-based modification of pyrazolone derivatives to inhibit mTORC1 by targeting the leucyl-tRNA synthetase-RagD interaction. Bioorganic Chem. 2021, 112, 104907. [Google Scholar] [CrossRef]
- Arif, A.; Terenzi, F.; Potdar, A.A.; Jia, J.; Sacks, J.; China, A.; Halawani, D.; Vasu, K.; Li, X.; Brown, J.M.; et al. EPRS is a critical mTORC1-S6K1 effector that influences adiposity in mice. Nature 2017, 542, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Tan, J.; Xiong, W.; Chen, S.; Fan, L.; Li, Y. Notch3 promotes 3T3-L1 pre-adipocytes differentiation by up-regulating the expression of LARS to activate the mTOR pathway. J. Cell Mol. Med. 2020, 24, 1116–1127. [Google Scholar] [CrossRef]
- Gwinn, D.M.; Shackelford, D.B.; Egan, D.F.; Mihaylova, M.M.; Mery, A.; Vasquez, D.S.; Turk, B.E.; Shaw, R.J. AMPK phosphorylation of raptor mediates a metabolic checkpoint. Mol. Cell. 2008, 30, 214–226. [Google Scholar] [CrossRef]
- Francklyn, C.S.; Mullen, P. Progress and challenges in aminoacyl-tRNA synthetase-based therapeutics. J. Biol. Chem. 2019, 294, 5365–5385. [Google Scholar] [CrossRef]
- Han, J.M.; Jeong, S.J.; Park, M.C.; Kim, G.; Kwon, N.H.; Kim, H.K.; Ha, S.H.; Ryu, S.H.; Kim, S. Leucyl-tRNA synthetase is an intracellular leucine sensor for the mTORC1-signaling pathway. Cell 2012, 149, 410–424. [Google Scholar] [CrossRef]
- Howell, J.J.; Manning, B.D. mTOR couples cellular nutrient sensing to organismal metabolic homeostasis. Trends Endocrinol. Metab. 2011, 22, 94–102. [Google Scholar] [CrossRef]
- Zeng, T.; Zhou, Y.; Yu, Y.; Wang, J.W.; Wu, Y.; Wang, X.; Zhu, L.; Zhou, L.M.; Wan, L.H. rmMANF prevents sepsis-associated lung injury via inhibiting endoplasmic reticulum stress-induced ferroptosis in mice. Int. Immunopharmacol. 2023, 114, 109608. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wang, G.; Hu, S.; Yin, C.; Zhao, P.; Zhou, X.; Shao, S.; Liu, R.; Hu, W.; Liu, G.L.; et al. FARSB Facilitates Hepatocellular Carcinoma Progression by Activating the mTORC1 Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 16709. https://doi.org/10.3390/ijms242316709
Wang Y, Wang G, Hu S, Yin C, Zhao P, Zhou X, Shao S, Liu R, Hu W, Liu GL, et al. FARSB Facilitates Hepatocellular Carcinoma Progression by Activating the mTORC1 Signaling Pathway. International Journal of Molecular Sciences. 2023; 24(23):16709. https://doi.org/10.3390/ijms242316709
Chicago/Turabian StyleWang, Yaofeng, Gengqiao Wang, Shaobo Hu, Chuanzheng Yin, Peng Zhao, Xing Zhou, Shuyu Shao, Ran Liu, Wenjun Hu, Gang Logan Liu, and et al. 2023. "FARSB Facilitates Hepatocellular Carcinoma Progression by Activating the mTORC1 Signaling Pathway" International Journal of Molecular Sciences 24, no. 23: 16709. https://doi.org/10.3390/ijms242316709
APA StyleWang, Y., Wang, G., Hu, S., Yin, C., Zhao, P., Zhou, X., Shao, S., Liu, R., Hu, W., Liu, G. L., Ke, W., & Song, Z. (2023). FARSB Facilitates Hepatocellular Carcinoma Progression by Activating the mTORC1 Signaling Pathway. International Journal of Molecular Sciences, 24(23), 16709. https://doi.org/10.3390/ijms242316709