
The Plasma Membrane Purinoreceptor P2K1/DORN1 Is Essential in Stomatal Closure Evoked by Extracellular Diadenosine Tetraphosphate (Ap4A) in Arabidopsis thaliana

 , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
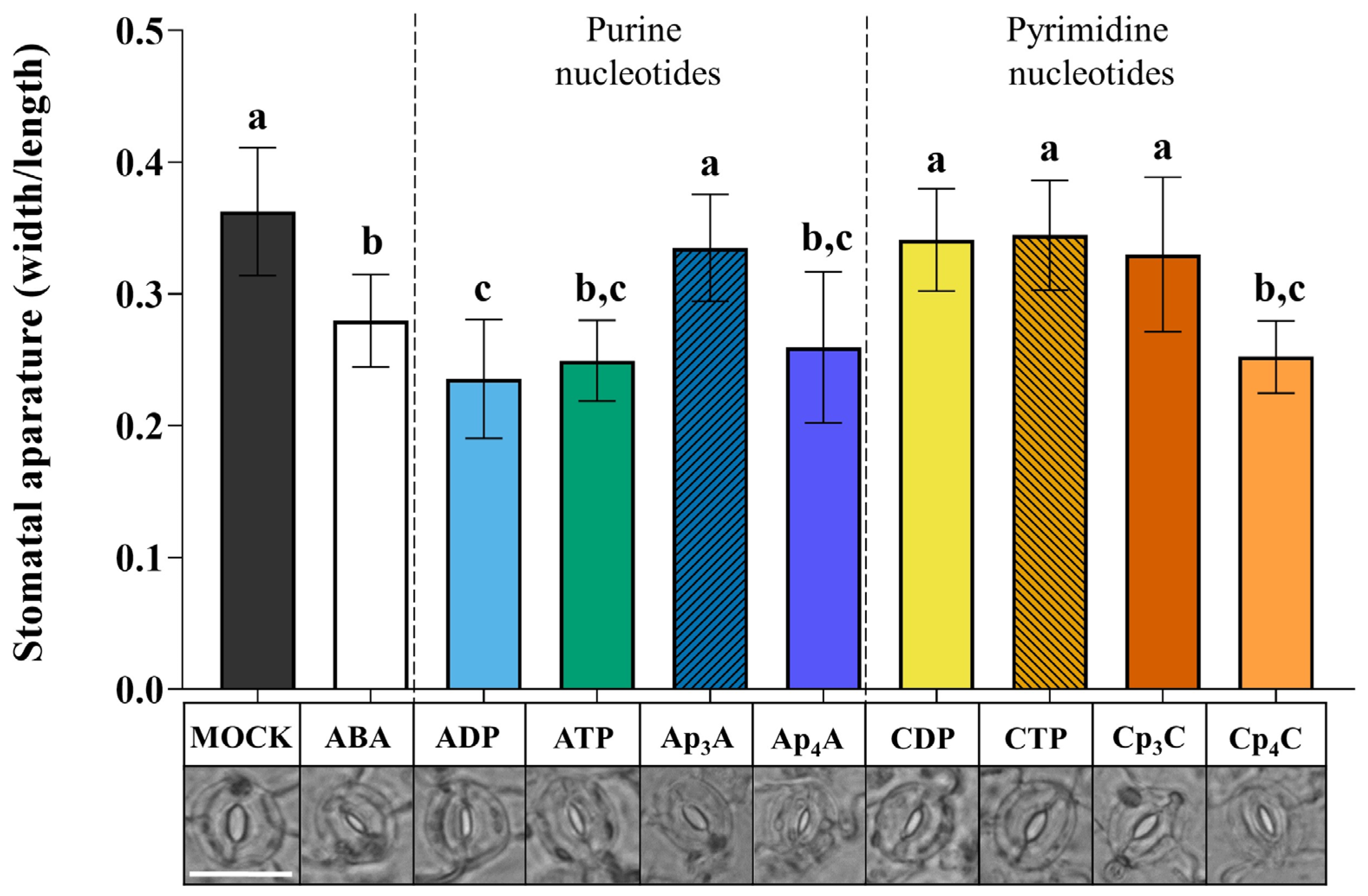
2.1. Ap4A and Cp4C Induce Stomatal Closure
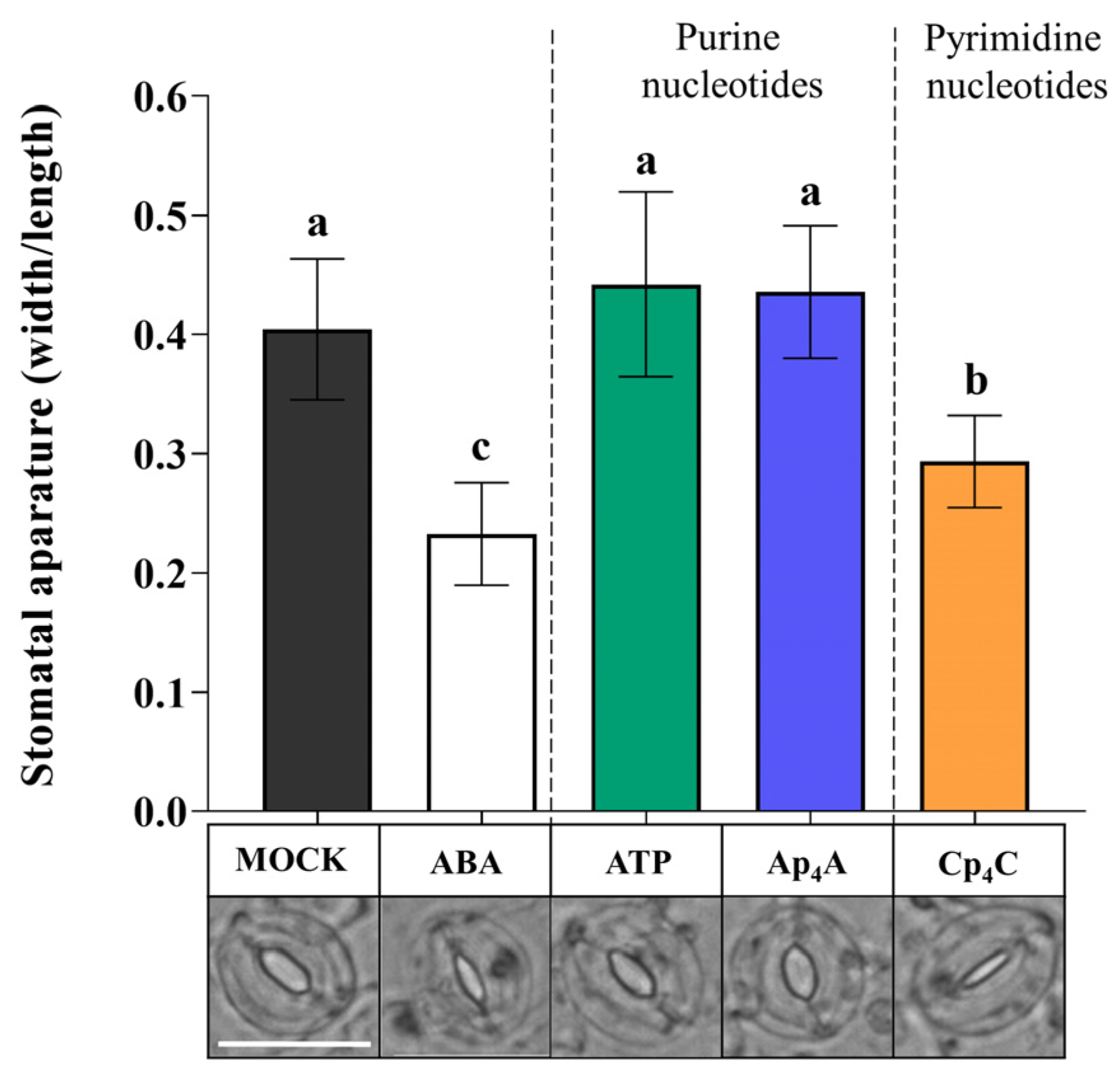
2.2. P2K1/DORN1 Is Involved in Signal Perception Evoked by Ap4A but Not Cp4C
2.3. ROS Are Produced in Leaves under Nucleotide Treatment
2.4. ROS Are Involved in Signal Transduction Evoked by eATP, Ap4A and Cp4C, Leading to Stomatal Closure
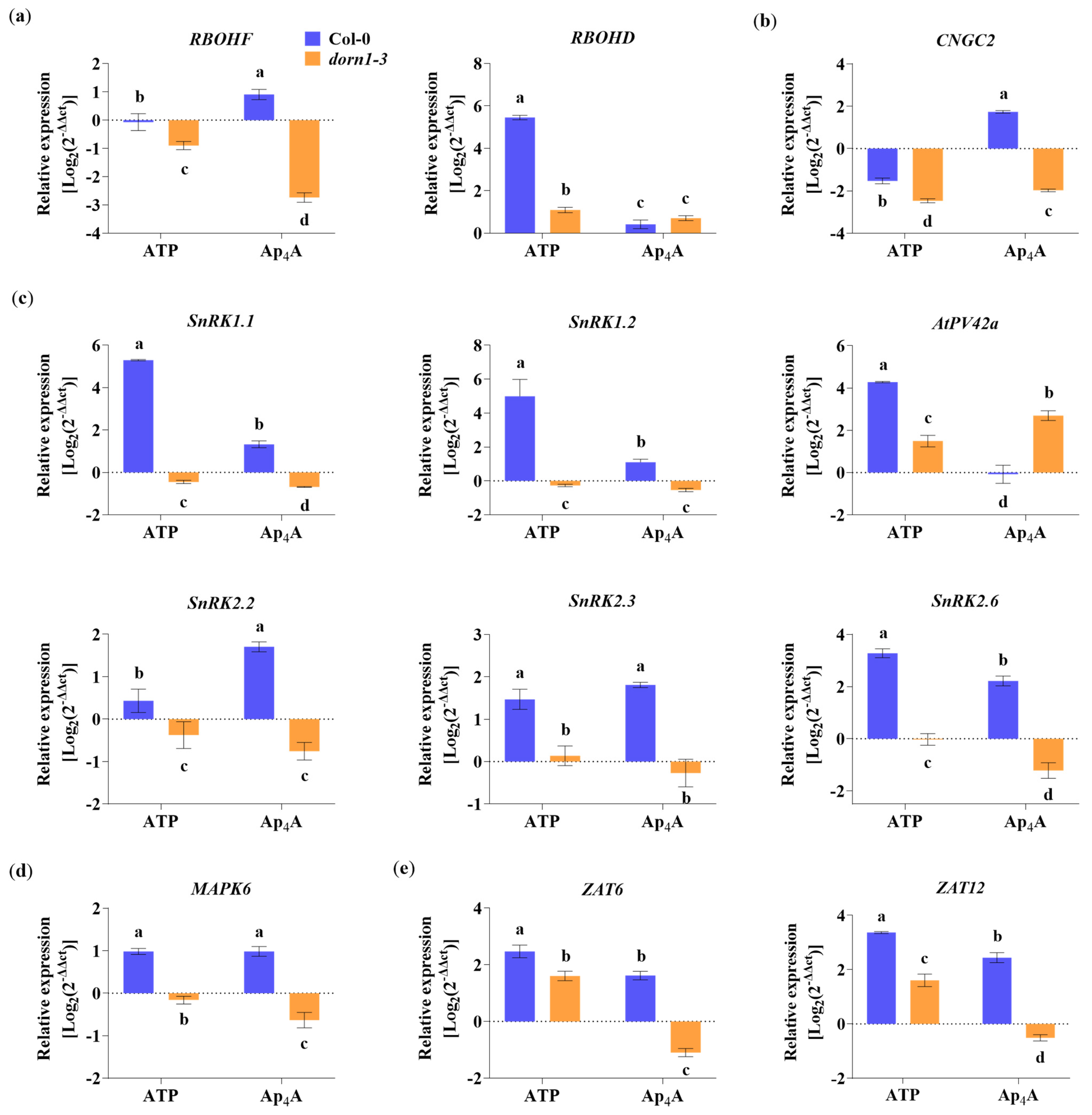
2.5. P2K1/DORN1 Is Implicated in Ap4A- and eATP-Responsive Gene Expression
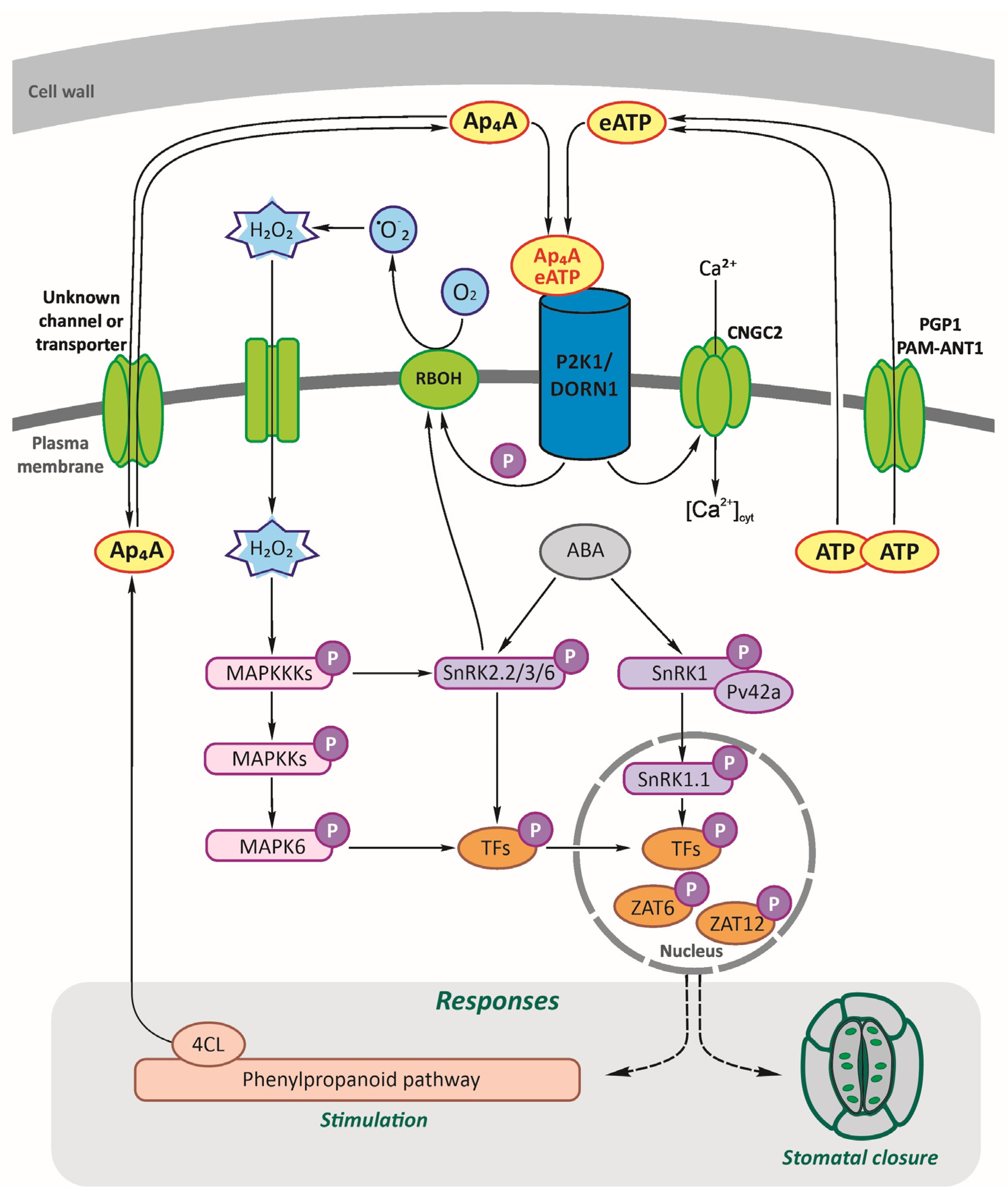
3. Discussion
4. Materials and Methods
4.1. Nucleotides
4.2. Plant Material
4.3. Stomatal Aperture Measurement
4.4. Detection of Intracellular ROS Burst in Leaves
4.5. Gene Expression Analyses
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, T.; Zhang, Y. Short- and Long-distance Signaling in Plant Defense. Plant J. 2021, 105, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Pietrowska-Borek, M.; Dobrogojski, J.; Sobieszczuk-Nowicka, E.; Borek, S. New Insight into Plant Signaling: Extracellular ATP and Uncommon Nucleotides. Cells 2020, 9, 345. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Sivaguru, M.; Stacey, G. Extracellular ATP in Plants. Visualization, Localization, and Analysis of Physiological Significance in Growth and Signaling. Plant Physiol. 2006, 142, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Steinebrunner, I.; Sun, Y.; Butterfield, T.; Torres, J.; Arnold, D.; Gonzalez, A.; Jacob, F.; Reichler, S.; Roux, S.J. Apyrases (Nucleoside Triphosphate-Diphosphohydrolases) Play a Key Role in Growth Control in Arabidopsis. Plant Physiol. 2007, 144, 961–975. [Google Scholar] [CrossRef]
- Riewe, D.; Grosman, L.; Fernie, A.R.; Wucke, C.; Geigenberger, P. The Potato-Specific Apyrase Is Apoplastically Localized and Has Influence on Gene Expression, Growth, and Development. Plant Physiol. 2008, 147, 1092–1109. [Google Scholar] [CrossRef]
- Tonón, C.; Cecilia Terrile, M.; José Iglesias, M.; Lamattina, L.; Casalongué, C. Extracellular ATP, Nitric Oxide and Superoxide Act Coordinately to Regulate Hypocotyl Growth in Etiolated Arabidopsis Seedlings. J. Plant Physiol. 2010, 167, 540–546. [Google Scholar] [CrossRef]
- Clark, G.; Wu, M.; Wat, N.; Onyirimba, J.; Pham, T.; Herz, N.; Ogoti, J.; Gomez, D.; Canales, A.A.; Aranda, G.; et al. Both the Stimulation and Inhibition of Root Hair Growth Induced by Extracellular Nucleotides in Arabidopsis Are Mediated by Nitric Oxide and Reactive Oxygen Species. Plant Mol. Biol. 2010, 74, 423–435. [Google Scholar] [CrossRef]
- Zhu, R.; Dong, X.; Xue, Y.; Xu, J.; Zhang, A.; Feng, M.; Zhao, Q.; Xia, S.; Yin, Y.; He, S.; et al. Redox-Responsive Transcription Factor 1 (RRFT1) Is Involved in Extracellular ATP-Regulated Arabidopsis thaliana Seedling Growth. Plant Cell Physiol. 2020, 61, 685–698. [Google Scholar] [CrossRef]
- Reichler, S.A.; Torres, J.; Rivera, A.L.; Cintolesi, V.A.; Clark, G.; Roux, S.J. Intersection of Two Signalling Pathways: Extracellular Nucleotides Regulate Pollen Germination and Pollen Tube Growth via Nitric Oxide. J. Exp. Bot. 2009, 60, 2129–2138. [Google Scholar] [CrossRef]
- Wu, Y.; Qin, B.; Feng, K.; Yan, R.; Kang, E.; Liu, T.; Shang, Z. Extracellular ATP Promoted Pollen Germination and Tube Growth of Nicotiana Tabacum through Promoting K+ and Ca2+ Absorption. Plant Reprod. 2018, 31, 399–410. [Google Scholar] [CrossRef]
- Chivasa, S.; Ndimba, B.K.; Simon, W.J.; Lindsey, K.; Slabas, A.R. Extracellular ATP Functions as an Endogenous External Metabolite Regulating Plant Cell Viability. Plant Cell 2005, 17, 3019–3034. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Cao, Y.; Li, H.; Kim, D.; Ahsan, N.; Thelen, J.; Stacey, G. Extracellular ATP Elicits DORN1-Mediated RBOHD Phosphorylation to Regulate Stomatal Aperture. Nat. Commun. 2017, 8, 2265. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.; Zhang, T.; Koo, A.J.; Stacey, G.; Tanaka, K. Extracellular ATP Acts on Jasmonate Signaling to Reinforce Plant Defense. Plant Physiol. 2018, 176, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Goodman, H.L.; Kroon, J.T.M.; Tomé, D.F.A.; Hamilton, J.M.U.; Alqarni, A.O.; Chivasa, S. Extracellular ATP Targets Arabidopsis RIBONUCLEASE 1 to Suppress Mycotoxin Stress-Induced Cell Death. New Phytol. 2022, 235, 1531–1542. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Rajagopal, A.; Windsor, B.; Dudler, R.; Lloyd, A.; Roux, S.J. A Role for Ectophosphatase in Xenobiotic Resistance. Plant Cell 2000, 12, 519–533. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Yang, S.H.; Kim, T.J.; Han, J.S.; Suh, J.W. Hypertonic Stress Increased Extracellular ATP Levels and the Expression of Stress-Responsive Genes in Arabidopsis thaliana Seedlings. Biosci. Biotechnol. Biochem. 2009, 73, 1252–1256. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, X.; Deng, S.; Zhang, C.; Wang, M.; Ding, M.; Zhao, R.; Shen, X.; Zhou, X.; Lu, C.; et al. Extracellular ATP Signaling Is Mediated by H2O2 and Cytosolic Ca2+ in the Salt Response of Populus euphratica Cells. PLoS ONE 2012, 7, e53136. [Google Scholar] [CrossRef]
- Hou, Q.Z.; Sun, K.; Zhang, H.; Su, X.; Fan, B.Q.; Feng, H.Q. The Responses of Photosystem II and Intracellular ATP Production of Arabidopsis Leaves to Salt Stress Are Affected by Extracellular ATP. J. Plant Res. 2018, 131, 331–339. [Google Scholar] [CrossRef]
- Duong, H.N.; Cho, S.H.; Wang, L.; Pham, A.Q.; Davies, J.M.; Stacey, G. Cyclic Nucleotide-Gated Ion Channel 6 Is Involved in Extracellular ATP Signaling and Plant Immunity. Plant J. 2021, 109, 1386–1396. [Google Scholar] [CrossRef]
- Wang, L.; Ning, Y.; Sun, J.; Wilkins, K.A.; Matthus, E.; McNelly, R.E.; Dark, A.; Rubio, L.; Moeder, W.; Yoshioka, K.; et al. Arabidopsis thaliana CYCLIC NUCLEOTIDE-GATED CHANNEL2 Mediates Extracellular ATP Signal Transduction in Root Epidermis. New Phytol. 2022, 234, 412–421. [Google Scholar] [CrossRef]
- Foresi, N.P.; Laxalt, A.M.; Tonón, C.V.; Casalongué, C.A.; Lamattina, L. Extracellular ATP Induces Nitric Oxide Production in Tomato Cell Suspensions. Plant Physiol. 2007, 145, 589–592. [Google Scholar] [CrossRef]
- Wu, S.-J.; Wu, J.-Y. Extracellular ATP-Induced NO Production and Its Dependence on Membrane Ca2+ Flux in Salvia miltiorrhiza Hairy Roots. J. Exp. Bot. 2008, 59, 4007–4016. [Google Scholar] [CrossRef]
- Song, C.J.; Steinebrunner, I.; Wang, X.; Stout, S.C.; Roux, S.J. Extracellular ATP Induces the Accumulation of Superoxide via NADPH Oxidases in Arabidopsis. Plant Physiol. 2006, 140, 1222–1232. [Google Scholar] [CrossRef]
- Wu, S.-J.; Liu, Y.-S.; Wu, J.-Y. The Signaling Role of Extracellular ATP and Its Dependence on Ca2+ Flux in Elicitation of Salvia miltiorrhiza Hairy Root Cultures. Plant Cell Physiol. 2008, 49, 617–624. [Google Scholar] [CrossRef]
- Demidchik, V.; Shang, Z.; Shin, R.; Thompson, E.; Rubio, L.; Laohavisit, A.; Mortimer, J.C.; Chivasa, S.; Slabas, A.R.; Glover, B.J.; et al. Plant Extracellular ATP Signalling by Plasma Membrane NADPH Oxidase and Ca2+ Channels. Plant J. 2009, 58, 903–913. [Google Scholar] [CrossRef]
- Choi, J.; Tanaka, K.; Cao, Y.; Qi, Y.; Qiu, J.; Liang, Y.; Lee, S.Y.; Stacey, G. Identification of a Plant Receptor for Extracellular ATP. Science 2014, 343, 290–294. [Google Scholar] [CrossRef]
- Li, P.; Zhao, L.; Qi, F.; Htwe, N.M.P.S.; Li, Q.; Zhang, D.; Lin, F.; Shang-Guan, K.; Liang, Y. The Receptor-like Cytoplasmic Kinase RIPK Regulates Broad-Spectrum ROS Signaling in Multiple Layers of Plant Immune System. Mol. Plant 2021, 14, 1652–1667. [Google Scholar] [CrossRef]
- Pietrowska-Borek, M.; Nuc, K.; Zielezińska, M.; Guranowski, A. Diadenosine Polyphosphates (Ap3A and Ap4A) Behave as Alarmones Triggering the Synthesis of Enzymes of the Phenylpropanoid Pathway in Arabidopsis thaliana. FEBS Open Bio 2011, 1, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pietrowska-Borek, M.; Czekała, Ł.; Belchí-Navarro, S.; Pedreño, M.A.; Guranowski, A. Diadenosine Triphosphate Is a Novel Factor Which in Combination with Cyclodextrins Synergistically Enhances the Biosynthesis of Trans-Resveratrol in Vitis vinifera Cv. Monastrell Suspension Cultured Cells. Plant Physiol. Biochem. 2014, 84, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Pietrowska-Borek, M.; Wojdyła-Mamoń, A.; Dobrogojski, J.; Młynarska-Cieślak, A.; Baranowski, M.R.; Dąbrowski, J.M.; Kowalska, J.; Jemielity, J.; Borek, S.; Pedreño, M.A.; et al. Purine and Pyrimidine Dinucleoside Polyphosphates Differentially Affect the Phenylpropanoid Pathway in Vitis vinifera L. Cv. Monastrell Suspension Cultured Cells. Plant Physiol. Biochem. 2020, 147, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Paiva, N.L. Stress-Induced Phenylpropanoid Metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, F.; McLennan, A.G.; Urbaniak, M.D.; Jones, N.J.; Copeland, N.A. Re-Evaluation of Diadenosine Tetraphosphate (Ap4A) from a Stress Metabolite to Bona Fide Secondary Messenger. Front. Mol. Biosci. 2020, 7, 606807. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.C.; Bochner, B.R.; Ames, B.N. AppppA, Heat-Shock Stress, and Cell Oxidation. Proc. Natl. Acad. Sci. USA 1983, 80, 7496–7500. [Google Scholar] [CrossRef]
- Bochner, B.R.; Lee, P.C.; Wilson, S.W.; Cutler, C.W.; Ames, B.N. AppppA and Related Adenylylated Nucleotides Are Synthesized as a Consequence of Oxidation Stress. Cell 1984, 37, 225–232. [Google Scholar] [CrossRef]
- Baltzinger, M.; Ebel, J.-P.; Remy, P. Accumulation of Dinucleoside Polyphosphates in Saccharomyces cerevisiae under Stress Conditions. High Levels Are Associated with Cell Death. Biochimie 1986, 68, 1231–1236. [Google Scholar] [CrossRef]
- Coste, H.; Brevet, A.; Plateau, P.; Blanquet, S. Non-Adenylylated Bis(5’-Nucleosidyl) Tetraphosphates Occur in Saccharomyces cerevisiae and in Escherichia coli and Accumulate upon Temperature Shift or Exposure to Cadmium. J. Biol. Chem. 1987, 262, 12096–12103. [Google Scholar] [CrossRef]
- Pálfi, Z.; Surányi, G.; Borbély, G. Alterations in the Accumulation of Adenylylated Nucleotides in Heavy-Metal-Ion-Stressed and Heat-Stressed Synechococcus Sp. Strain PCC 6301, a Cyanobacterium, in Light and Dark. Biochem. J. 1991, 276, 487–491. [Google Scholar] [CrossRef][Green Version]
- Pham, A.Q.; Cho, S.-H.; Nguyen, C.T.; Stacey, G. Arabidopsis Lectin Receptor Kinase P2K2 Is a Second Plant Receptor for Extracellular ATP and Contributes to Innate Immunity. Plant Physiol. 2020, 183, 1364–1375. [Google Scholar] [CrossRef]
- Cho, S.-H.; Nguyen, C.T.; Pham, A.Q.; Stacey, G. Computational Prediction and in Vitro Analysis of the Potential Ligand Binding Site within the Extracellular ATP Receptor, P2K2. Plant Signal. Behav. 2023, 18, e2173146. [Google Scholar] [CrossRef]
- Jose, J.; Ghantasala, S.; Choudhury, S.R. Arabidopsis Transmembrane Receptor-like Kinases (RLKS): A Bridge between Extracellular Signal and Intracellular Regulatory Machinery. Int. J. Mol. Sci. 2020, 21, 4000. [Google Scholar] [CrossRef] [PubMed]
- Vigne, P.; Breittmayer, J.P.; Frelin, C. Diadenosine Polyphosphates as Antagonists of the Endogenous P2Y1 Receptor in Rat Brain Capillary Endothelial Cells of the B7 and B10 Clones: APnAs as Antagonists of P2Y1 Receptors. Br. J. Pharmacol. 2000, 129, 1506–1512. [Google Scholar] [CrossRef] [PubMed][Green Version]
- McDonald, H.A.; Chu, K.L.; Bianchi, B.R.; McKenna, D.G.; Briggs, C.A.; Burgard, E.C.; Lynch, K.J.; Faltynek, C.; Cartmell, J.; Jarvis, M.F. Potent Desensitization of Human P2X3 Receptors by Diadenosine Polyphosphates. Eur. J. Pharmacol. 2002, 435, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chang, C.-F.; Morales, M.; Chiang, Y.-H.; Harvey, B.K.; Su, T.-P.; Tsao, L.-I.; Chen, S.; Thiemermann, C. Diadenosine Tetraphosphate Protects against Injuries Induced by Ischemia and 6-Hydroxydopamine in Rat Brain. J. Neurosci. 2003, 23, 7958–7965. [Google Scholar] [CrossRef] [PubMed]
- Verspohl, E.J.; Johannwille, B.; Kaiserling-Buddemeier, I.; Schlüter, H.; Hagemann, J. Diadenosine Polyphosphates in Cultured Vascular Smooth-Muscle Cells and Endothelium Cells—Their Interaction with Specific Receptors and Their Degradation. J. Pharm. Pharmacol. 2010, 51, 1175–1181. [Google Scholar] [CrossRef]
- Burnstock, G. Purine and Purinergic Receptors. Brain Neurosci. Adv. 2018, 2, 1–10. [Google Scholar] [CrossRef]
- Hsu, P.-K.; Dubeaux, G.; Takahashi, Y.; Schroeder, J.I. Signaling Mechanisms in Abscisic Acid-Mediated Stomatal Closure. Plant J. Cell Mol. Biol. 2021, 105, 307–321. [Google Scholar] [CrossRef]
- Danquah, A.; De Zelicourt, A.; Colcombet, J.; Hirt, H. The Role of ABA and MAPK Signaling Pathways in Plant Abiotic Stress Responses. Biotechnol. Adv. 2014, 32, 40–52. [Google Scholar] [CrossRef]
- Bharath, P.; Gahir, S.; Raghavendra, A.S. Abscisic Acid-Induced Stomatal Closure: An Important Component of Plant Defense against Abiotic and Biotic Stress. Front. Plant Sci. 2021, 12, 615114. [Google Scholar] [CrossRef]
- Guranowski, A. Metabolism of Diadenosine Tetraphosphate (Ap4A) and Related Nucleotides in Plants; Review with Historical and General Perspective. Front. Biosci. 2004, 9, 1398. [Google Scholar] [CrossRef]
- Waszczak, C.; Carmody, M.; Kangasjarvi, J. Reactive Oxygen Species in Plant Signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef]
- Smirnoff, N.; Arnaud, D. Hydrogen Peroxide Metabolism and Functions in Plants. New Phytol. 2019, 221, 1197–1214. [Google Scholar] [CrossRef] [PubMed]
- Macho, A.P.; Zipfel, C. Plant PRRs and the Activation of Innate Immune Signaling. Mol. Cell 2014, 54, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive Oxygen Species Signalling in Plant Stress Responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Khokon, A.R.; Okuma, E.; Hossain, M.A.; Munemasa, S.; Uraji, M.; Nakamura, Y.; Mori, I.C.; Murata, Y. Involvement of Extracellular Oxidative Burst in Salicylic Acid-Induced Stomatal Closure in Arabidopsis: Extracellular ROS Mediate SA-Induced Stomatal Closure. Plant Cell Environ. 2011, 34, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Jewell, J.B.; Sowders, J.M.; He, R.; Willis, M.A.; Gang, D.R.; Tanaka, K. Extracellular ATP Shapes a Defense-Related Transcriptome Both Independently and along with Other Defense Signaling Pathways. Plant Physiol. 2019, 179, 1144–1158. [Google Scholar] [CrossRef] [PubMed]
- Duszyn, M.; Świeżawska, B.; Szmidt-Jaworska, A.; Jaworski, K. Cyclic Nucleotide Gated Channels (CNGCs) in Plant Signalling-Current Knowledge and Perspectives. J. Plant Physiol. 2019, 241, 153035. [Google Scholar] [CrossRef]
- Jarratt-Barnham, E.; Wang, L.; Ning, Y.; Davies, J.M. The Complex Story of Plant Cyclic Nucleotide-Gated Channels. Int. J. Mol. Sci. 2021, 22, 874. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Y.; Zhu, J.-K. Thriving under Stress: How Plants Balance Growth and the Stress Response. Dev. Cell 2020, 55, 529–543. [Google Scholar] [CrossRef]
- Baudry, K.; Barbut, F.; Domenichini, S.; Guillaumot, D.; Thy, M.P.; Vanacker, H.; Majeran, W.; Krieger-Liszkay, A.; Issakidis-Bourguet, E.; Lurin, C. Adenylates Regulate Arabidopsis Plastidial Thioredoxin Activities through the Binding of a CBS Domain Protein. Plant Physiol. 2022, 189, 2298–2314. [Google Scholar] [CrossRef]
- Matsushita, M.; Nakamura, T.; Moriizumi, H.; Miki, H.; Takekawa, M. Stress-Responsive MTK1 SAPKKK Serves as a Redox Sensor That Mediates Delayed and Sustained Activation of SAPKs by Oxidative Stress. Sci. Adv. 2020, 6, eaay9778. [Google Scholar] [CrossRef] [PubMed]
- Byrne, D.P.; Shrestha, S.; Galler, M.; Cao, M.; Daly, L.A.; Campbell, A.E.; Eyers, C.E.; Veal, E.A.; Kannan, N.; Eyers, P.A. Aurora A Regulation by Reversible Cysteine Oxidation Reveals Evolutionarily Conserved Redox Control of Ser/Thr Protein Kinase Activity. Sci. Signal. 2020, 13, eaax2713. [Google Scholar] [CrossRef] [PubMed]
- Smékalová, V.; Doskočilová, A.; Komis, G.; Šamaj, J. Crosstalk between Secondary Messengers, Hormones and MAPK Modules during Abiotic Stress Signalling in Plants. Biotechnol. Adv. 2014, 32, 2–11. [Google Scholar] [CrossRef] [PubMed]
- McLennan, A. Dinucleoside Polyphosphates—Friend or Foe? Pharmacol. Ther. 2000, 87, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Pietrowska-Borek, M.; Stuible, H.-P.; Kombrink, E.; Guranowski, A. 4-Coumarate:Coenzyme A Ligase Has the Catalytic Capacity to Synthesize and Reuse Various (Di)Adenosine Polyphosphates. Plant Physiol. 2003, 131, 1401–1410. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.-G.; Hilleary, R.; Swanson, S.J.; Kim, S.-H.; Gilroy, S. Rapid, Long-Distance Electrical and Calcium Signaling in Plants. Annu. Rev. Plant Biol. 2016, 67, 287–307. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, S. MAPK Cascades in Plant Disease Resistance Signaling. Annu. Rev. Phytopathol. 2013, 51, 245–266. [Google Scholar] [CrossRef]
- Medina-Castellanos, E.; Esquivel-Naranjo, E.U.; Heil, M.; Herrera-Estrella, A. Extracellular ATP Activates MAPK and ROS Signaling during Injury Response in the Fungus Trichoderma atroviride. Front. Plant Sci. 2014, 5, 659. [Google Scholar] [CrossRef]
- Chen, D.; Hao, F.; Mu, H.; Ahsan, N.; Thelen, J.J.; Stacey, G. S-Acylation of P2K1 Mediates Extracellular ATP-Induced Immune Signaling in Arabidopsis. Nat. Commun. 2021, 12, 2750. [Google Scholar] [CrossRef]
- Cho, S.-H.; Tóth, K.; Kim, D.; Vo, P.H.; Lin, C.-H.; Handakumbura, P.P.; Ubach, A.R.; Evans, S.; Paša-Tolić, L.; Stacey, G. Activation of the Plant Mevalonate Pathway by Extracellular ATP. Nat. Commun. 2022, 13, 450. [Google Scholar] [CrossRef]
- Zou, M.; Guo, M.; Zhou, Z.; Wang, B.; Pan, Q.; Li, J.; Zhou, J.-M.; Li, J. MPK3- and MPK6-Mediated VLN3 Phosphorylation Regulates Actin Dynamics during Stomatal Immunity in Arabidopsis. Nat. Commun. 2021, 12, 6474. [Google Scholar] [CrossRef] [PubMed]
- Jossier, M.; Bouly, J.-P.; Meimoun, P.; Arjmand, A.; Lessard, P.; Hawley, S.; Grahame Hardie, D.; Thomas, M. SnRK1 (SNF1-Related Kinase 1) Has a Central Role in Sugar and ABA Signalling in Arabidopsis thaliana. Plant J. 2009, 59, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Li, T.; Zhao, Y.; Chang, Y.; Wu, L.; Chen, G.; Day, B.; Jiang, K. Calcium-Dependent ABA Signaling Functions in Stomatal Immunity by Regulating Rapid SA Responses in Guard Cells. J. Plant Physiol. 2022, 268, 153585. [Google Scholar] [CrossRef]
- Signorelli, S.; Tarkowski, Ł.P.; Van den Ende, W.; Bassham, D.C. Linking Autophagy to Abiotic and Biotic Stress Responses. Trends Plant Sci. 2019, 24, 413–430. [Google Scholar] [CrossRef]
- Belda-Palazón, B.; Adamo, M.; Valerio, C.; Ferreira, L.J.; Confraria, A.; Reis-Barata, D.; Rodrigues, A.; Meyer, C.; Rodriguez, P.L.; Baena-González, E. A Dual Function of SnRK2 Kinases in the Regulation of SnRK1 and Plant Growth. Nat. Plants 2020, 6, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Fang, X.; Xiao, C.; Ma, Z.; Huang, X.; Su, J.; Li, J.; Wang, J.; Wang, S.; Luan, S.; et al. Kinase SnRK1.1 Regulates Nitrate Channel SLAH3 Engaged in Nitrate-Dependent Alleviation of Ammonium Toxicity. Plant Physiol. 2021, 186, 731–749. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Micallef, B.J.; Tetlow, I.J.; Mullen, R.T.; Feil, R.; Lunn, J.E.; Emes, M.J. AKINβ1, a Subunit of SnRK1, Regulates Organic Acid Metabolism and Acts as a Global Modulator of Genes Involved in Carbon, Lipid, and Nitrogen Metabolism. J. Exp. Bot. 2020, 71, 1010–1028. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, Q.; Tian, C.; Liu, G.; Pan, Y.; Xu, X.; Shi, X.; Zhang, Z.; Meng, L. Physiological and Transcriptome Analyses of CaCl2 Treatment to Alleviate Chilling Injury in Pineapple. Plants 2022, 11, 2215. [Google Scholar] [CrossRef]
- Son, S.; Im, J.H.; Ko, J.-H.; Han, K.-H. SNF1-Related Protein Kinase 1 Represses Arabidopsis Growth through Post-Translational Modification of E2Fa in Response to Energy Stress. New Phytol. 2023, 237, 823–839. [Google Scholar] [CrossRef]
- Shi, H.; Liu, G.; Wei, Y.; Chan, Z. The Zinc-Finger Transcription Factor ZAT6 Is Essential for Hydrogen Peroxide Induction of Anthocyanin Synthesis in Arabidopsis. Plant Mol. Biol. 2018, 97, 165–176. [Google Scholar] [CrossRef]
- Opdenakker, K.; Remans, T.; Keunen, E.; Vangronsveld, J.; Cuypers, A. Exposure of Arabidopsis thaliana to Cd or Cu Excess Leads to Oxidative Stress Mediated Alterations in MAPKinase Transcript Levels. Environ. Exp. Bot. 2012, 83, 53–61. [Google Scholar] [CrossRef]
- Shi, H.; Wang, X.; Ye, T.; Chen, F.; Deng, J.; Yang, P.; Zhang, Y.; Chan, Z. The Cysteine2/Histidine2-Type Transcription Factor ZINC FINGER OF ARABIDOPSIS THALIANA6 Modulates Biotic and Abiotic Stress Responses by Activating Salicylic Acid-Related Genes and C-REPEAT-BINDING FACTOR Genes in Arabidopsis. Plant Physiol. 2014, 165, 1367–1379. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, L.; Yan, X.; Liu, Y.; Wang, R.; Fan, T.; Ren, Y.; Tang, X.; Xiao, F.; Liu, Y.; et al. Zinc-Finger Transcription Factor ZAT6 Positively Regulates Cadmium Tolerance through the Glutathione-Dependent Pathway in Arabidopsis. Plant Physiol. 2016, 171, 707–719. [Google Scholar] [CrossRef] [PubMed]
- Dang, F.; Li, Y.; Wang, Y.; Lin, J.; Du, S.; Liao, X. ZAT10 Plays Dual Roles in Cadmium Uptake and Detoxification in Arabidopsis. Front. Plant Sci. 2022, 13, 994100. [Google Scholar] [CrossRef]
- Brumbarova, T.; Le, C.T.T.; Ivanov, R.; Bauer, P. Regulation of ZAT12 Protein Stability: The Role of Hydrogen Peroxide. Plant Signal. Behav. 2016, 11, e1137408. [Google Scholar] [CrossRef][Green Version]
- Myers, R.J., Jr.; Fichman, Y.; Stacey, G.; Mittler, R. Extracellular ATP Plays an Important Role in Systemic Wound Response Activation. Plant Physiol. 2022, 189, 1314–1325. [Google Scholar] [CrossRef]
- Rieder, B.; Neuhaus, H.E. Identification of an Arabidopsis Plasma Membrane-Located ATP Transporter Important for Anther Development. Plant Cell 2011, 23, 1932–1944. [Google Scholar] [CrossRef]
- Müller, K.; Carstens, A.C.; Linkies, A.; Torres, M.A.; Leubner-Metzger, G. The NADPH-oxidase AtrbohB Plays a Role in Arabidopsis Seed After-ripening. New Phytol. 2009, 184, 885–897. [Google Scholar] [CrossRef]
- Daudi, A.; O’Brien, J.A. Detection of Hydrogen Peroxide by DAB Staining in Arabidopsis Leaves. Bio-Protocol 2012, 2, e263. [Google Scholar] [CrossRef]
- Pietrowska-Borek, M.; Nuc, K.; Guranowski, A. Exogenous Adenosine 5’-Phosphoramidate Behaves as a Signal Molecule in Plants; It Augments Metabolism of Phenylpropanoids and Salicylic Acid in Arabidopsis thaliana Seedlings. Plant Physiol. Biochem. 2015, 94, 144–152. [Google Scholar] [CrossRef]
- Pietrowska-Borek, M.; Nuc, K. Both Cyclic-AMP and Cyclic-GMP Can Act as Regulators of the Phenylpropanoid Pathway in Arabidopsis thaliana Seedlings. Plant Physiol. Biochem. 2013, 70, 142–149. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Doyle, J.L.; Doyle, J.M. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Nakashima, K.; Fujita, Y.; Kanamori, N.; Katagiri, T.; Umezawa, T.; Kidokoro, S.; Maruyama, K.; Yoshida, T.; Ishiyama, K.; Kobayashi, M.; et al. Three Arabidopsis SnRK2 Protein Kinases, SRK2D/SnRK2.2, SRK2E/SnRK2.6/OST1 and SRK2I/SnRK2.3, Involved in ABA Signaling Are Essential for the Control of Seed Development and Dormancy. Plant Cell Physiol. 2009, 50, 1345–1363. [Google Scholar] [CrossRef]
- Fang, L.; Hou, X.; Lee, L.Y.C.; Liu, L.; Yan, X.; Yu, H. AtPV42a and AtPV42b Redundantly Regulate Reproductive Development in Arabidopsis thaliana. PLoS ONE 2011, 6, e19033. [Google Scholar] [CrossRef] [PubMed]
- Kannan, P.; Pandey, D.; Gupta, A.K.; Punetha, H.; Taj, G.; Kumar, A. Expression Analysis of MAP2K9 and MAPK6 during Pathogenesis of Alternaria Blight in Arabidopsis thaliana Ecotype Columbia. Mol. Biol. Rep. 2012, 39, 4439–4444. [Google Scholar] [CrossRef] [PubMed]
- Le, C.T.T.; Brumbarova, T.; Ivanov, R.; Stoof, C.; Weber, E.; Mohrbacher, J.; Fink-Straube, C.; Bauer, P. ZINC FINGER OF ARABIDOPSIS THALIANA12 (ZAT12) Interacts with FER-LIKE IRON DEFICIENCY-INDUCED TRANSCRIPTION FACTOR (FIT) Linking Iron Deficiency and Oxidative Stress Responses. Plant Physiol. 2016, 170, 540–557. [Google Scholar] [CrossRef] [PubMed]
- Morales, J.; Kadota, Y.; Zipfel, C.; Molina, A.; Torres, M.-A. The Arabidopsis NADPH Oxidases RbohD and RbohF Display Differential Expression Patterns and Contributions during Plant Immunity. EXBOTJ 2016, 67, 1663–1676. [Google Scholar] [CrossRef] [PubMed]
- Guranowski, A.; Starzyńska, E.; Pietrowska-Borek, M.; Rejman, D.; Blackburn, G.M. Novel Diadenosine Polyphosphate Analogs with Oxymethylene Bridges Replacing Oxygen in the Polyphosphate Chain: Potential Substrates and/or Inhibitors of Ap4A Hydrolases. FEBS J. 2009, 276, 1546–1553. [Google Scholar] [CrossRef]
- Guranowski, A.; Wojdyła, A.M.; Pietrowska-Borek, M.; Bieganowski, P.; Khurs, E.N.; Cliff, M.J.; Blackburn, G.M.; Błaziak, D.; Stec, W.J. Fhit Proteins Can Also Recognize Substrates Other than Dinucleoside Polyphosphates. FEBS Lett. 2008, 582, 3152–3158. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dobrogojski, J.; Nguyen, V.H.; Kowalska, J.; Borek, S.; Pietrowska-Borek, M. The Plasma Membrane Purinoreceptor P2K1/DORN1 Is Essential in Stomatal Closure Evoked by Extracellular Diadenosine Tetraphosphate (Ap4A) in Arabidopsis thaliana. Int. J. Mol. Sci. 2023, 24, 16688. https://doi.org/10.3390/ijms242316688
Dobrogojski J, Nguyen VH, Kowalska J, Borek S, Pietrowska-Borek M. The Plasma Membrane Purinoreceptor P2K1/DORN1 Is Essential in Stomatal Closure Evoked by Extracellular Diadenosine Tetraphosphate (Ap4A) in Arabidopsis thaliana. International Journal of Molecular Sciences. 2023; 24(23):16688. https://doi.org/10.3390/ijms242316688
Chicago/Turabian StyleDobrogojski, Jędrzej, Van Hai Nguyen, Joanna Kowalska, Sławomir Borek, and Małgorzata Pietrowska-Borek. 2023. "The Plasma Membrane Purinoreceptor P2K1/DORN1 Is Essential in Stomatal Closure Evoked by Extracellular Diadenosine Tetraphosphate (Ap4A) in Arabidopsis thaliana" International Journal of Molecular Sciences 24, no. 23: 16688. https://doi.org/10.3390/ijms242316688
APA StyleDobrogojski, J., Nguyen, V. H., Kowalska, J., Borek, S., & Pietrowska-Borek, M. (2023). The Plasma Membrane Purinoreceptor P2K1/DORN1 Is Essential in Stomatal Closure Evoked by Extracellular Diadenosine Tetraphosphate (Ap4A) in Arabidopsis thaliana. International Journal of Molecular Sciences, 24(23), 16688. https://doi.org/10.3390/ijms242316688