
Thermostable Basic Fibroblast Growth Factor Enhances the Production and Activity of Human Wharton’s Jelly Mesenchymal Stem Cell-Derived Extracellular Vesicles

 , , , and
, , , and 
Abstract
:
1. Introduction
2. Results
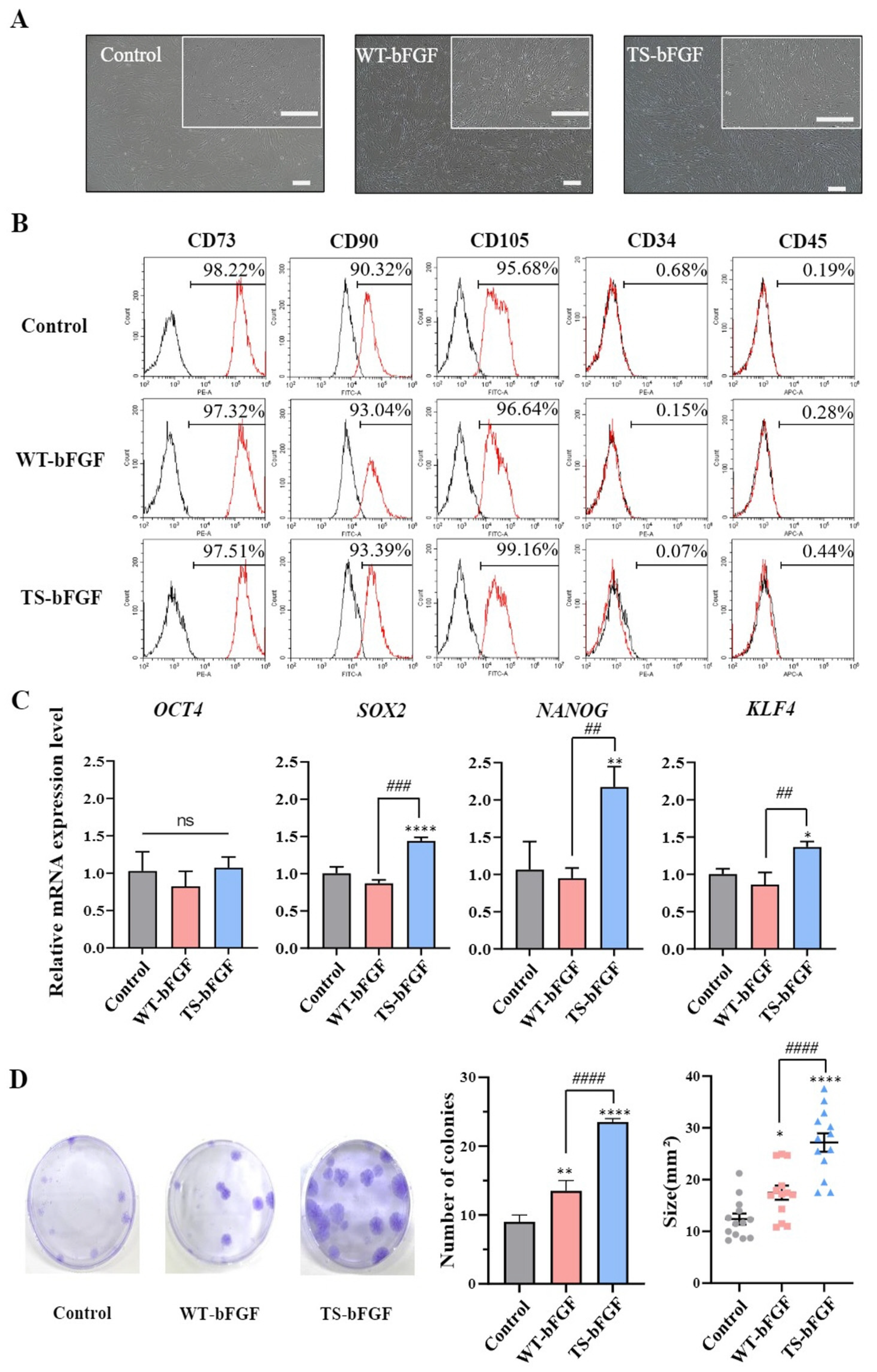
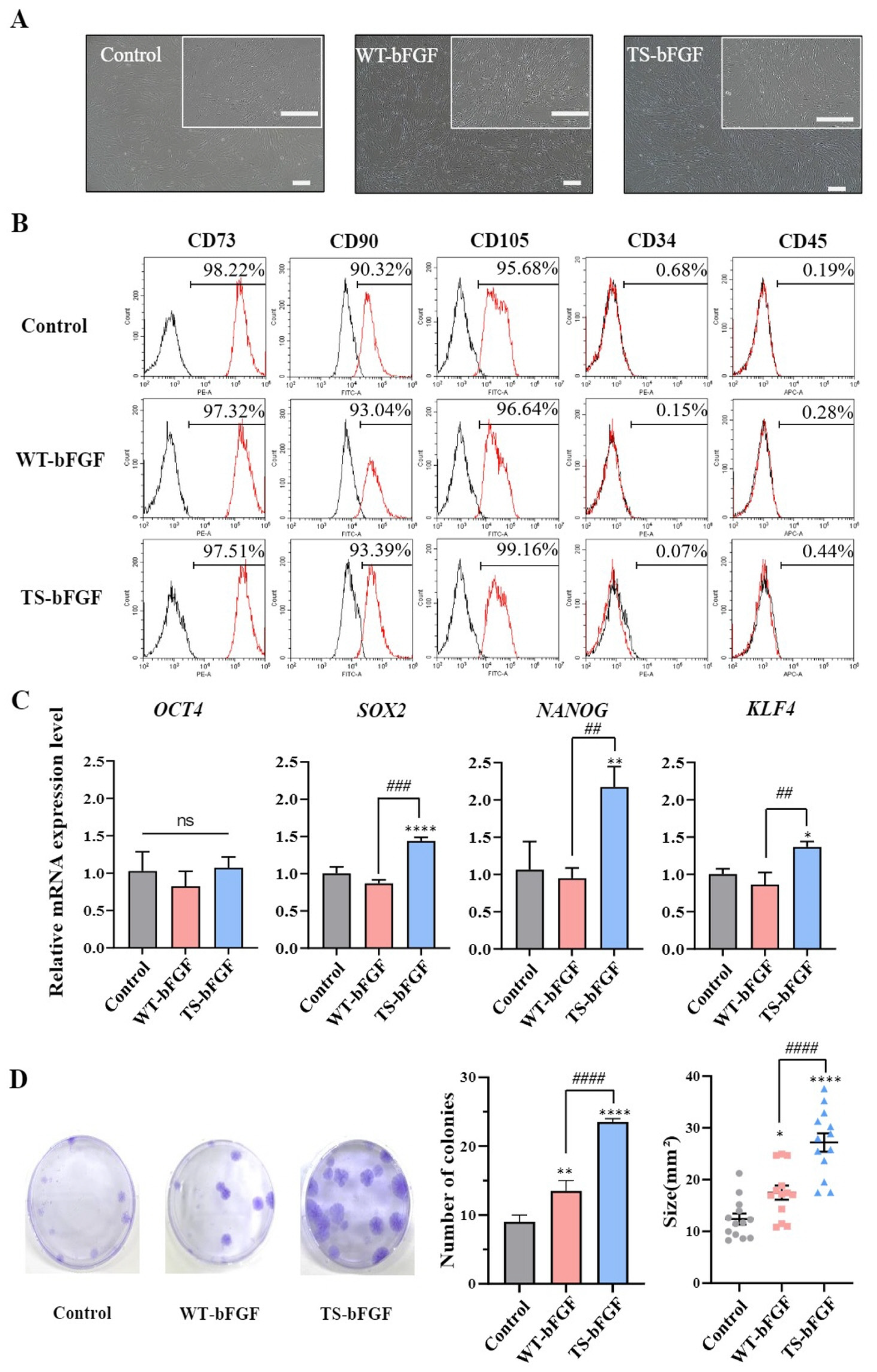
2.1. Characterization of WJ-MSCs Treated with Wild-Type bFGF or TS-bFGF
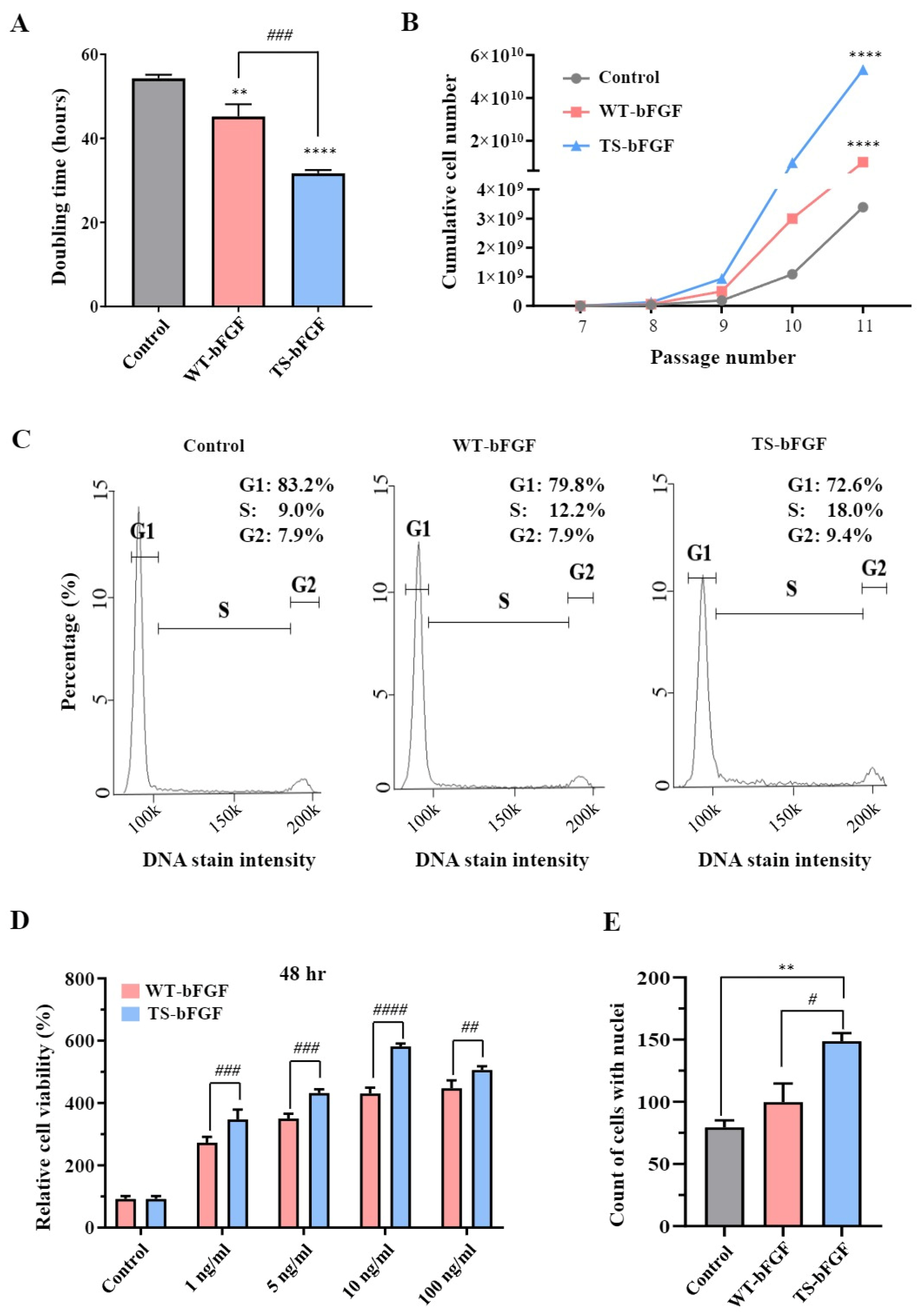
2.2. TS-bFGF Increases Cell Proliferation
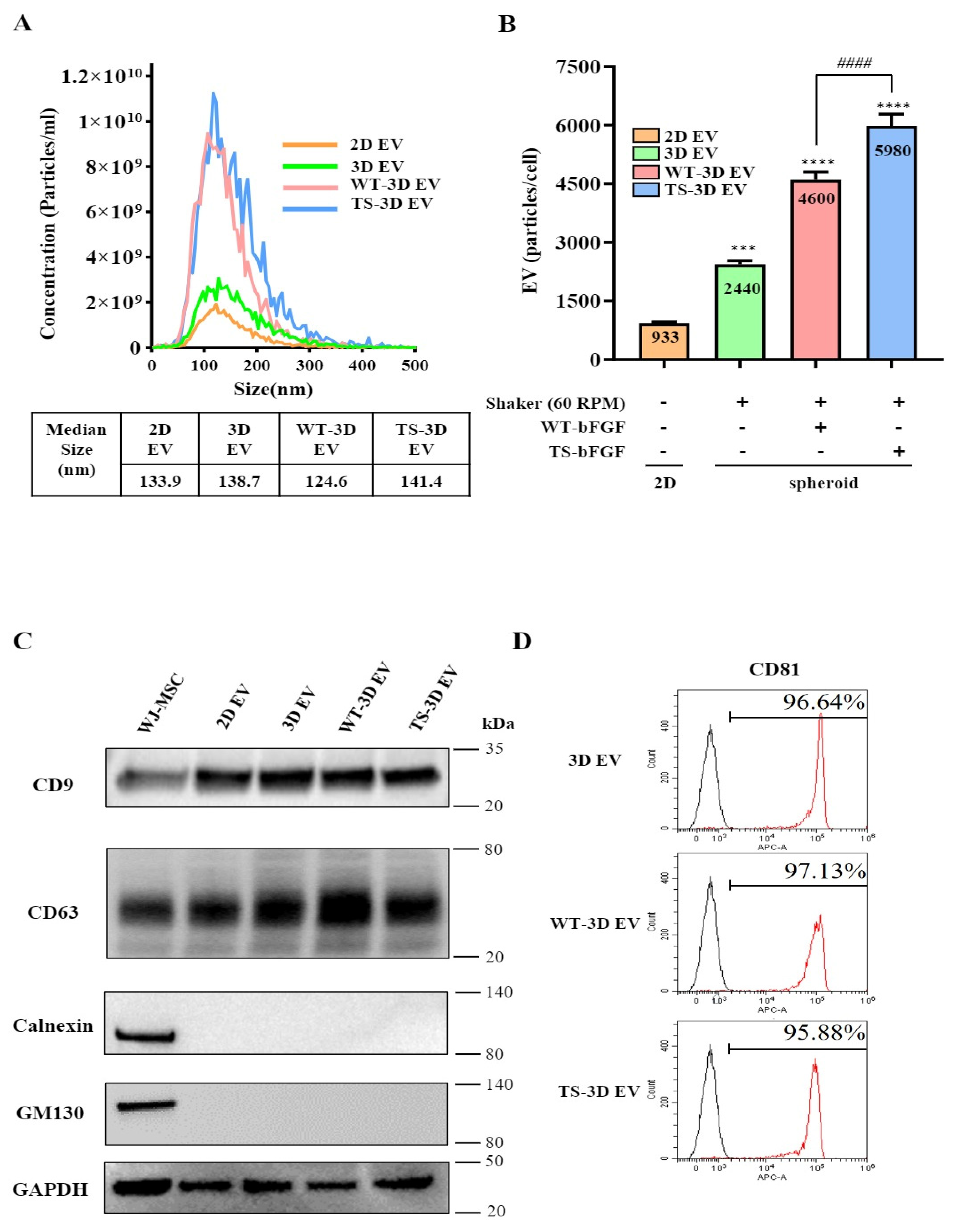
2.3. Characterization of WT-WJ-MSCs- and TS-WJ-MSCs-Derived Exosomes
2.4. TS-3D EVs Increased Cell Proliferation and Wound Closure Compared to WT-3D EVs
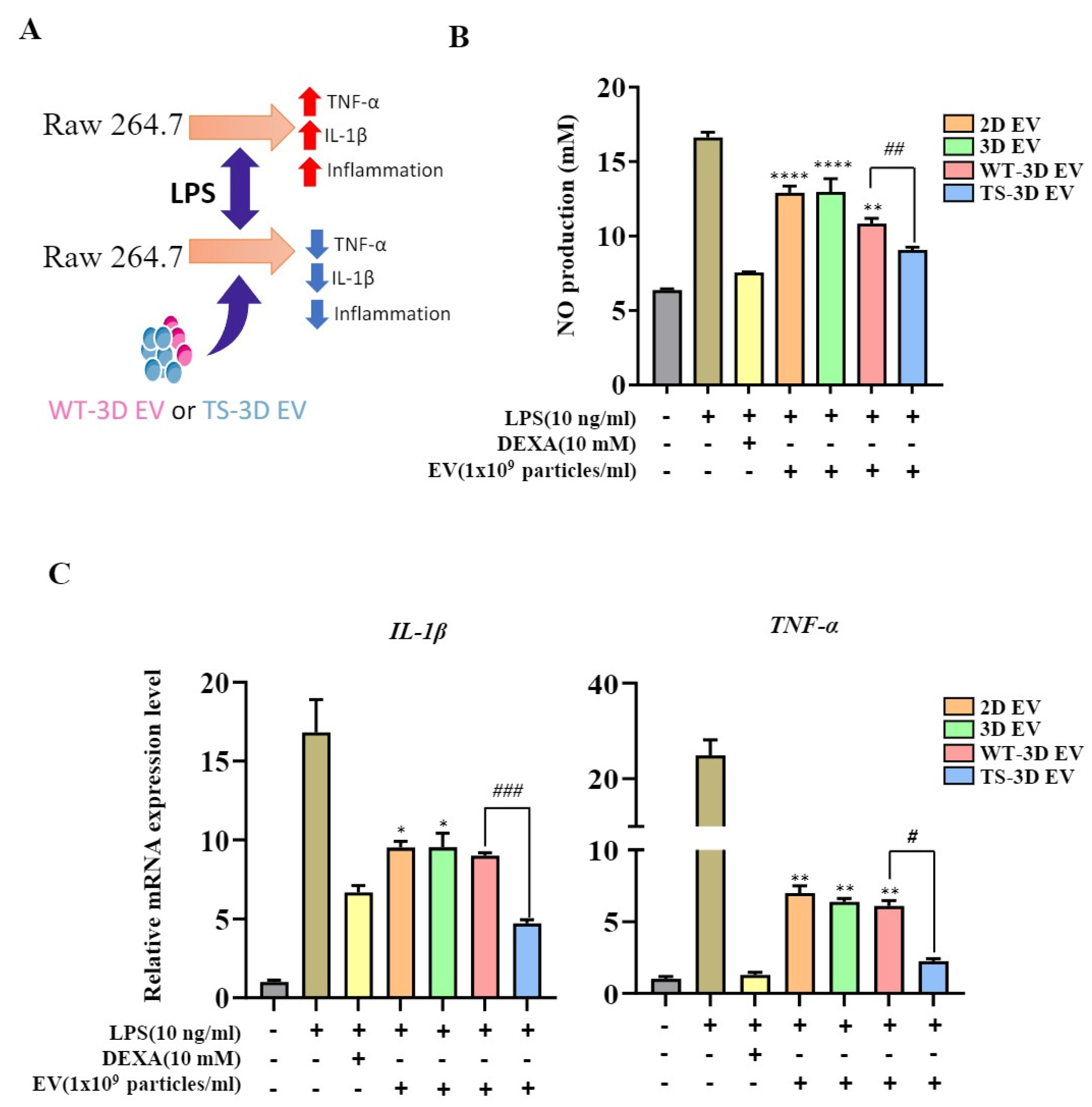
2.5. Anti-Inflammatory Activity of EVs from WT-WJ-MSCs and TS-WJ-MSCs
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Flow Cytometry
4.3. Quantitative RT-PCR
4.4. Fibroblast Colony-Forming Unit Assay
4.5. Cell Growth Kinetics
4.6. Cell Cycle Assay
4.7. Cell Proliferation Test
4.8. DAPI Staining Assay
4.9. Generation of EVs-Treated bFGF
4.10. EVs Produced with WT-bFGF or TS-bFGF
4.11. Western Blotting
4.12. In Vitro Migration Assay
4.13. NO Assay
4.14. Statistical Analyses
5. Patent
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Margiana, R.; Markov, A.; Zekiy, A.O.; Hamza, M.U.; Al-Dabbagh, K.A.; Al-Zubaidi, S.H.; Hameed, N.M.; Ahmad, I.; Sivaraman, R.; Kzar, H.H.; et al. Clinical application of mesenchymal stem cell in regenerative medicine: A narrative review. Stem Cell Res. Ther. 2022, 13, 366. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Garza, L.E.; Barrera-Barrera, S.A.; Barrera-Saldaña, H.A. Mesenchymal Stem Cell Therapies Approved by Regulatory Agencies around the World. Pharmaceuticals 2023, 16, 1334. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Lim, K.M.; Cho, J.-M.; Park, H.J.; Hwang, S.; Dayem, A.A.; Jeong, Y.J.; Shin, Y.; Hong, Y.; Song, K.; et al. Ductal delivery of extracellular vesicles promote the recovery from salivary gland inflammation. J. Control. Release 2023, 357, 235–248. [Google Scholar] [CrossRef]
- Han, Y.; Yang, J.; Fang, J.; Zhou, Y.; Candi, E.; Wang, J.; Hua, D.; Shao, C.; Shi, Y. The secretion profile of mesenchymal stem cells and potential applications in treating human diseases. Signal Transduct. Target. Ther. 2022, 7, 92. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Dayem, A.A.; Lee, S.; Choi, Y.; Lim, K.M.; Kim, S.; An, J.; Shin, Y.; Park, H.; Jeon, T.-I.; et al. Superior therapeutic activity of TGF-β-induced extracellular vesicles against interstitial cystitis. J. Control. Release 2022, 348, 924–937. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, Q.; Wang, Z.; Tong, H.; Ma, L.; Zhang, Y.; Shan, F.; Meng, Y.; Yuan, Z. Comparative analysis of human mesenchymal stem cells from fetal-bone marrow, adipose tissue, and Warton’s jelly as sources of cell immunomodulatory therapy. Hum. Vaccines Immunother. 2016, 12, 85–96. [Google Scholar] [CrossRef]
- Chetty, S.; Yarani, R.; Swaminathan, G.; Primavera, R.; Regmi, S.; Rai, S.; Zhong, J.; Ganguly, A.; Thakor, A.S. Umbilical cord mesenchymal stromal cells—From bench to bedside. Front. Cell Dev. Biol. 2022, 10, 1006295. [Google Scholar] [CrossRef]
- Liau, L.; Ruszymah, B.; Ng, M.; Law, J. Characteristics and clinical applications of Wharton’s jelly-derived mesenchymal stromal cells. Curr. Res. Transl. Med. 2020, 68, 5–16. [Google Scholar] [CrossRef]
- Subramanian, A.; Fong, C.-Y.; Biswas, A.; Bongso, A. Comparative characterization of cells from the various compartments of the human umbilical cord shows that the wharton’s jelly compartment provides the best source of clinically utilizable mesenchymal stem cells. PLoS ONE 2015, 10, e0127992. [Google Scholar] [CrossRef]
- Lim, K.M.; Dayem, A.A.; Choi, Y.; Lee, Y.; An, J.; Gil, M.; Lee, S.; Kwak, H.J.; Vellingirl, B.; Shin, H.J.; et al. High therapeutic and esthetic properties of extracellular vesicles produced from the stem cells and their spheroids cultured from ocular surgery-derived waste orbicularis oculi muscle tissues. Antioxidants 2021, 10, 1292. [Google Scholar] [CrossRef]
- Nourian Dehkordi, A.; Mirahmadi Babaheydari, F.; Chehelgerdi, M.; Raeisi Dehkordi, S. Skin tissue engineering: Wound healing based on stem-cell-based therapeutic strategies. Stem Cell Res. Ther. 2019, 10, 111. [Google Scholar] [CrossRef] [PubMed]
- Hazrati, A.; Malekpour, K.; Soudi, S.; Hashemi, S.M. Mesenchymal stromal/stem cells spheroid culture effect on the therapeutic efficacy of these cells and their exosomes: A new strategy to overcome cell therapy limitations. Biomed. Pharmacother. 2022, 152, 113211. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.M.; Han, J.; Lee, Y.; Park, J.; Dayem, A.A.; Myung, S.; An, J.; Song, K.; Kang, G.; Kim, S.; et al. Rapid production method with increased yield of high-purity extracellular vesicles obtained using extended mitochondrial targeting domain peptide. J. Extracell. Vesicles 2022, 11, e12274. [Google Scholar] [CrossRef] [PubMed]
- Lötvall, J.; Hill, A.F.; Hochberg, F.; Buzás, E.I.; Di Vizio, D.; Gardiner, C.; Gho, Y.S.; Kurochkin, I.V.; Mathivanan, S.; Quesenberry, P.; et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: A position statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2014, 3, 26913. [Google Scholar] [CrossRef]
- Kou, M.; Huang, L.; Yang, J.; Chiang, Z.; Chen, S.; Liu, J.; Guo, L.; Zhang, X.; Zhou, X.; Xu, X.; et al. Mesenchymal stem cell-derived extracellular vesicles for immunomodulation and regeneration: A next generation therapeutic tool? Cell Death Dis. 2022, 13, 580. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Rhim, W.-K.; Seo, H.J.; Lee, J.Y.; Park, C.G.; Han, D.K. Comparative analysis of MSC-derived exosomes depending on cell culture media for regenerative bioactivity. Tissue Eng. Regen. Med. 2021, 18, 355–367. [Google Scholar] [CrossRef]
- Dayem, A.A.; Lee, S.B.; Lim, K.M.; Kim, A.; Shin, H.J.; Vellingiri, B.; Kim, Y.B.; Cho, S.-G. Bioactive peptides for boosting stem cell culture platform: Methods and applications. Biomed. Pharmacother. 2023, 160, 114376. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.-H.; Kim, J.-H. A Comprehensive review on factors influences biogenesis, functions, therapeutic and clinical implications of exosomes. Int. J. Nanomed. 2021, 16, 1281–1312. [Google Scholar] [CrossRef]
- Baldasici, O.; Pileczki, V.; Cruceriu, D.; Gavrilas, L.I.; Tudoran, O.; Balacescu, L.; Vlase, L.; Balacescu, O. Breast cancer-delivered exosomal miRNA as liquid biopsy biomarkers for metastasis prediction: A focus on translational research with clinical applicability. Int. J. Mol. Sci. 2022, 23, 9371. [Google Scholar] [CrossRef]
- Raj, D.A.; Fiume, I.; Capasso, G.; Pocsfalvi, G. A multiplex quantitative proteomics strategy for protein biomarker studies in urinary exosomes. Kidney Int. 2012, 81, 1263–1272. [Google Scholar] [CrossRef]
- Jafari, D.; Shajari, S.; Jafari, R.; Mardi, N.; Gomari, H.; Ganji, F.; Forouzandeh Moghadam, M.; Samadikuchaksaraei, A. Designer Exosomes: A new platform for biotechnology therapeutics. BioDrugs 2020, 34, 567–586. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Song, S.; Chen, N.; Liao, J.; Zeng, L. Stem cell-derived exosomes: A supernova in cosmetic dermatology. J. Cosmet. Dermatol. 2021, 20, 3812–3817. [Google Scholar] [CrossRef]
- Olumesi, K.R.; Goldberg, D.J. A review of exosomes and their application in cutaneous medical aesthetics. J. Cosmet. Dermatol. 2023, 22, 2628–2634. [Google Scholar] [CrossRef]
- Zhang, R.; Zhu, Y.; Li, Y.; Liu, W.; Yin, L.; Yin, S.; Ji, C.; Hu, Y.; Wang, Q.; Zhou, X.; et al. Human umbilical cord mesenchymal stem cell exosomes alleviate sepsis-associated acute kidney injury via regulating microRNA-146b expression. Biotechnol. Lett. 2020, 42, 669–679. [Google Scholar] [CrossRef]
- Li, T.; Yan, Y.; Wang, B.; Qian, H.; Zhang, X.; Shen, L.; Wang, M.; Zhou, Y.; Zhu, W.; Li, W.; et al. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate liver fibrosis. Stem Cells Dev. 2013, 22, 845–854. [Google Scholar] [CrossRef]
- Leggio, L.; Paternò, G.; Vivarelli, S.; L’episcopo, F.; Tirolo, C.; Raciti, G.; Pappalardo, F.; Giachino, C.; Caniglia, S.; Serapide, M.F.; et al. Extracellular vesicles as nanotherapeutics for parkinson’s disease. Biomolecules 2020, 10, 1327. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhang, B.; Yang, Y.; Jiang, Q.; Li, T.; Gong, J.; Tang, H.; Zhang, Q. Stem cell-derived exosomes: Emerging therapeutic opportunities for wound healing. Stem Cell Res. Ther. 2023, 14, 107. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhang, S.; Hu, H.; Yang, J.; Wang, X.; Ma, Y.; Jiang, J.; Wang, J.; Zhong, L.; Chen, M.; et al. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate acute liver failure by reducing the activity of the NLRP3 inflammasome in macrophages. Biochem. Biophys. Res. Commun. 2019, 508, 735–741. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef]
- Vlassov, A.V.; Magdaleno, S.; Setterquist, R.; Conrad, R. Exosomes: Current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2012, 1820, 940–948. [Google Scholar] [CrossRef]
- Kim, S.; Kang, G.H.; Lim, K.M.; Shin, Y.; Song, K.; Park, S.; An, J.; Kim, D.Y.; Shin, H.C.; Cho, S.G. Thermostable Human Basic Fibroblast Growth Factor (TS-bFGF) Engineered with a Disulfide Bond Demonstrates Superior Culture Outcomes in Human Pluripotent Stem Cell. Biology 2023, 12, 888. [Google Scholar] [CrossRef]
- Gharibi, B.; Hughes, F.J. Effects of medium supplements on proliferation, differentiation potential, and in vitro expansion of mesenchymal stem cells. STEM CELLS Transl. Med. 2012, 1, 771–782. [Google Scholar] [CrossRef]
- Saei Arezoumand, K.; Alizadeh, E.; Pilehvar-Soltanahmadi, Y.; Esmaeillou, M.; Zarghami, N. An overview on different strategies for the stemness maintenance of MSCs. Artif. Cells Nanomed. Biotechnol. 2017, 45, 1255–1271. [Google Scholar] [CrossRef]
- Chen, L.; Carlton, M.; Chen, X.; Kaur, N.; Ryan, H.; Parker, T.J.; Lin, Z.; Xiao, Y.; Zhou, Y. Effect of fibronectin, FGF-2, and BMP4 in the stemness maintenance of BMSCs and the metabolic and proteomic cues involved. Stem Cell Res. Ther. 2021, 12, 165. [Google Scholar] [CrossRef]
- Tesarova, L.; Jaresova, K.; Simara, P.; Koutna, I. Umbilical Cord-Derived Mesenchymal Stem Cells are Able to Use bFGF Treatment and Represent a Superb Tool for Immunosuppressive Clinical Applications. Int. J. Mol. Sci. 2020, 21, 5366. [Google Scholar] [CrossRef]
- Salehinejad, P.; Alitheen, N.B.; Mandegary, A.; Nematollahi-Mahani, S.N.; Janzamin, E. Effect of EGF and FGF on the expansion properties of human umbilical cord mesenchymal cells. Vitr. Cell Dev. Biol.-Anim. 2013, 49, 515–523. [Google Scholar] [CrossRef]
- Okamura, G.; Ebina, K.; Hirao, M.; Chijimatsu, R.; Yonetani, Y.; Etani, Y.; Miyama, A.; Takami, K.; Goshima, A.; Yoshikawa, H.; et al. Promoting effect of basic fibroblast growth factor in synovial mesenchymal stem cell-based cartilage regeneration. Int. J. Mol. Sci. 2020, 22, 300. [Google Scholar] [CrossRef]
- Liang, X.; Ding, Y.; Zhang, Y.; Tse, H.F.; Lian, Q. Paracrine Mechanisms of Mesenchymal Stem Cell-Based Therapy: Current Status and Perspectives. Cell Transplant. 2014, 23, 1045–1059. [Google Scholar] [CrossRef]
- Phinney, D.G.; Pittenger, M.F. Concise Review: MSC-Derived Exosomes for Cell-Free Therapy. Stem Cells 2017, 35, 851–858. [Google Scholar] [CrossRef]
- Bulati, M.; Gallo, A.; Zito, G.; Busà, R.; Iannolo, G.; Cuscino, N.; Castelbuono, S.; Carcione, C.; Centi, C.; Martucci, G.; et al. 3D Culture and Interferon-γ Priming Modulates Characteristics of Mesenchymal Stromal/Stem Cells by Modifying the Expression of Both Intracellular and Exosomal microRNAs. Biology 2023, 12, 1063. [Google Scholar] [CrossRef]
- Min Lim, K.; Kim, S.; Yeom, J.; Choi, Y.; Lee, Y.; An, J.; Gil, M.; Abdal Dayem, A.; Kim, K.; Kang, G.H.; et al. Advanced 3D dynamic culture system with transforming growth factor-beta3 enhances production of potent extracellular vesicles with modified protein cargoes via upregulation of TGF-beta signaling. J. Adv. Res. 2023, 47, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Eom, Y.W.; Oh, J.-E.; Lee, J.I.; Baik, S.K.; Rhee, K.-J.; Shin, H.C.; Kim, Y.M.; Ahn, C.M.; Kong, J.H.; Kim, H.S.; et al. The role of growth factors in maintenance of stemness in bone marrow-derived mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2014, 445, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Fujimiya, M. Potential effects of mesenchymal stem cell derived extracellular vesicles and exosomal miRNAs in neurological disorders. Neural Regen. Res. 2021, 16, 2359–2366. [Google Scholar] [CrossRef]
- Bray, E.R.; Oropallo, A.R.; Grande, D.A.; Kirsner, R.S.; Badiavas, E.V. Extracellular vesicles as therapeutic tools for the treatment of chronic wounds. Pharmaceutics 2021, 13, 1543. [Google Scholar] [CrossRef] [PubMed]
- Mottola, A.; Antoniotti, S.; Lovisolo, D.; Munaron, L. Regulation of noncapacitative calcium entry by arachidonic acid and nitric oxide in endothelial cells. FASEB J. 2005, 19, 2075–2077. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Davies, J.E.; Harper, M.T. 2-Aminoethoxydiphenylborate (2-APB) inhibits release of phosphatidylserine-exposing extracellular vesicles from platelets. Cell Death Discov. 2020, 6, 10. [Google Scholar] [CrossRef]
- Morrell, A.E.; Brown, G.N.; Robinson, S.T.; Sattler, R.L.; Baik, A.D.; Zhen, G.; Cao, X.; Bonewald, L.F.; Jin, W.; Kam, L.C.; et al. Mechanically induced Ca2+ oscillations in osteocytes release extracellular vesicles and enhance bone formation. Bone Res. 2018, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gu, Z.; Zhao, X.; Yang, N.; Wang, F.; Deng, A.; Zhao, S.; Luo, L.; Wei, H.; Guan, L.; et al. Extracellular Vesicles Released from Human Umbilical Cord-Derived Mesenchymal Stromal Cells Prevent Life-Threatening Acute Graft-Versus-Host Disease in a Mouse Model of Allogeneic Hematopoietic Stem Cell Transplantation. Stem Cells Dev. 2016, 25, 1874–1883. [Google Scholar] [CrossRef]
- Li, X.; Liu, L.; Yang, J.; Yu, Y.; Chai, J.; Wang, L.; Ma, L.; Yin, H. Exosome Derived From Human Umbilical Cord Mesenchymal Stem Cell Mediates MiR-181c Attenuating Burn-induced Excessive Inflammation. EBioMedicine 2016, 8, 72–82. [Google Scholar] [CrossRef]
- Joo, H.; Oh, M.-K.; Kang, J.Y.; Park, H.S.; Chae, D.-H.; Kim, J.; Lee, J.-H.; Yoo, H.M.; Choi, U.; Kim, D.-K.; et al. Extracellular vesicles from thapsigargin-treated mesenchymal stem cells ameliorated experimental colitis via enhanced immunomodulatory properties. Biomedicines 2021, 9, 209. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Species | Accession No. | Primer Sequence(5′ to 3′) | Product Size (bp) |
|---|---|---|---|---|
| GAPDH | Human | NM_001357943.2 | F: GTCTCCTCTGACTTCAACAGCG; R: ACCACCCTGTTGCTGTAGCCAA | 131 |
| OCT4 | Human | NM_203289.6 | F: CCTGAAGCAGAAGAGGATCACC; R: AAAGCGGCAGATGGTCGTTTGG | 106 |
| SOX2 | Human | NM_003106.4 | F: GCTACAGCATGATGCAGGACCA; R: TCTGCGAGCTGGTCATGGAGTT | 135 |
| KLF4 | Human | NM_001314052.2 | F: CATCTCAAGGCACACCTGCGAA; R: TCGGTCGCATTTTTGGCACTGG | 156 |
| IL-1β | Mouse | NM_008361.4 | F: TGGACCTTCCAGGATGAGGACA; R: GTTCATCTCGGAGCCTGTAGTG | 148 |
| TNF-α | Mouse | NM_013693.3 | F: GGTGCCTATGTCTCAGCCTCTT; R: GCCATAGAACTGATGAGAGGGAG | 139 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.; Kim, S.; Lim, K.; Shin, Y.; Song, K.; Kang, G.-H.; Kim, D.Y.; Shin, H.-C.; Cho, S.-G. Thermostable Basic Fibroblast Growth Factor Enhances the Production and Activity of Human Wharton’s Jelly Mesenchymal Stem Cell-Derived Extracellular Vesicles. Int. J. Mol. Sci. 2023, 24, 16460. https://doi.org/10.3390/ijms242216460
Park S, Kim S, Lim K, Shin Y, Song K, Kang G-H, Kim DY, Shin H-C, Cho S-G. Thermostable Basic Fibroblast Growth Factor Enhances the Production and Activity of Human Wharton’s Jelly Mesenchymal Stem Cell-Derived Extracellular Vesicles. International Journal of Molecular Sciences. 2023; 24(22):16460. https://doi.org/10.3390/ijms242216460
Chicago/Turabian StylePark, SangRok, SeJong Kim, KyungMin Lim, YeoKyung Shin, Kwonwoo Song, Geun-Ho Kang, Dae Young Kim, Hang-Cheol Shin, and Ssang-Goo Cho. 2023. "Thermostable Basic Fibroblast Growth Factor Enhances the Production and Activity of Human Wharton’s Jelly Mesenchymal Stem Cell-Derived Extracellular Vesicles" International Journal of Molecular Sciences 24, no. 22: 16460. https://doi.org/10.3390/ijms242216460
APA StylePark, S., Kim, S., Lim, K., Shin, Y., Song, K., Kang, G.-H., Kim, D. Y., Shin, H.-C., & Cho, S.-G. (2023). Thermostable Basic Fibroblast Growth Factor Enhances the Production and Activity of Human Wharton’s Jelly Mesenchymal Stem Cell-Derived Extracellular Vesicles. International Journal of Molecular Sciences, 24(22), 16460. https://doi.org/10.3390/ijms242216460