
Akt, IL-4, and STAT Proteins Play Distinct Roles in Prostaglandin Production in Human Follicular Dendritic Cell-like Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
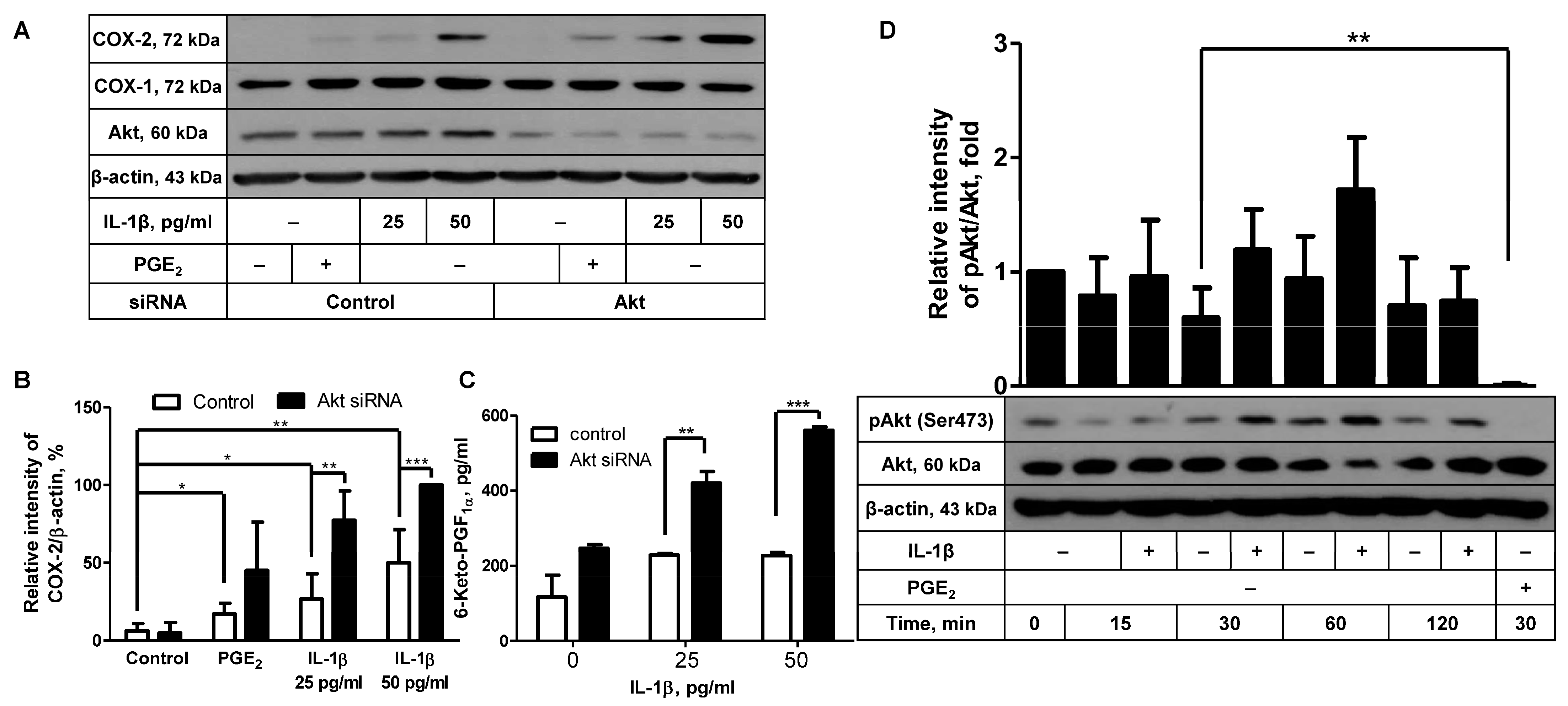
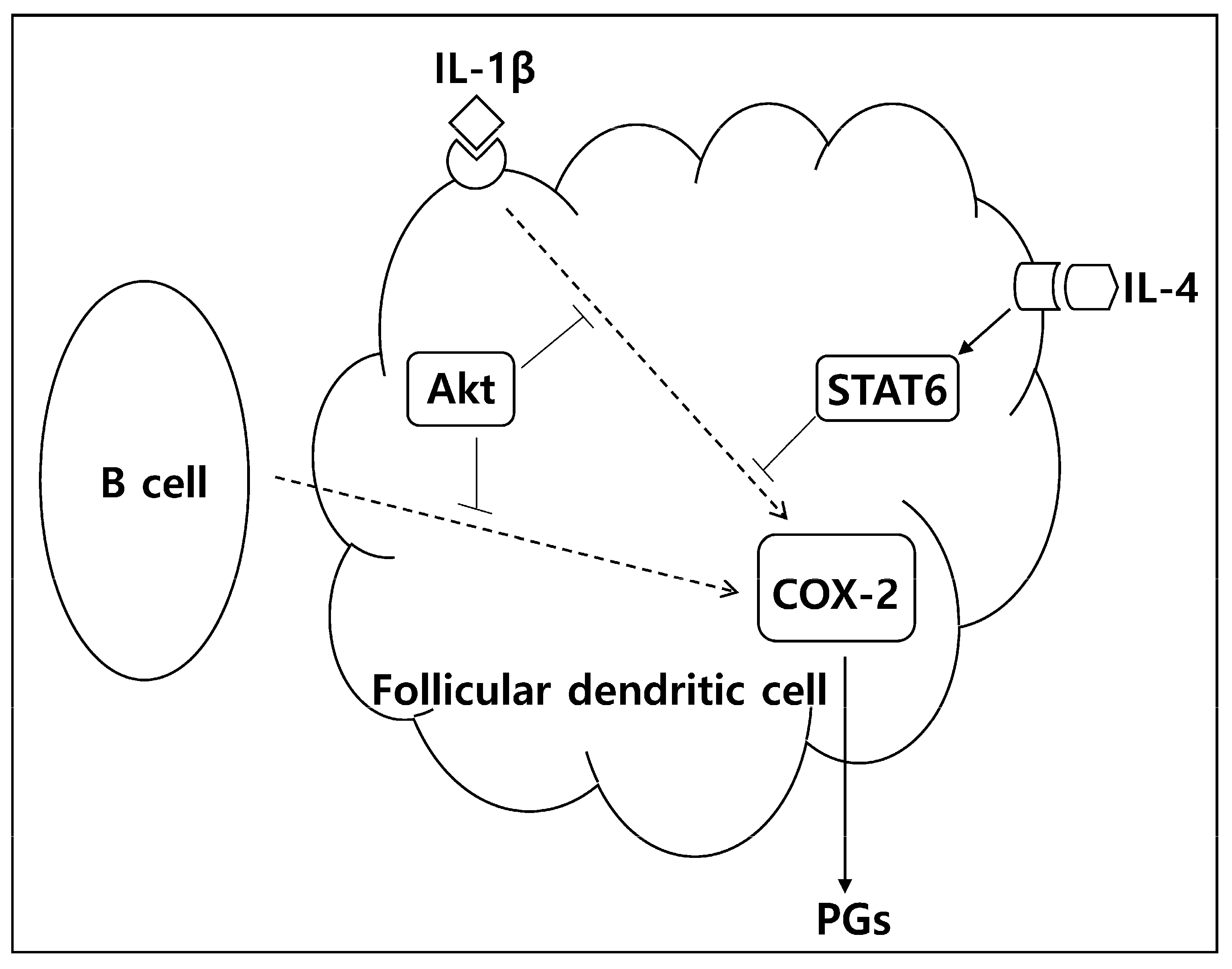
2.1. Akt Knockdown Leads to Augmentation of IL-1β-Induced COX-2 Expression
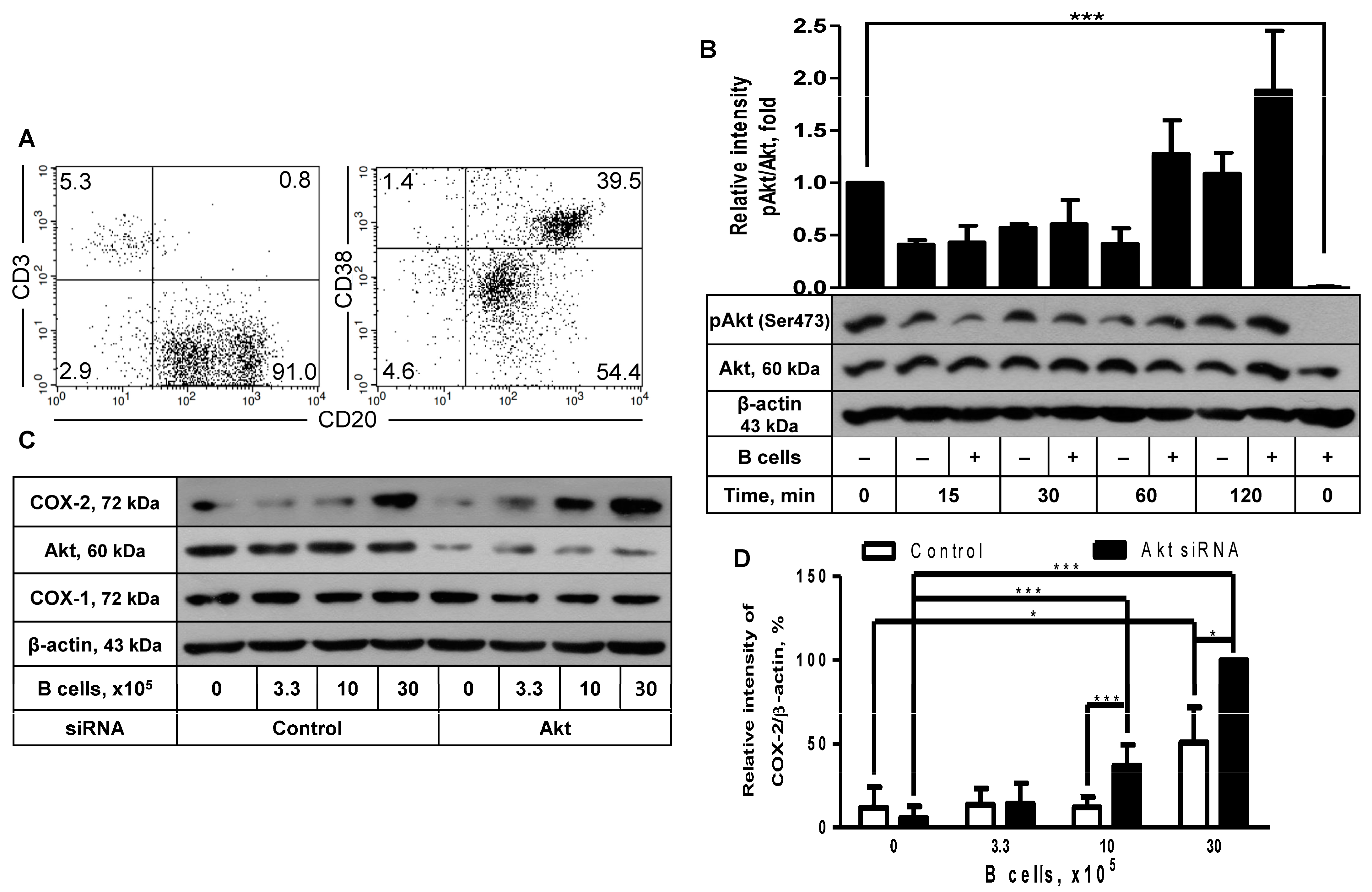
2.2. The Inhibitory Effect of Akt Was Also Observed in COX-2 Induction after Coculture with B Cells
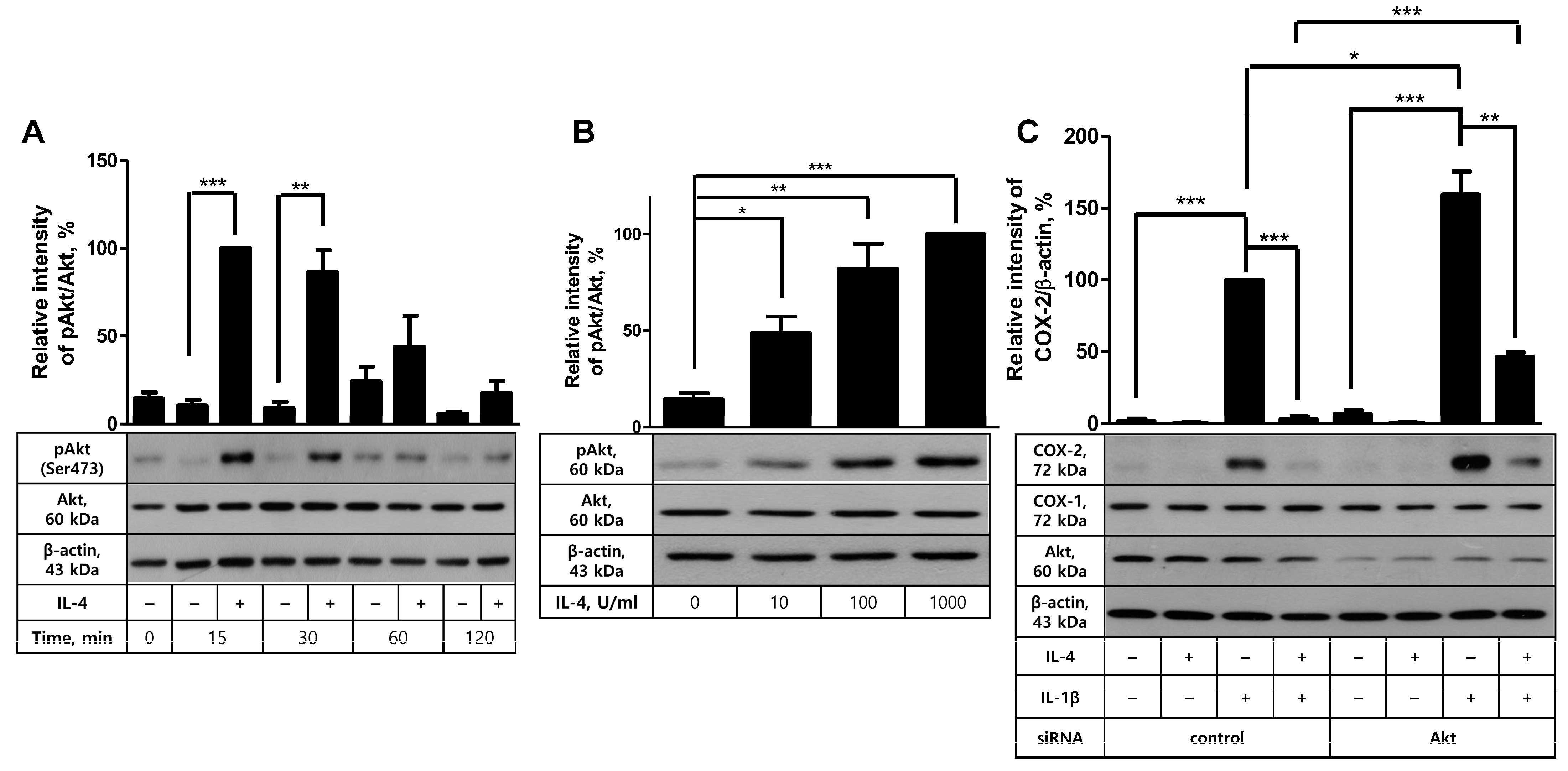
2.3. Akt Is Not Involved in IL-4-Mediated Inhibition of COX-2 Induction in FDC-like Cells
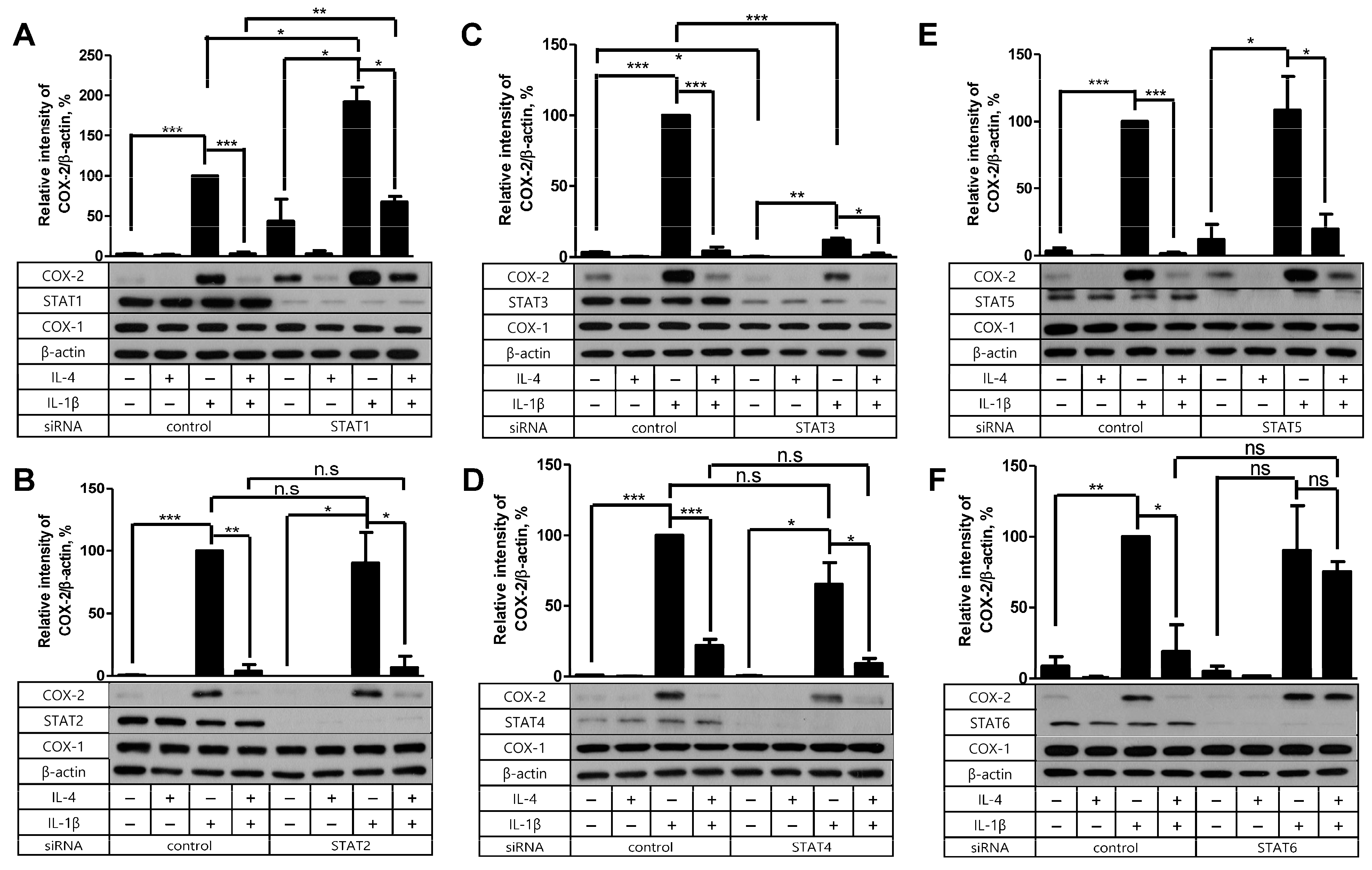
2.4. STAT6 Is Necessary for IL-4 to Suppress the IL-1β-Driven COX-2 Expression
3. Discussion
4. Material and Methods
4.1. Reagents
4.2. Isolation of FDC-like Cells and B Cells
4.3. Enzyme Immunoassay to Measure PGI2 (6-keto-PGF1α)
4.4. siRNA Transfection
4.5. Immunoblotting
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- El Shikh, M.E.M.; Pitzalis, C. Follicular dendritic cells in health and disease. Front. Immunol. 2012, 3, 292. [Google Scholar] [CrossRef]
- Munoz-Fernandez, R.; Blanco, F.J.; Frecha, C.; Martin, F.; Kimatrai, M.; Abadia-Molina, A.C.; Garcia-Pacheco, J.M.; Olivares, E.G. Follicular dendritic cells are related to bone marrow stromal cell progenitors and to myofibroblasts. J. Immunol. 2006, 177, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Aydar, Y.; Wu, J.; Song, J.; Szakal, A.K.; Tew, J.G. FcγRII expression on follicular dendritic cells and immunoreceptor tyrosine-based inhibition motif signaling in B cells. Eur. J. Immunol. 2004, 34, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Chen, X.; Hase, K.; Sakamoto, A.; Nishigaki, C.; Ohno, H. Splenic CD19-CD35+B220+ cells function as an inducer of follicular dendritic cell network formation. Blood 2007, 110, 1215–1224. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-J.; Xu, J.; de Bouteiller, O.; Parham, C.L.; Grouard, G.; Djossou, O.; de Saint-Vis, B.; Lebecque, S.; Banchereau, J.; Moore, K.W. Follicular dendritic cells specifically express the long CR2/CD21 isoform. J. Exp. Med. 1997, 185, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Vermi, W.; Lonardi, S.; Bosisio, D.; Uguccioni, M.; Danelon, G.; Pileri, S.; Fletcher, C.; Sozzani, S.; Zorzi, F.; Arrigoni, G.; et al. Identification of CXCL13 as a new marker for follicular dendritic cell sarcoma. J. Pathol. 2008, 216, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Heinen, E.; Cormann, N.; Braun, M.; Kinet-Denoel, C.; Vanderschelden, J.; Simar, L.J. Isolation of follicular dendritic cells from human tonsils and adenoids. VI. Analysis of prostaglandin secretion. Ann. Inst. Pasteur Immunol. 1986, 137D, 369–382. [Google Scholar]
- Clark, E.A.; Grabstein, K.H.; Shu, G.L. Cultured human follicular dendritic cells: Growth characteristics and interactions with B lymphocytes. J. Immunol. 1992, 148, 3327–3335. [Google Scholar] [CrossRef]
- Lee, I.Y.; Ko, E.-M.; Kim, S.-H.; Jeoung, D.-I.; Choe, J. Human follicular dendritic cells express prostacyclin synthase: A novel mechanism to control T cell numbers in the germinal center. J. Immunol. 2005, 175, 1658–1664. [Google Scholar] [CrossRef]
- Kim, J.; Kim, Y.-M.; Jeoung, D.-I.; Choe, J. Human follicular dendritic cells promote the APC capability of B cells by enhancing CD86 expression levels. Cell. Immunol. 2012, 273, 109–114. [Google Scholar] [CrossRef]
- Kim, J.; Lee, S.; Jeoung, D.; Kim, Y.-M.; Choe, J. Activated human B cells stimulate COX-2 expression in follicular dendritic cell-like cells via TNF-α. Mol. Immunol. 2018, 94, 1–6. [Google Scholar] [CrossRef]
- Cho, W.; Jeoung, D.; Kim, Y.-M.; Choe, J. STAT6 and JAK1 are essential for IL-4-mediated suppression of prostaglandin production in human follicular dendritic cells: Opposing roles of phosphorylated and unphosphorylated STAT6. Int. Immunopharmacol. 2012, 12, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Kim, J.; Kim, Y.-M.; Ha, K.S.; Jeoung, D.-I.; Sin, J.-I.; Choe, J. Beraprost enhances production of antigen-specific IgG isotypes without modulating germinal center B cell generation and the affinity maturation. Int. Immunopharmacol. 2013, 15, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Bernard, M.P.; Bancos, S.; Chapman, T.J.; Ryan, E.P.; Treanor, J.J.; Rose, R.C.; Topham, D.J.; Phipps, R.P. Chronic inhibition of cyclooxygenase-2 attenuates antibody responses against vaccinia infection. Vaccine 2010, 28, 1363–1372. [Google Scholar] [CrossRef]
- Kojima, F.; Frolov, A.; Matnani, R.; Woodward, J.G.; Crofford, L.J. Reduced T cell-dependent humoral immune response in microsomal prostaglandin E synthase-1 null mice is mediated by nonhematopoietic cells. J. Immunol. 2013, 191, 4979–4988. [Google Scholar] [CrossRef]
- Manning, B.D.; Toker, A. AKT/PKB signaling: Navigating the network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef]
- Choe, J.; Yoon, Y.; Kim, J.; Jung, Y.-J. Positive feedback effect of PGE2 on cyclooxygenase-2 expression is mediated by inhibition of Akt phosphorylation in human follicular dendritic cell-like cells. Mol. Immunol. 2017, 87, 60–66. [Google Scholar] [CrossRef]
- Ritvo, P.-G.; Klatzmann, D. Interleukin-1 in the response of follicular helper and follicular regulatory T cells. Front. Immunol. 2019, 10, 250. [Google Scholar] [CrossRef]
- Butch, A.W.; Chung, G.-H.; Hoffman, J.W.; Nahm, M.H. Cytokine expression by germinal center cells. J. Immunol. 1993, 150, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Goval, J.-J.; Greimers, R.; Boniver, J.; de Leval, L. Germinal center dendritic cells express more ICAM-1 than extrafollicular dendritic cells and ICAM-1/LFA-1 interactions are involved in the capacity of dendritic cells to induce PBMCs proliferation. J. Histochem. Cytochem. 2006, 54, 75–84. [Google Scholar] [CrossRef]
- Mantovani, A.; Dinarello, C.A.; Molgora, M.; Garlanda, C. IL-1 and related cytokines in innate and adaptive immunity in health and disease. Immunity 2019, 50, 778–795. [Google Scholar] [CrossRef]
- Heinen, E.; Bosseloir, A. Follicular dendritic cells: Whose children? Immunol. Today 1994, 15, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Kaji, M.; Takagashi, T.; Berg, K.; Dijkstra, C.D. Host origin of follicular dendritic cells induced in the spleen of SCID mice after transfer of allogeneic lymphocytes. Immunology 1995, 84, 117–126. [Google Scholar] [PubMed]
- Yao, C.; Narumiya, S. Prostaglandin-cytokine crosstalk in chronic inflammation. Br. J. Pharmacol. 2019, 176, 337–354. [Google Scholar] [CrossRef]
- Harris, S.G.; Padilla, J.; Koumas, L.; Ray, D.; Phipps, R.P. Prostaglandins as modulators of immunity. Trends Immunol. 2002, 23, 144–150. [Google Scholar] [CrossRef]
- Mandal, A.K.; Zhang, Z.; Kim, S.-J.; Tsai, P.-C.; Mukherjee, A.B. Yin-Yang: Balancing act of prostaglandins with opposing functions to regulate inflammation. J. Immunol. 2005, 175, 6271–6273. [Google Scholar] [CrossRef]
- Takeda, K.; Tanaka, T.; Shi, W.; Matsumoto, M.; Minami, M.; Kashiwamura, S.-I.; Nakanishi, K.; Yoshida, N.; Kishimoto, T.; Akira, S. Essential role of Stat6 in IL-4 signalling. Nature 1996, 380, 627–630. [Google Scholar] [CrossRef]
- Müller-Ladner, U.; Judex, M.; Ballhorn, W.; Kullmann, F.; Distler, O.; Scholmerich, J.; Gay, S. Activation of the IL-4 STAT pathway in rheumatoid synovium. J. Immunol. 2000, 164, 3894–3901. [Google Scholar] [CrossRef]
- Duan, L.; Liu, D.; Chen, H.; Mintz, M.A.; Chou, M.Y.; Kotov, D.I.; Xu, Y.; An, J.; Laidlaw, B.J.; Cyster, J.G. Follicular dendritic cells restrict interleukin-4 availability in germinal centers and foster memory B cell generation. Immunity 2021, 54, 2256–2272. [Google Scholar] [CrossRef]
- Li, X.; Han, Z.; Wang, F.; Qiao, J. The STAT6 inhibitor AS1517499 reduces the risk of asthma in mice with 2,4-dinitrochlorobenzene-induced atopic dermatitis by blocking the STAT6 signaling pathway. Allergy Asthma Clin. Immunol. 2022, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-E.; Jung, K.; Kim, J.-A.; Kim, S.-H.; Park, H.-S.; Kim, Y.-S. Engineering of anti-human interleukin-4 receptor alpha antibodies with potent antagonistic activity. Sci. Rep. 2019, 9, 7772. [Google Scholar] [CrossRef]
- Shirakawa, T.; Kawazoe, Y.; Tsujikawa, T.; Jung, D.; Sato, S.; Uesugi, M. Deactivation of STAT6 through serine 707 phosphorylation by JNK. J. Biol. Chem. 2011, 286, 4003–4010. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Choi, I.H.; Lee, D.G.; Han, S.S.; Ha, T.Y.; Lee, J.H.; Lee, W.H.; Park, Y.M.; Lee, H.K. Anti-IL-4 monoclonal antibody prevents antibiotics-induced active fatal anaphylaxis. J. Immunol. 1997, 158, 5002–5006. [Google Scholar] [CrossRef] [PubMed]
- Hua, H.; Zhang, H.; Chen, J.; Wang, J.; Liu, J.; Jiang, Y. Targeting Akt in cancer for precision therapy. J. Hematol. Oncol. 2021, 14, 128. [Google Scholar] [CrossRef]
- Adefemi, F.; Fruman, D.A.; Marshall, A.J. A case for phosphoinositide 3-kinase-targeted therapy for infectious disease. J. Immunol. 2020, 205, 3237–3245. [Google Scholar] [CrossRef] [PubMed]
- Nitulescu, G.M.; Margina, D.; Juzenas, P.; Peng, Q.; Olaru, O.T.; Saloustros, E.; Fenga, C.; Spandidos, D.A.; Libra, M.; Tsatsakis, A.M. Akt inhibitors in cancer treatment: The long journey from drug discovery to clinical use. Int. J. Oncol. 2016, 48, 869–885. [Google Scholar] [CrossRef]
- Husson, H.; Lugli, S.M.; Ghia, P.; Cardoso, A.; Roth, A.; Brohmi, K.; Carideo, E.G.; Choi, Y.S.; Browning, J.; Freedman, A.S. Functional effects of TNF and lymphotoxin α1β2 on FDC-like cells. Cell. Immunol. 2000, 203, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Bernard, A.; Gelin, C.; Raynal, B.; Pham, D.; Gosse, C.; Boumsell, L. Phenomenon of human T cells resetting with sheep erythrocytes analyzed with monoclonal antibodies. “Modulation” of a partially hidden epitope determining the conditions of interaction between T cells and erythrocytes. J. Exp. Med. 1982, 155, 1317–1333. [Google Scholar] [CrossRef]
- Shinmura, K.; Tang, X.-L.; Wang, Y.; Xuan, Y.-T.; Liu, S.-Q.; Takano, H.; Bhatnagar, A.; Bolli, R. Cyclooxygenase-2 mediates the cardioprotective effects of the late phase of ischemic preconditioning in conscious rabbits. Proc. Natl. Acad. Sci. USA 2000, 97, 10197–10202. [Google Scholar] [CrossRef]
- Taylor, S.C.; Rosselli-Murai, L.K.; Crobeddu, B.; Plante, I. A critical path to producing high quality, reproducible data from quantitative western blot experiments. Sci. Rep. 2022, 12, 17599. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, J.; Choe, J. Akt, IL-4, and STAT Proteins Play Distinct Roles in Prostaglandin Production in Human Follicular Dendritic Cell-like Cells. Int. J. Mol. Sci. 2023, 24, 16692. https://doi.org/10.3390/ijms242316692
Jeong J, Choe J. Akt, IL-4, and STAT Proteins Play Distinct Roles in Prostaglandin Production in Human Follicular Dendritic Cell-like Cells. International Journal of Molecular Sciences. 2023; 24(23):16692. https://doi.org/10.3390/ijms242316692
Chicago/Turabian StyleJeong, Jihye, and Jongseon Choe. 2023. "Akt, IL-4, and STAT Proteins Play Distinct Roles in Prostaglandin Production in Human Follicular Dendritic Cell-like Cells" International Journal of Molecular Sciences 24, no. 23: 16692. https://doi.org/10.3390/ijms242316692
APA StyleJeong, J., & Choe, J. (2023). Akt, IL-4, and STAT Proteins Play Distinct Roles in Prostaglandin Production in Human Follicular Dendritic Cell-like Cells. International Journal of Molecular Sciences, 24(23), 16692. https://doi.org/10.3390/ijms242316692