
asb5a/asb5b Double Knockout Affects Zebrafish Cardiac Contractile Function

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
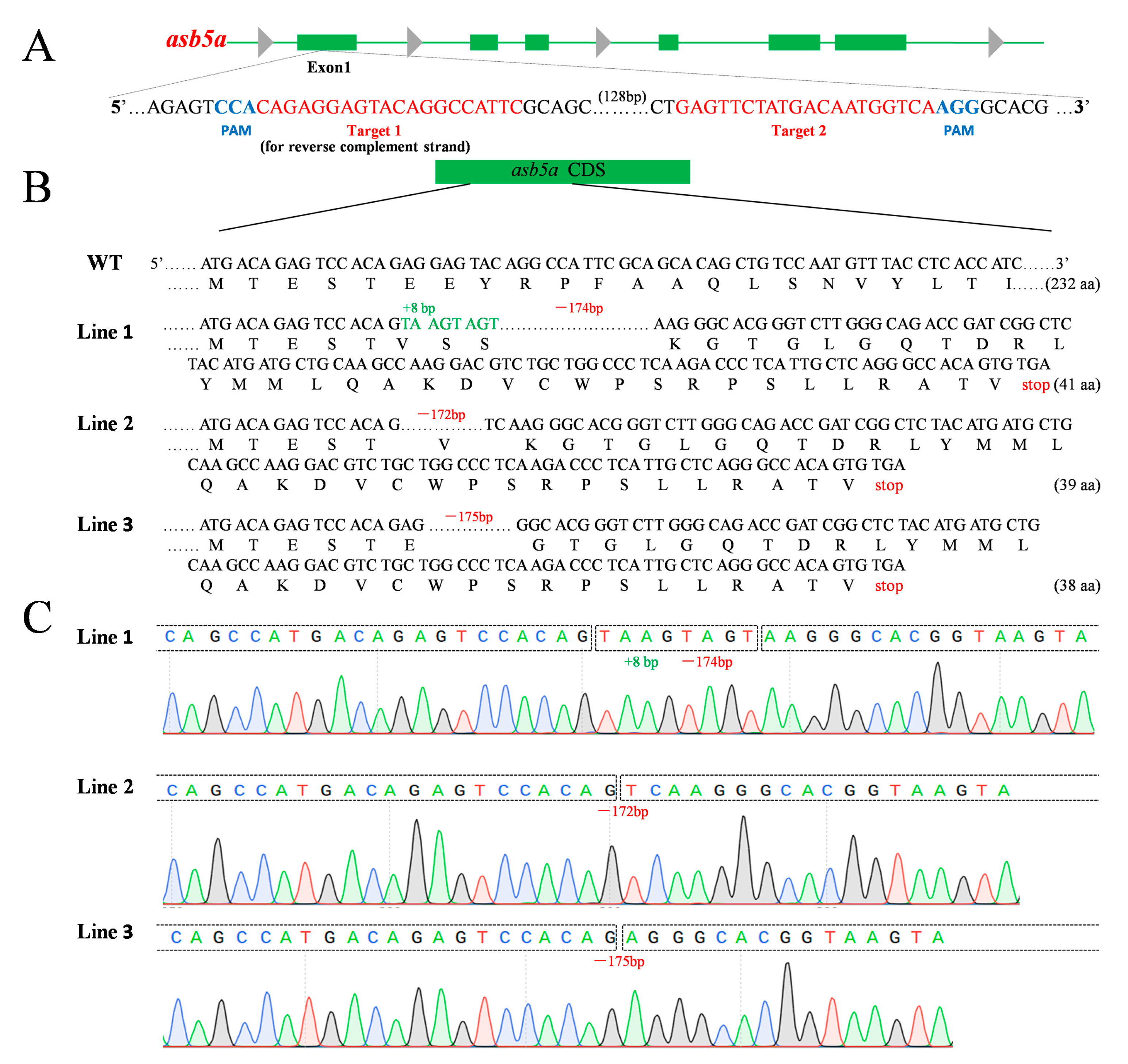
2.1. Construction of asb5a-Knockout Zebrafish Line
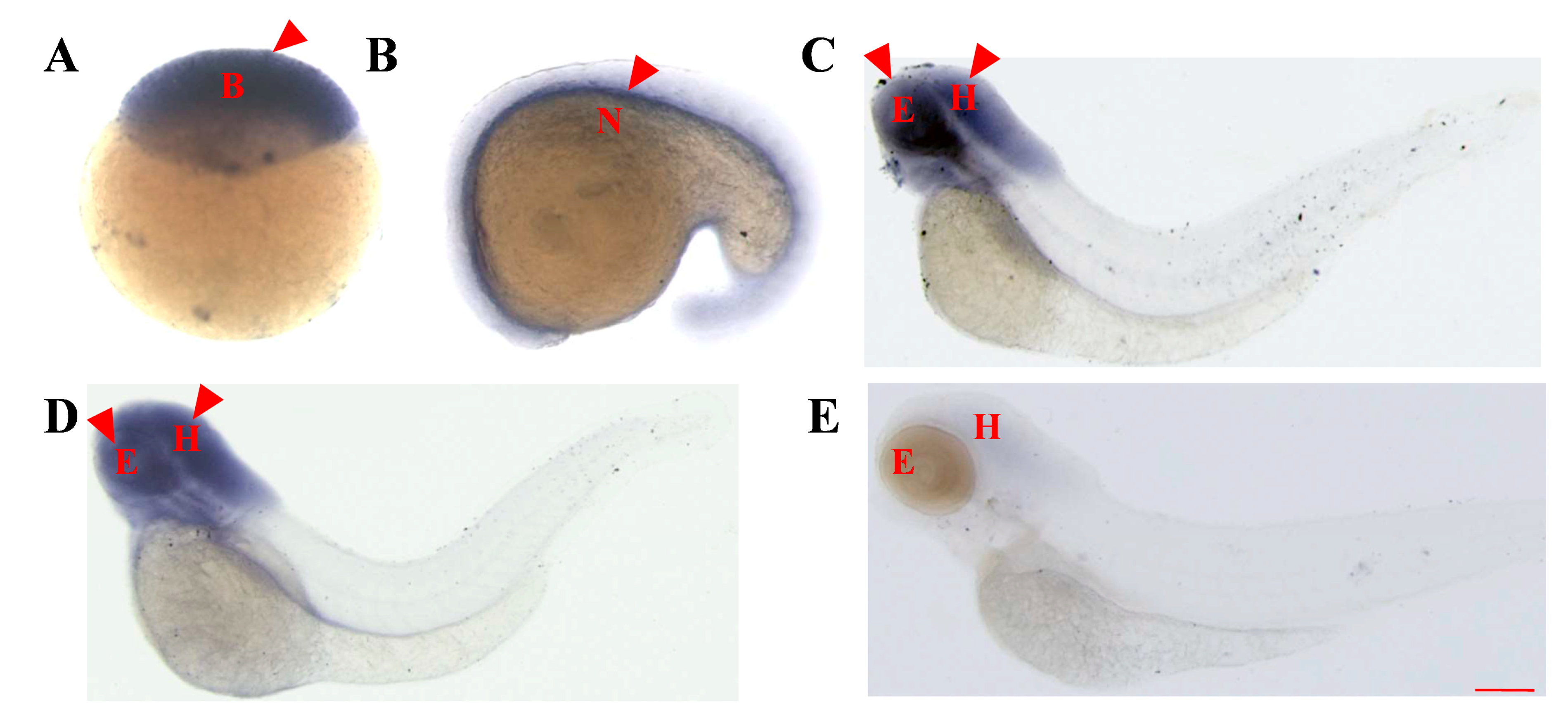
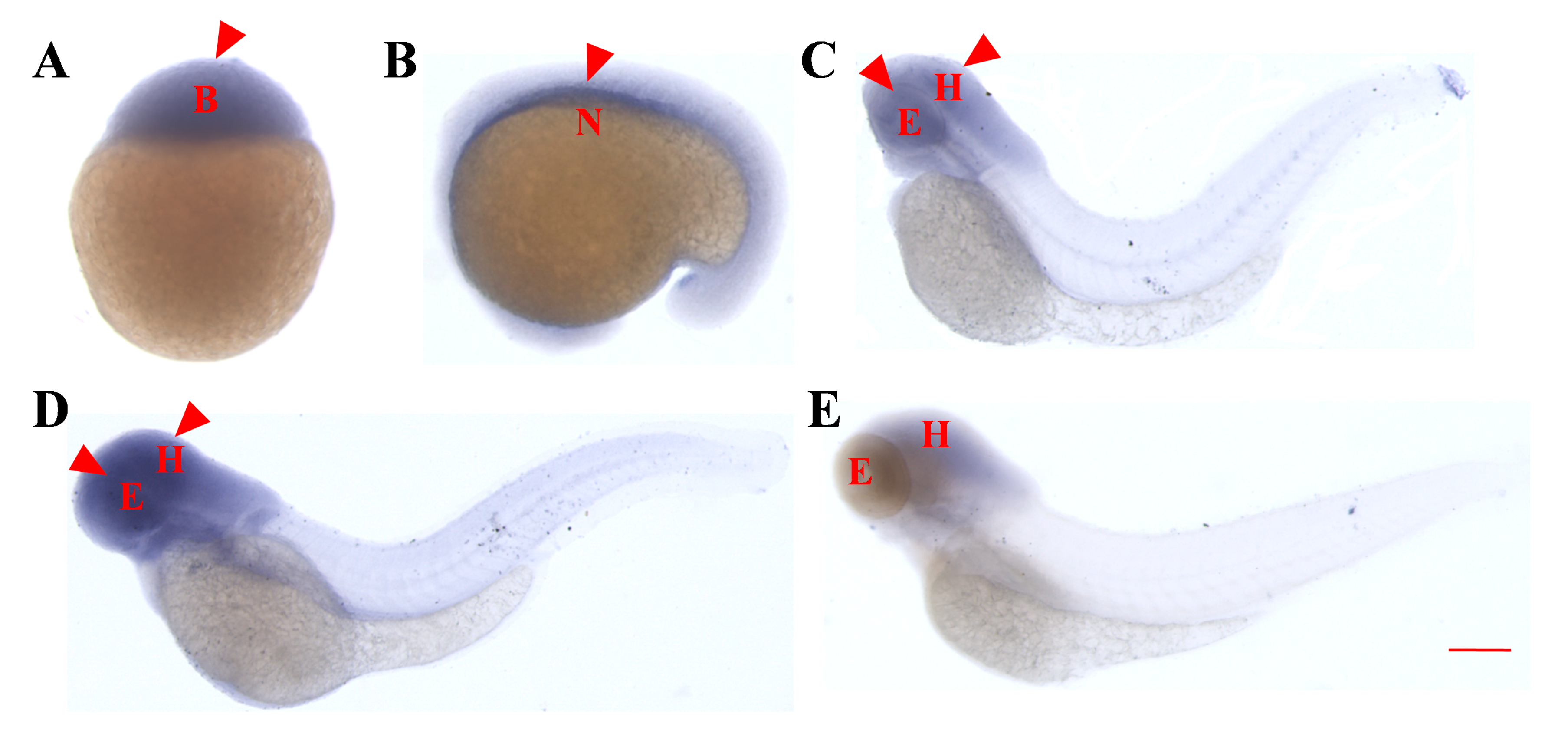
2.2. Whole-Embryo In Situ Hybridization Reveals the Similar Early Expression Patterns of asb5a and asb5b
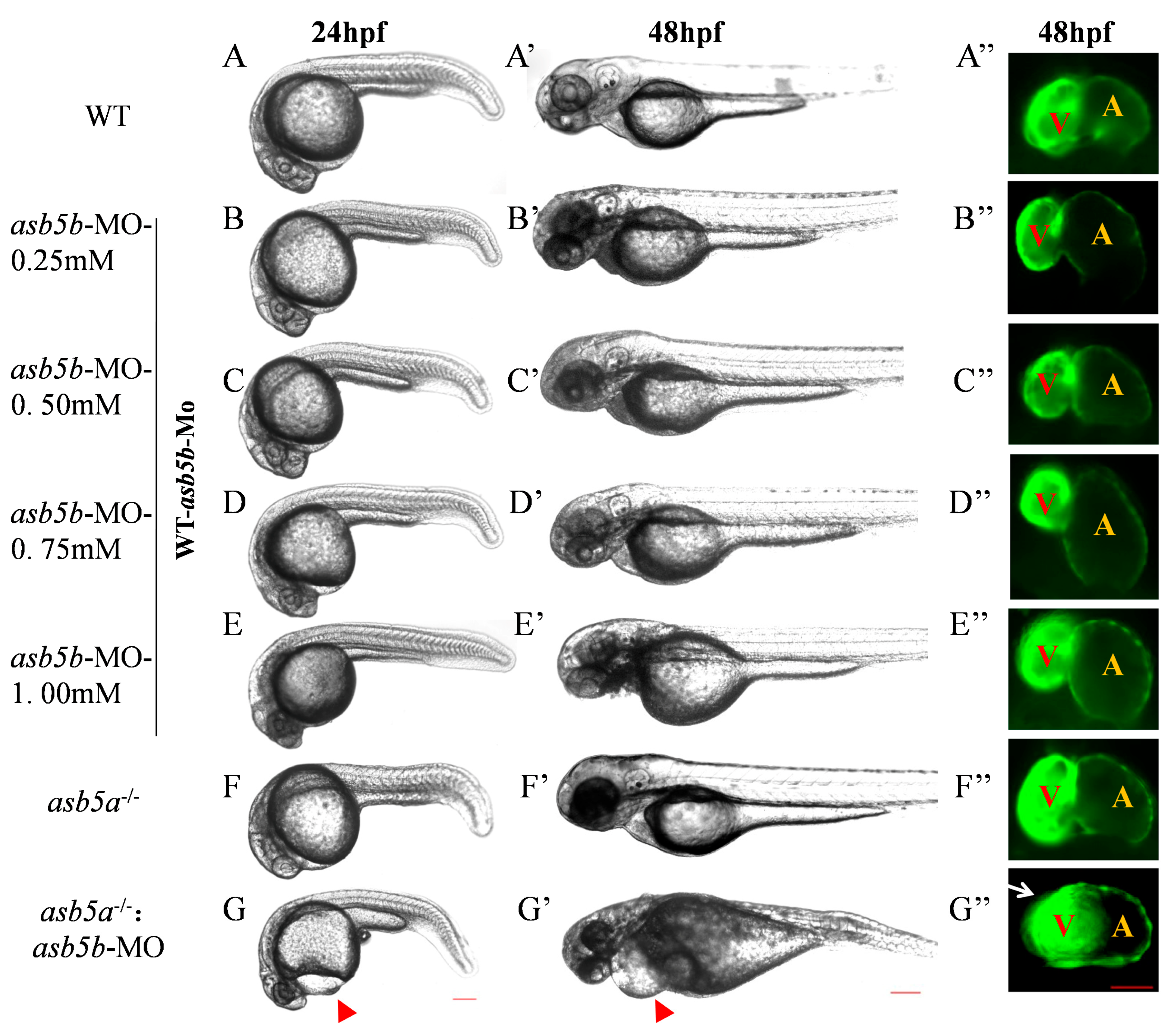
2.3. Construction of asb5a−/−:asb5b-ATGmo Double-Knockout Zebrafish Strain
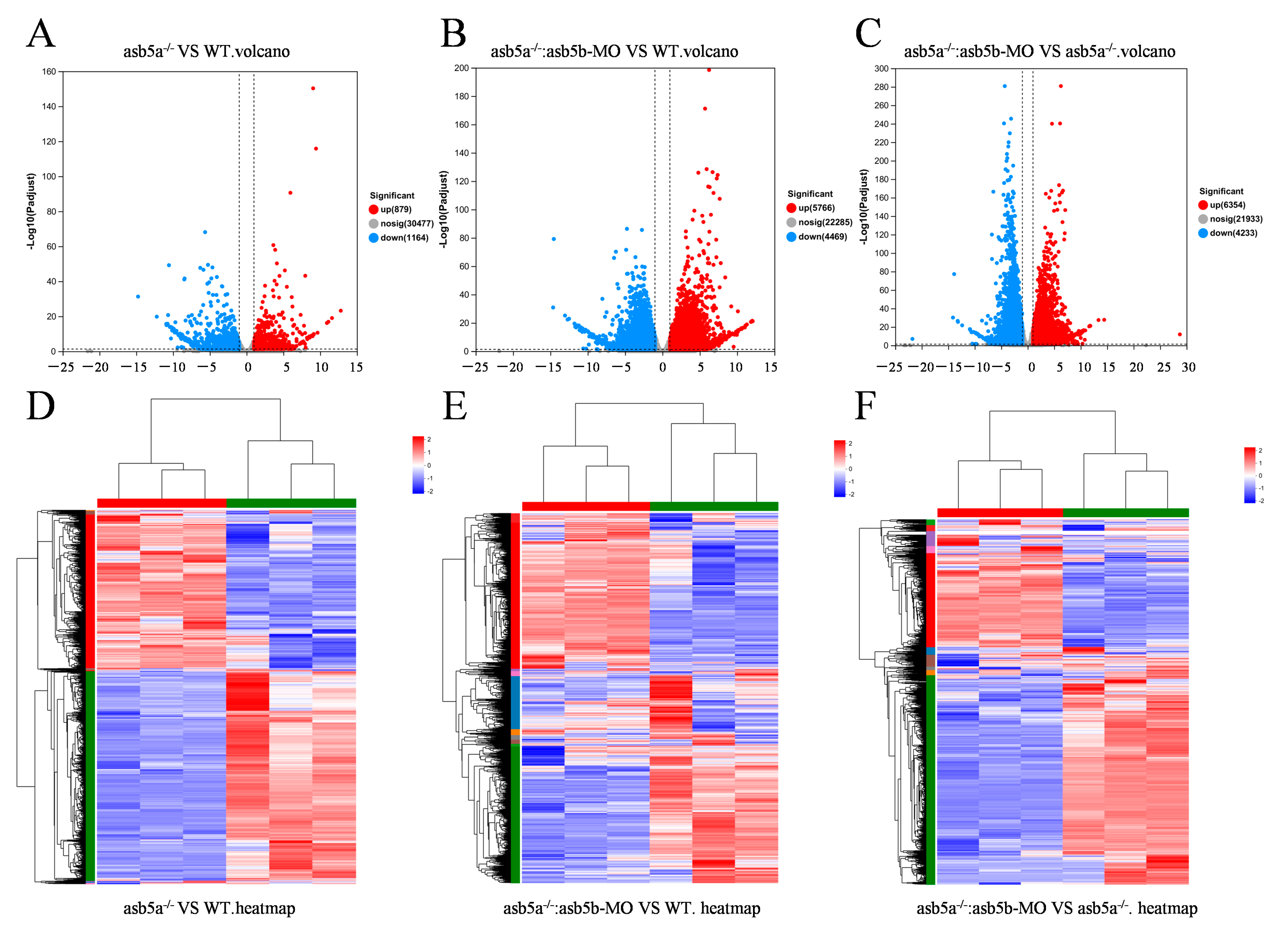
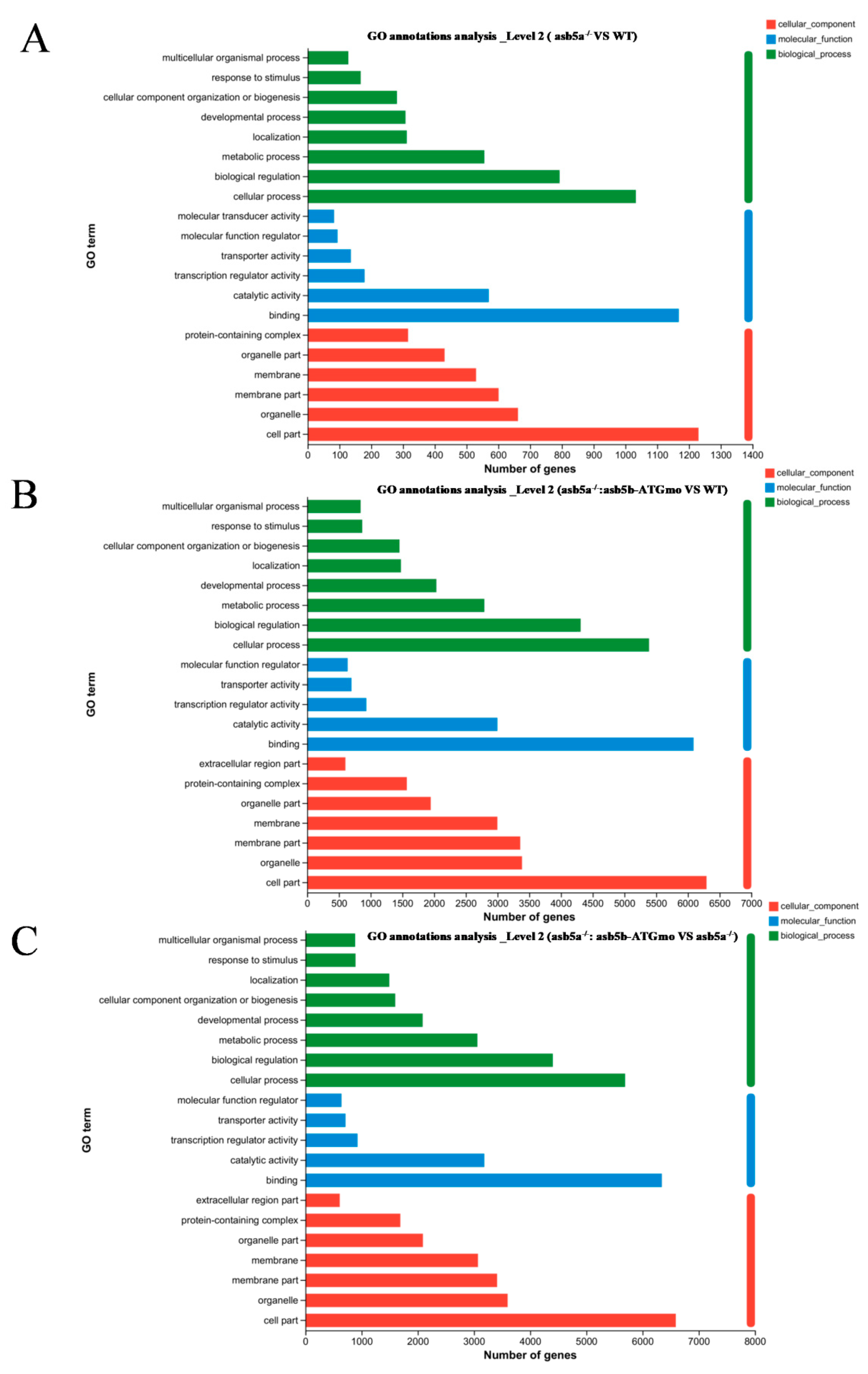
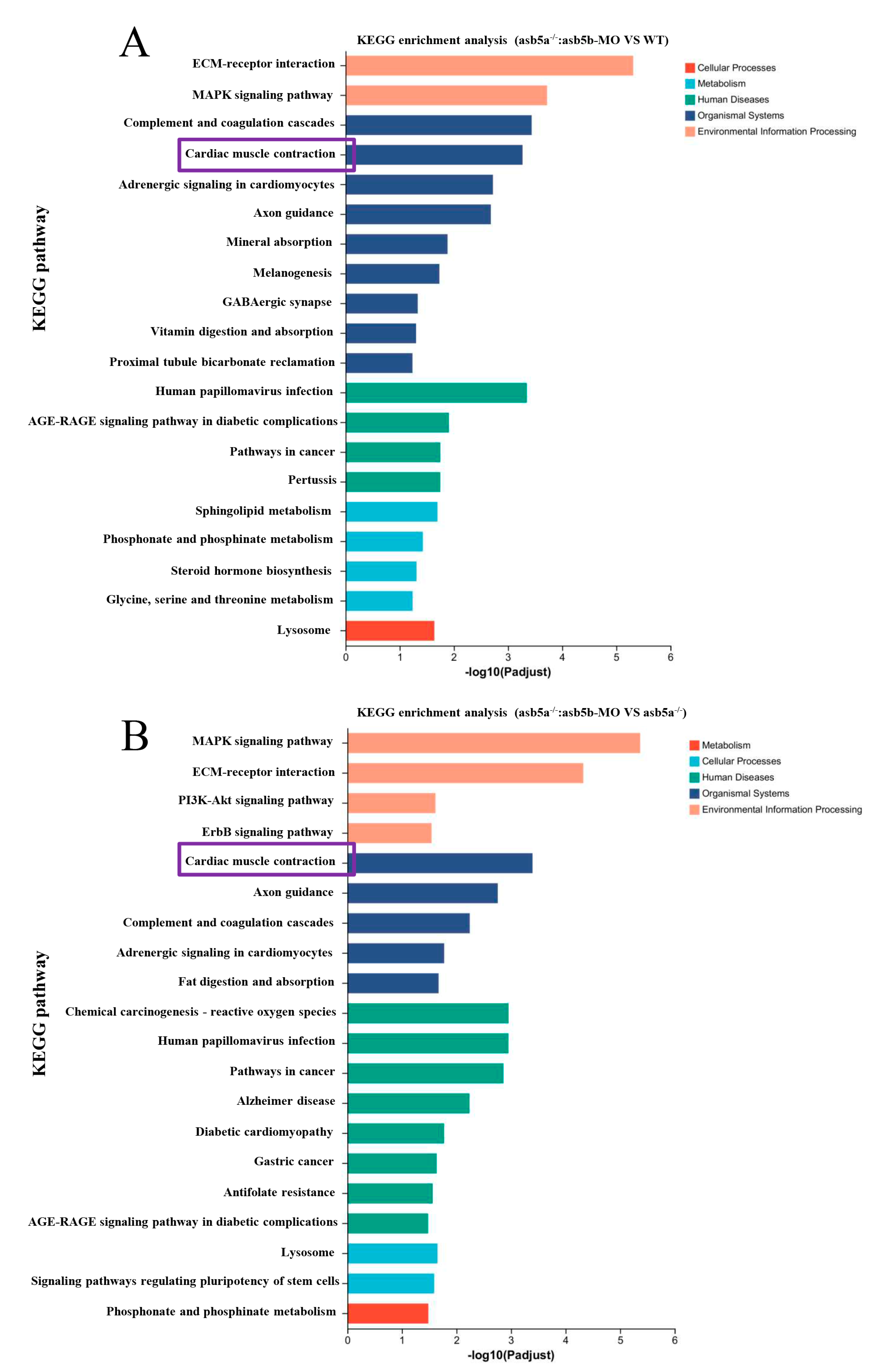
2.4. Transcriptome Sequencing Data Analysis
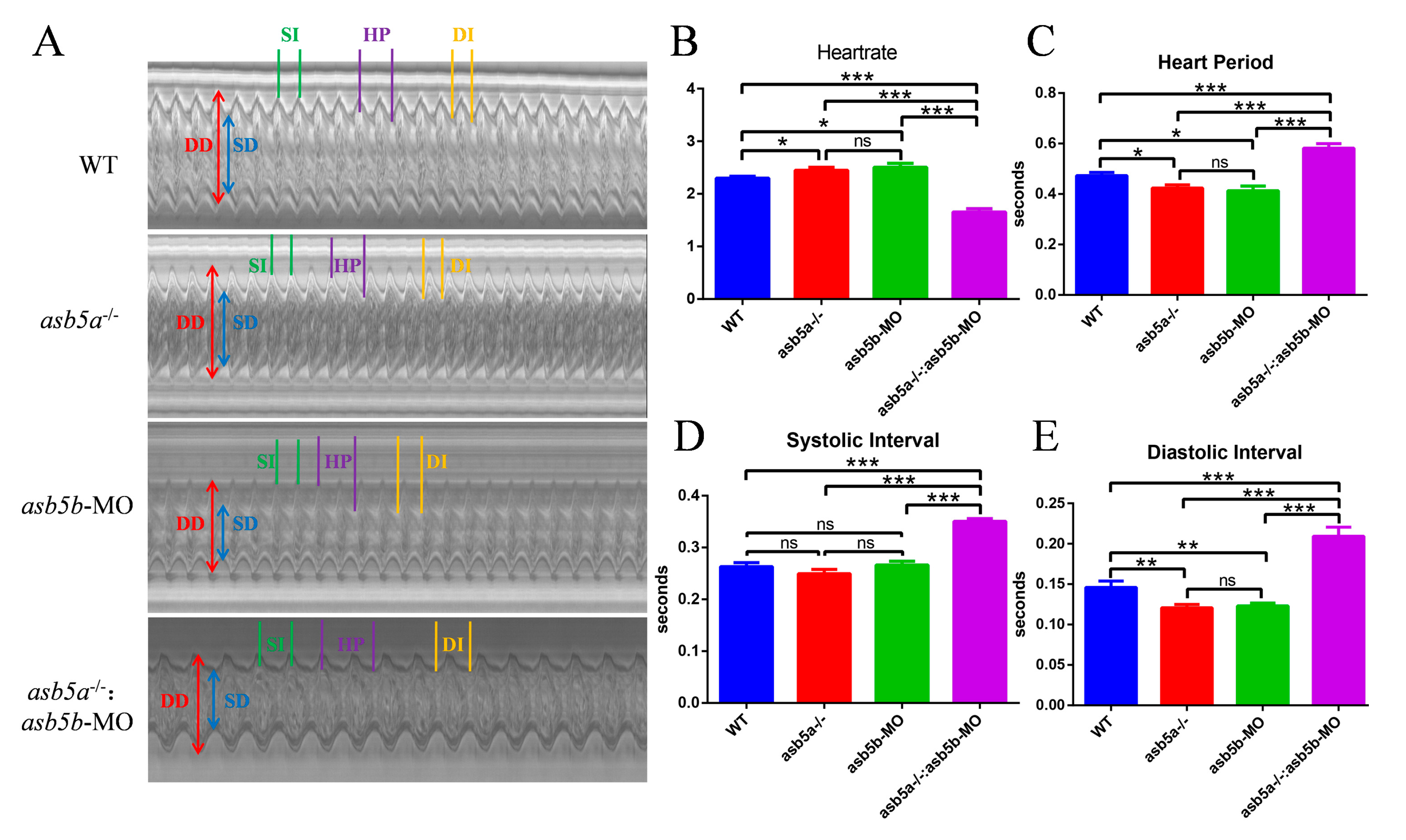
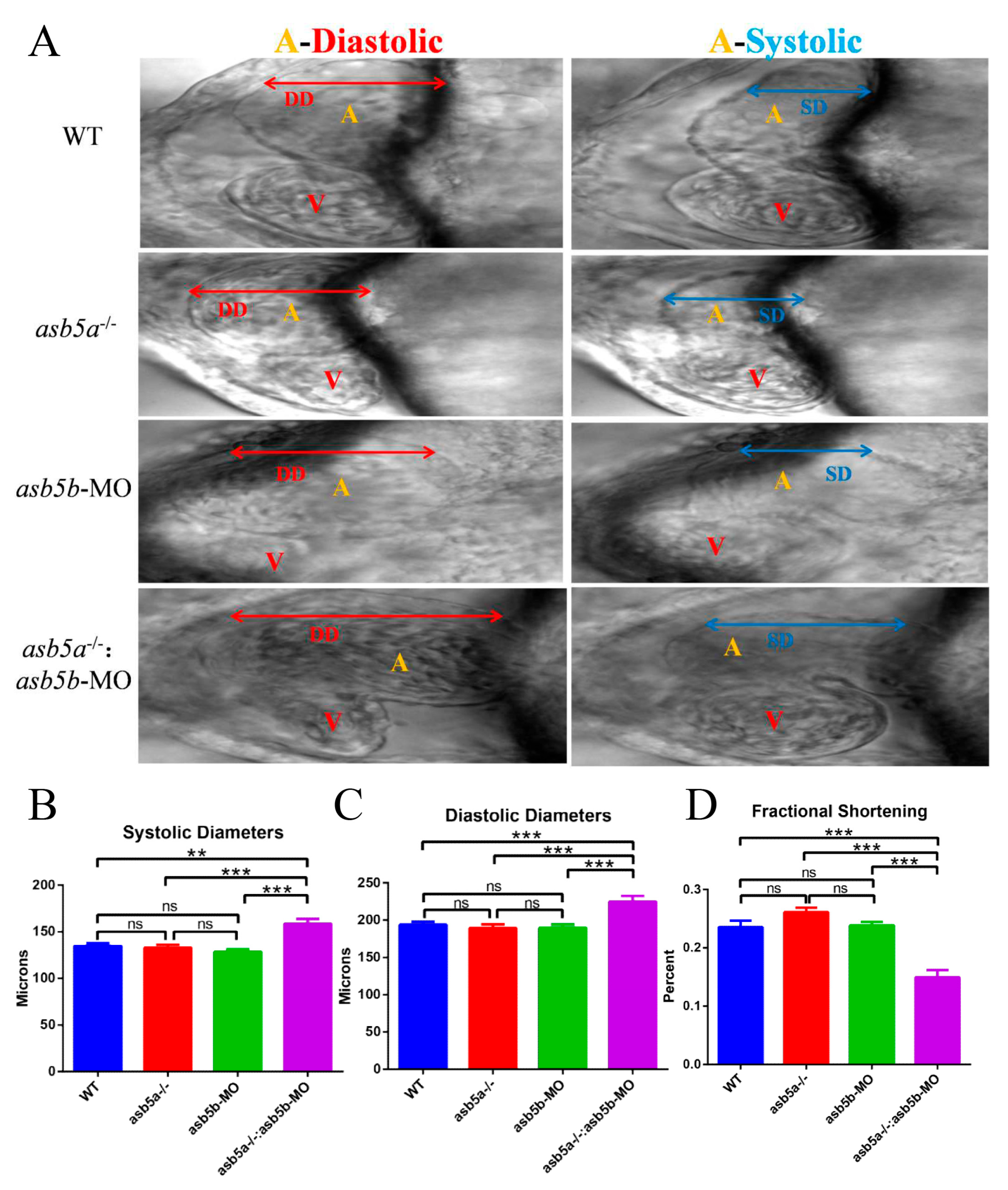
2.5. Early Cardiac Contraction Abnormalities Induced by asb5a−/−:asb5b-ATGmo Double-Knockout in Zebrafish
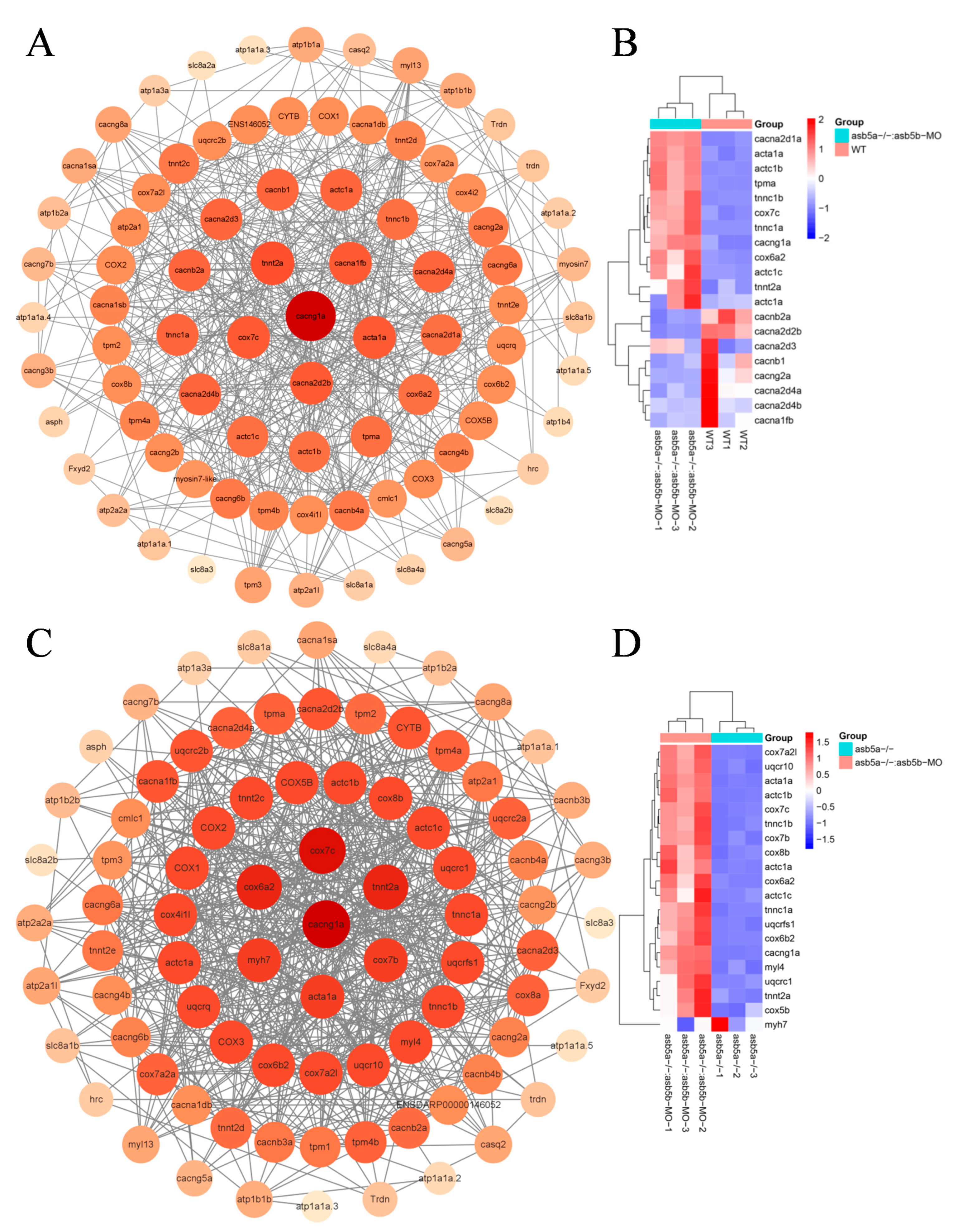
2.6. PPI Network Construction and Hub Gene Screening
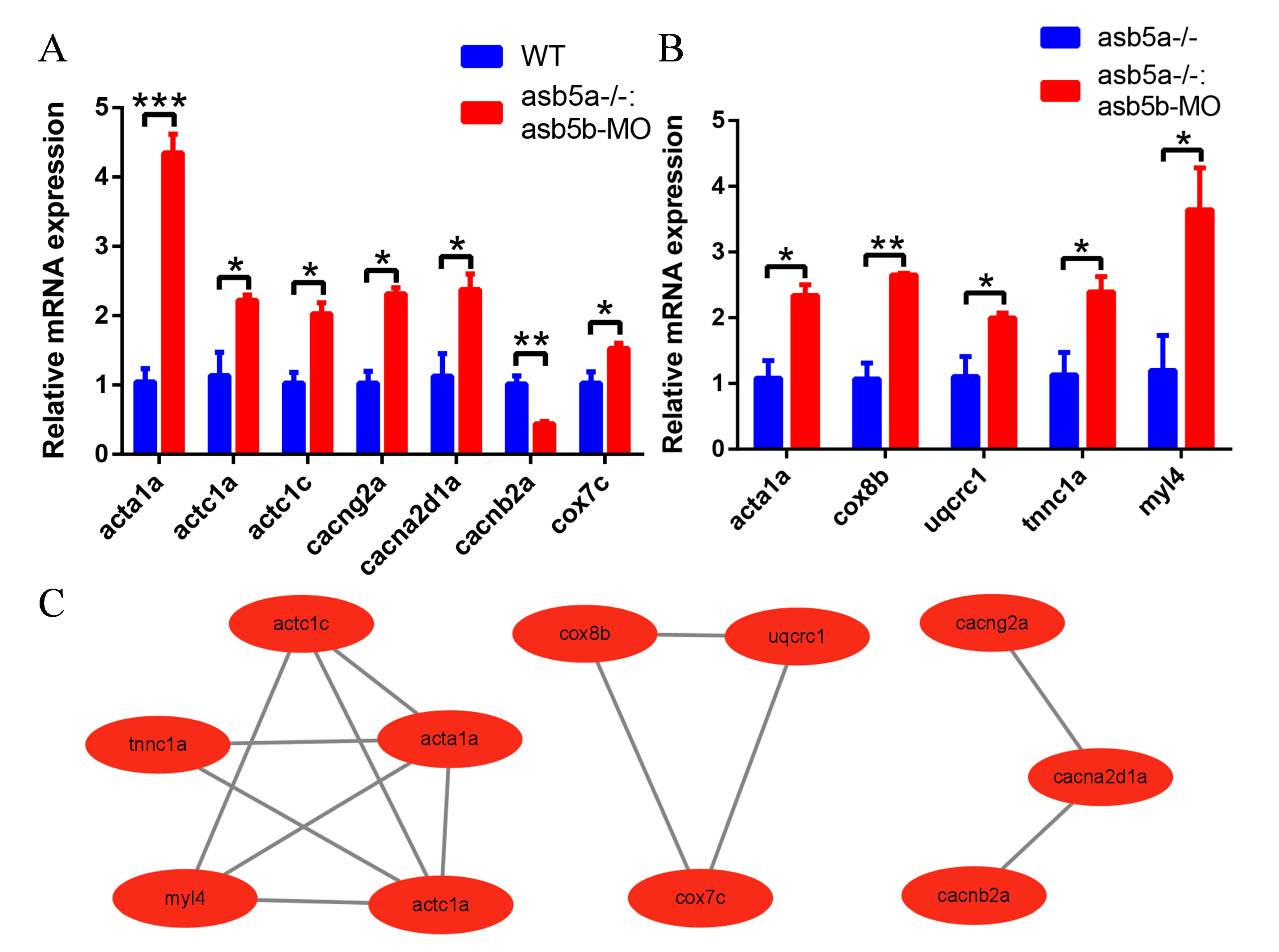
2.7. The qPCR Validation of 20 Abnormally Regulated Hub Genes
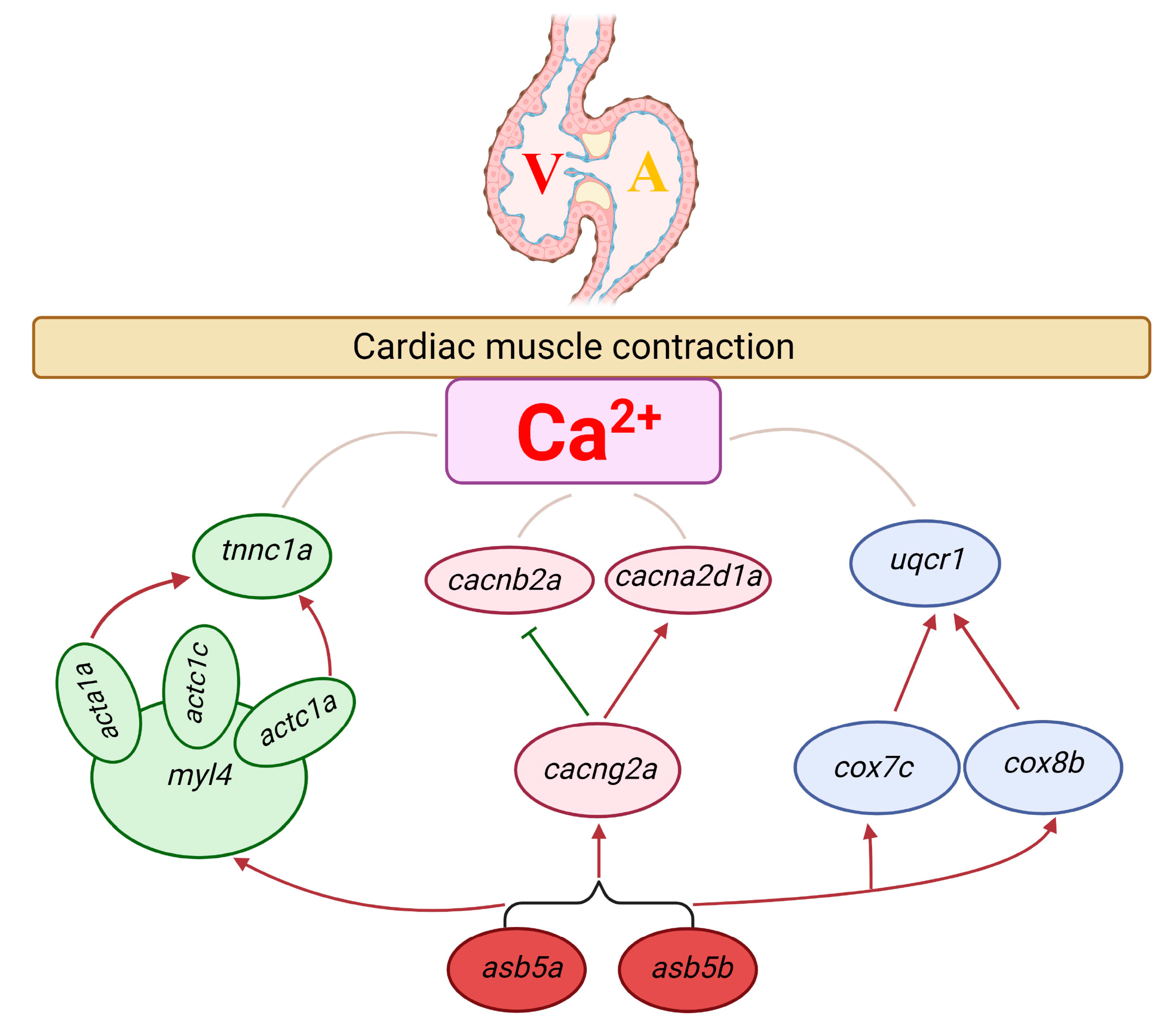
3. Discussion
4. Materials and Methods
4.1. Zebrafish Strain Rearing and Breeding
4.2. CRISPR/Cas9-Mediated asb5a Gene Knockout
4.3. RNA Probe Synthesis and WISH
4.4. Morpholino-Mediated Specific Knockdown of the asb5b Gene
4.5. RNA-Seq-Based Transcriptome Analysis
4.6. Imaging Technique and Image Analysis
4.7. PPI Network Construction and Module Selection
4.8. The qPCR detection
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, P.; Verhaar, A.P.; Peppelenbosch, M.P. Signaling Size: Ankyrin and SOCS Box-Containing ASB E3 Ligases in Action. Trends Biochem. Sci. 2019, 44, 64–74. [Google Scholar] [CrossRef]
- Boengler, K.; Pipp, F.; Fernandez, B.; Richter, A.; Schaper, W.; Deindl, E. The ankyrin repeat containing SOCS box protein 5: A novel protein associated with arteriogenesis. Biochem. Biophys. Res. Commun. 2003, 302, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, K.C.; Lacey, M.; Ehrlich, M. Epigenetics of Skeletal Muscle-Associated Genes in the ASB, LRRC, TMEM, and OSBPL Gene Families. Epigenomes 2020, 4, 1. [Google Scholar] [CrossRef]
- Jensen, J.H.; Conley, L.N.; Hedegaard, J.; Nielsen, M.; Young, J.F.; Oksbjerg, N.; Hornshøj, H.; Bendixen, C.; Thomsen, B. Gene expression profiling of porcine skeletal muscle in the early recovery phase following acute physical activity. Exp. Physiol. 2012, 97, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.X.; Garcia-Gras, E.; Wycuff, D.R.; Marriot, S.J.; Kadeer, N.; Yu, W.; Olson, E.N.; Garry, D.J.; Parmacek, M.S.; Schwartz, R.J. Identification of Direct Serum-response Factor Gene Targets during Me2SO-induced P19 Cardiac Cell Differentiation. J. Biol. Chem. 2005, 280, 19115–19126. [Google Scholar] [CrossRef] [PubMed]
- Albadri, S.; Del Bene, F.; Revenu, C. Genome editing using CRISPR/Cas9-based knock-in approaches in zebrafish. Methods 2017, 121–122, 77–85. [Google Scholar] [CrossRef]
- Sharma, P.; Sharma, B.S.; Verma, R.J. CRISPR-based genome editing of zebrafish. Prog. Mol. Biol. Transl. Sci. 2021, 180, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, B.; Luo, L.; Shi, D.-L.; Wang, H.; Cui, Z.; Huang, H.; Cao, Y.; Shu, X.; Zhang, W.; et al. Systematic genome editing of the genes on zebrafish Chromosome 1 by CRISPR/Cas9. Genome Res. 2019, 30, 118–126. [Google Scholar] [CrossRef]
- Thisse, C.; Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 2007, 3, 59–69. [Google Scholar] [CrossRef]
- Li, B.; Chen, K.; Liu, F.; Zhang, J.; Chen, X.; Chen, T.; Chen, Q.; Yao, Y.; Hu, W.; Wang, L.; et al. Developmental Angiogenesis Requires the Mitochondrial Phenylalanyl-tRNA Synthetase. Front. Cardiovasc. Med. 2021, 8, 724846. [Google Scholar] [CrossRef]
- Ujibe, K.; Nishimura, K.; Kashima, M.; Hirata, H. Direct-TRI: High-throughput RNA-extracting Method for all Stages of Zebrafish Development. Bio-Protocol 2021, 11, e4136. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhao, W.; Zhang, S.; Ma, J.; Sultan, Y.; Li, X. High-throughput transcriptome sequencing reveals the key stages of cardiovascular development in zebrafish embryos. BMC Genom. 2022, 23, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ocorr, K.; Fink, M.; Cammarato, A.; Bernstein, S.I.; Bodmer, R. Semi-automated Optical Heartbeat Analysis of Small Hearts. J. Vis. Exp. 2009, 31, e1435. [Google Scholar] [CrossRef]
- Gregorich, Z.R.; Cai, W.; Lin, Z.; Chen, A.J.; Peng, Y.; Kohmoto, T.; Ge, Y. Distinct sequences and post-translational modifications in cardiac atrial and ventricular myosin light chains revealed by top-down mass spectrometry. J. Mol. Cell. Cardiol. 2017, 107, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Morano, I.; Haase, H. Different actin affinities of human cardiac essential myosin light chain isoforms. FEBS Lett. 1997, 408, 71–74. [Google Scholar] [CrossRef]
- El-Mezgueldi, M. Calponin. Int. J. Biochem. Cell Biol. 1996, 28, 1185–1189. [Google Scholar] [CrossRef]
- Metskas, L.A.; Rhoades, E. Order–Disorder Transitions in the Cardiac Troponin Complex. J. Mol. Biol. 2016, 428, 2965–2977. [Google Scholar] [CrossRef]
- Acosta, L.; Haase, H.; Morano, I.; Moorman, A.F.; Franco, D. Regional expression of L-type calcium channel subunits during cardiac development. Dev. Dyn. 2004, 230, 131–136. [Google Scholar] [CrossRef]
- Chernyavskaya, Y.; Ebert, A.M.; Milligan, E.; Garrity, D.M. Voltage-gated calcium channel CACNB2 (β2.1) protein is required in the heart for control of cell proliferation and heart tube integrity. Dev. Dyn. 2012, 241, 648–662. [Google Scholar] [CrossRef]
- Wu, H.; Kaur, G.; Griffiths, G.L. An improved synthesis of a fluorescent gabapentin–choline conjugate for single molecule detection. Tetrahedron Lett. 2009, 50, 2100–2102. [Google Scholar] [CrossRef][Green Version]
- Ramzan, R.; Kadenbach, B.; Vogt, S. Multiple Mechanisms Regulate Eukaryotic Cytochrome C Oxidase. Cells 2021, 10, 514. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.; Niu, Y.; Zhang, X.; Zhang, J.; Wang, X.; Shen, X.; Chen, B.; Yu, J. Integrative Proteomic and Metabolomic Analysis Reveals Metabolic Phenotype in Mice with Cardiac-Specific Deletion of Natriuretic Peptide Receptor A. Mol. Cell. Proteom. 2021, 20, 100072. [Google Scholar] [CrossRef] [PubMed]
- Yi, T.; Chen, H.; Zhan, J.; Li, Y.; Long, Z.; Wu, Z.; Yang, M.; Peng, T.; Li, H. Ubiquinol-cytochrome c reductase core protein 1 contributes to cardiac tolerance to acute exhaustive exercise. Exp. Biol. Med. 2021, 247, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, T.P. Demographic model for inheritable cardiac disease. Arch. Biochem. Biophys. 2019, 672, 108056. [Google Scholar] [CrossRef]
- Chong, J.X.; Childers, M.C.; Marvin, C.T.; Marcello, A.J.; Gonorazky, H.; Hazrati, L.-N.; Dowling, J.J.; Al Amrani, F.; Alanay, Y.; Nieto, Y.; et al. Variants in ACTC1 underlie distal arthrogryposis accompanied by congenital heart defects. Hum. Genet. Genom. Adv. 2023, 4, 100213. [Google Scholar] [CrossRef]
- Lindqvist, J.; Levy, Y.; Pati-Alam, A.; Hardeman, E.C.; Gregorevic, P.; Ochala, J. Modulating myosin restores muscle function in a mouse model of nemaline myopathy. Ann. Neurol. 2016, 79, 717–725. [Google Scholar] [CrossRef]
- Zhu, X.-Y.; Xia, B.; Wu, Y.-Y.; Yang, H.; Li, C.-Q.; Li, P. Fenobucarb induces heart failure and cerebral hemorrhage in zebrafish. Aquat. Toxicol. 2019, 209, 34–41. [Google Scholar] [CrossRef]
- Yu, M.; Ji, L.; Li, S.; Zhang, Y.; Sun, X.; Sun, R.; Chen, X.; Luo, Y.; Cui, X. Exosomal circ-CACNG2 promotes cardiomyocyte apoptosis in multiple myeloma via modulating miR-197-3p/caspase3 axis. Exp. Cell Res. 2022, 417, 113229. [Google Scholar] [CrossRef]
- Briot, J.; Tétreault, M.P.; Bourdin, B.; Parent, L. Inherited Ventricular Arrhythmias: The Role of the Multi-Subunit Structure of the L-Type Calcium Channel Complex. Adv. Exp. Med. Biol. 2017, 966, 55–64. [Google Scholar] [CrossRef]
- Zhou, W.; Zhang, K.; Chen, D.; Gao, P.; Wang, Q. Gene microarray analyses for potential biomarkers of single and recurrent venous thromboembolism. Mol. Med. Rep. 2015, 12, 7358–7366. [Google Scholar] [CrossRef]
- Rocca, C.J.; Goodman, S.M.; Dulin, J.N.; Haquang, J.H.; Gertsman, I.; Blondelle, J.; Smith, J.L.M.; Heyser, C.J.; Cherqui, S. Transplantation of wild-type mouse hematopoietic stem and progenitor cells ameliorates deficits in a mouse model of Friedreich’s ataxia. Sci. Transl. Med. 2017, 9, eaaj2347. [Google Scholar] [CrossRef] [PubMed]
- Carroll, D. Genome Engineering with Targetable Nucleases. Annu. Rev. Biochem. 2014, 83, 409–439. [Google Scholar] [CrossRef] [PubMed]
- Kamachi, Y.; Kawahara, A. CRISPR-Cas9-Mediated Genome Modifications in Zebrafish. Methods Mol. Biol. (Clifton N.J.) 2023, 2637, 313–324. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2009, 26, 136–138. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, W.; Wang, Y.; Luo, Y.; Gao, L.; Zhang, J.; Jiang, Z.; Fan, X.; Li, F.; Xie, Y.; Wu, X.; et al. asb5a/asb5b Double Knockout Affects Zebrafish Cardiac Contractile Function. Int. J. Mol. Sci. 2023, 24, 16364. https://doi.org/10.3390/ijms242216364
Cai W, Wang Y, Luo Y, Gao L, Zhang J, Jiang Z, Fan X, Li F, Xie Y, Wu X, et al. asb5a/asb5b Double Knockout Affects Zebrafish Cardiac Contractile Function. International Journal of Molecular Sciences. 2023; 24(22):16364. https://doi.org/10.3390/ijms242216364
Chicago/Turabian StyleCai, Wanwan, Yuequn Wang, Ying Luo, Luoqing Gao, Jian Zhang, Zhigang Jiang, Xiongwei Fan, Fang Li, Yulian Xie, Xiushan Wu, and et al. 2023. "asb5a/asb5b Double Knockout Affects Zebrafish Cardiac Contractile Function" International Journal of Molecular Sciences 24, no. 22: 16364. https://doi.org/10.3390/ijms242216364
APA StyleCai, W., Wang, Y., Luo, Y., Gao, L., Zhang, J., Jiang, Z., Fan, X., Li, F., Xie, Y., Wu, X., Li, Y., & Yuan, W. (2023). asb5a/asb5b Double Knockout Affects Zebrafish Cardiac Contractile Function. International Journal of Molecular Sciences, 24(22), 16364. https://doi.org/10.3390/ijms242216364