
Physiological and Transcriptomic Analysis Provides Insights into Low Nitrogen Stress in Foxtail Millet (Setaria italica L.)

 , ,
, ,
Abstract
:1. Introduction
2. Results
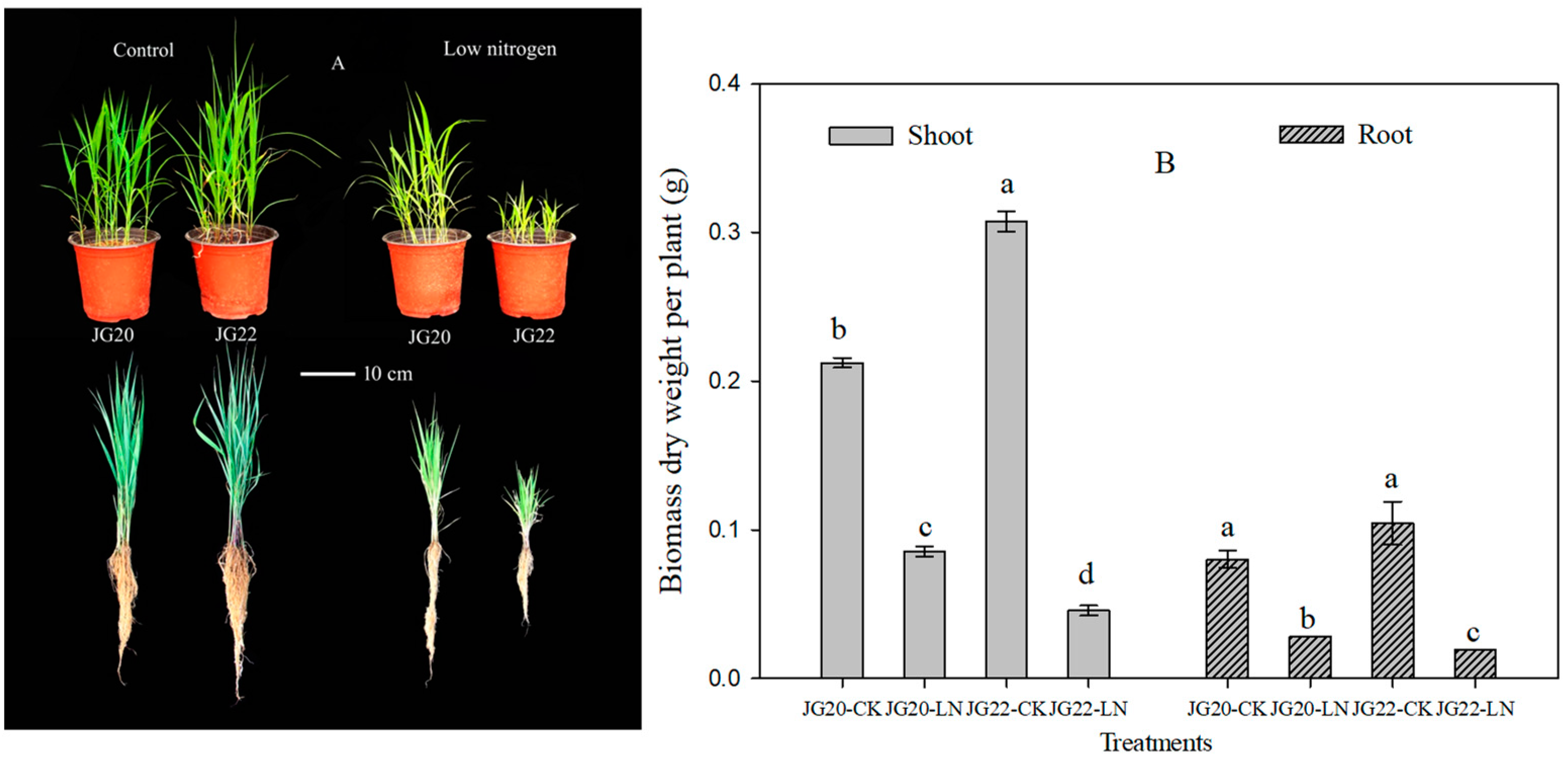
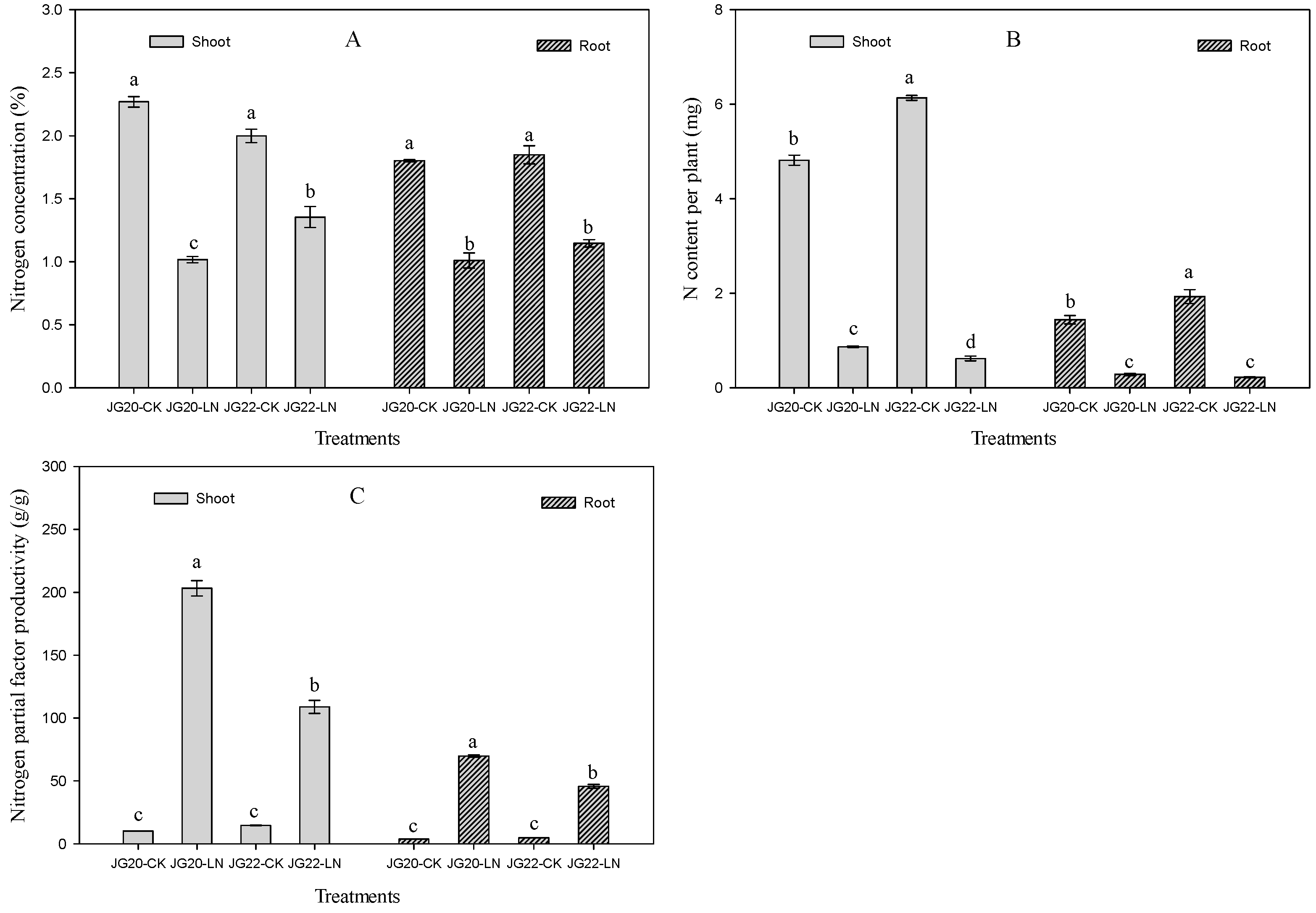
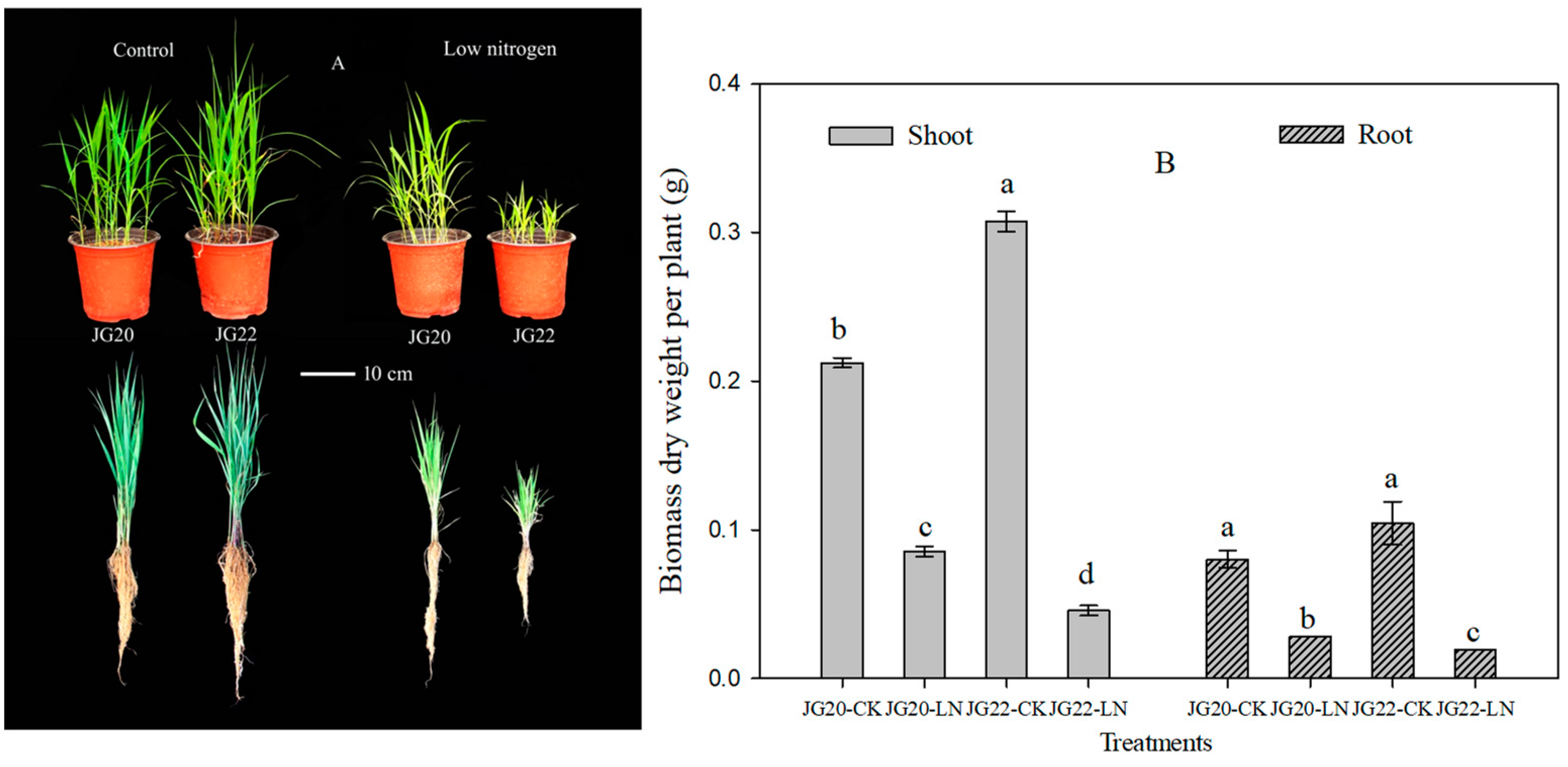
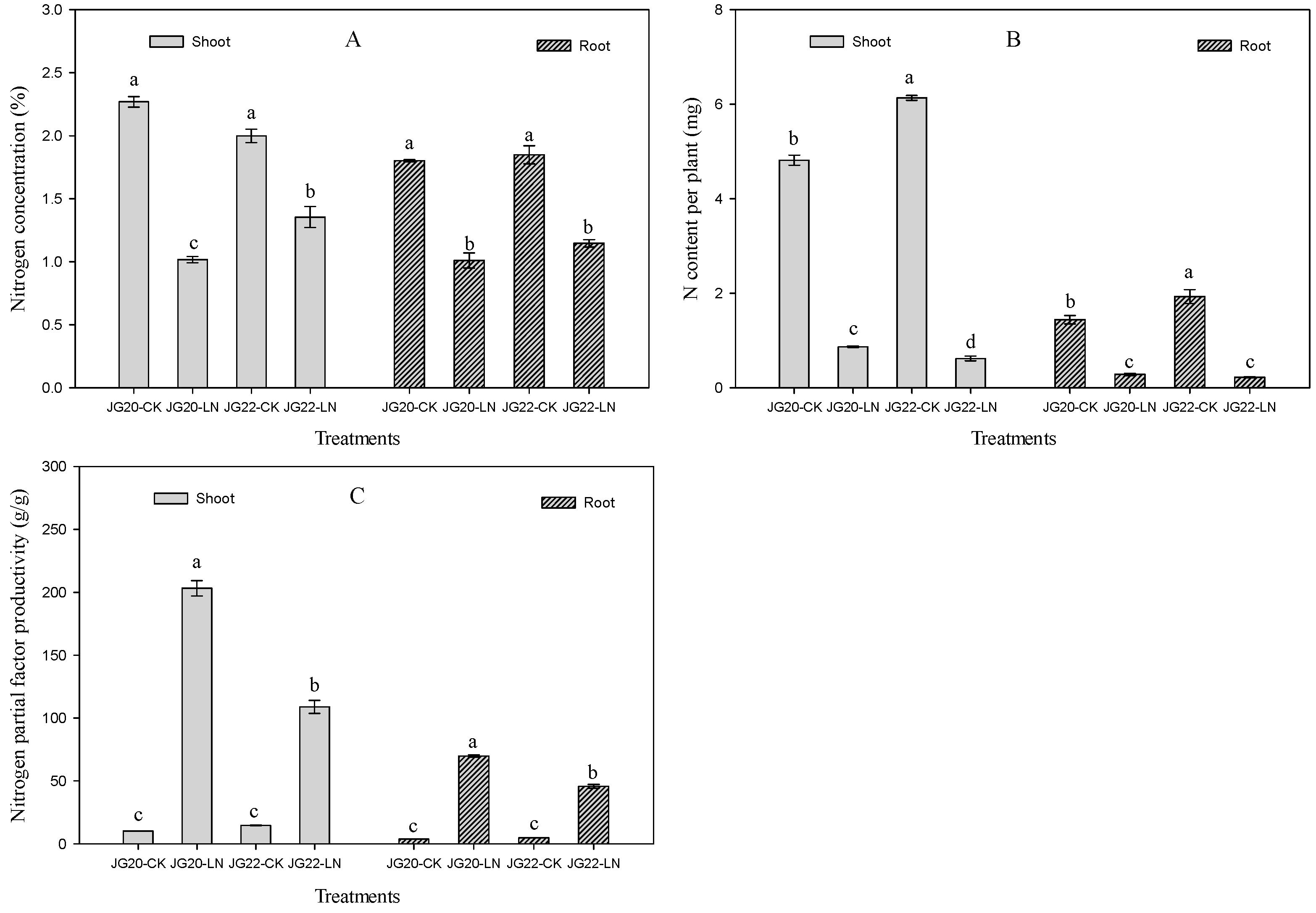
2.1. Phenotypic Characteristics and N Accumulation of Foxtail Millet
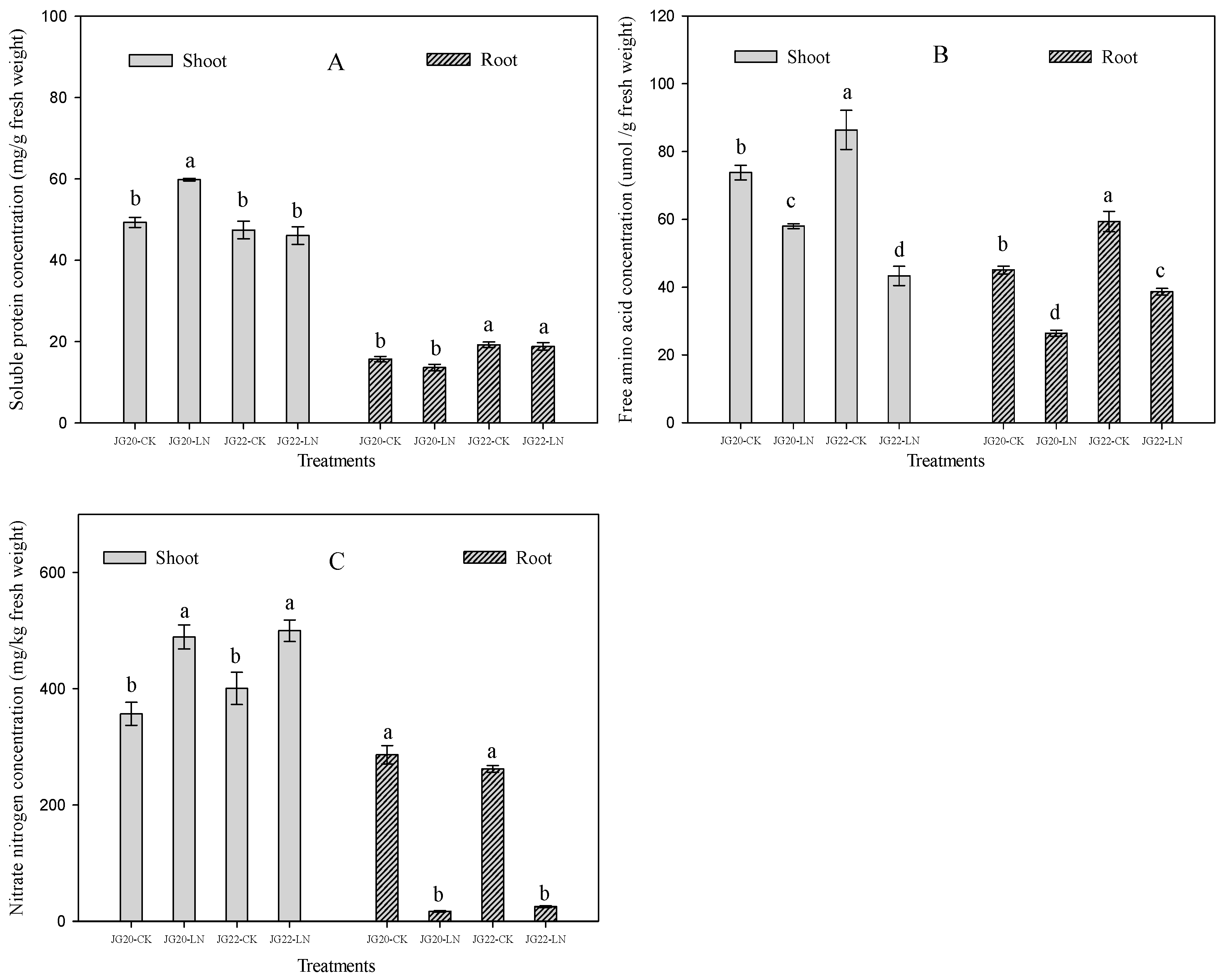
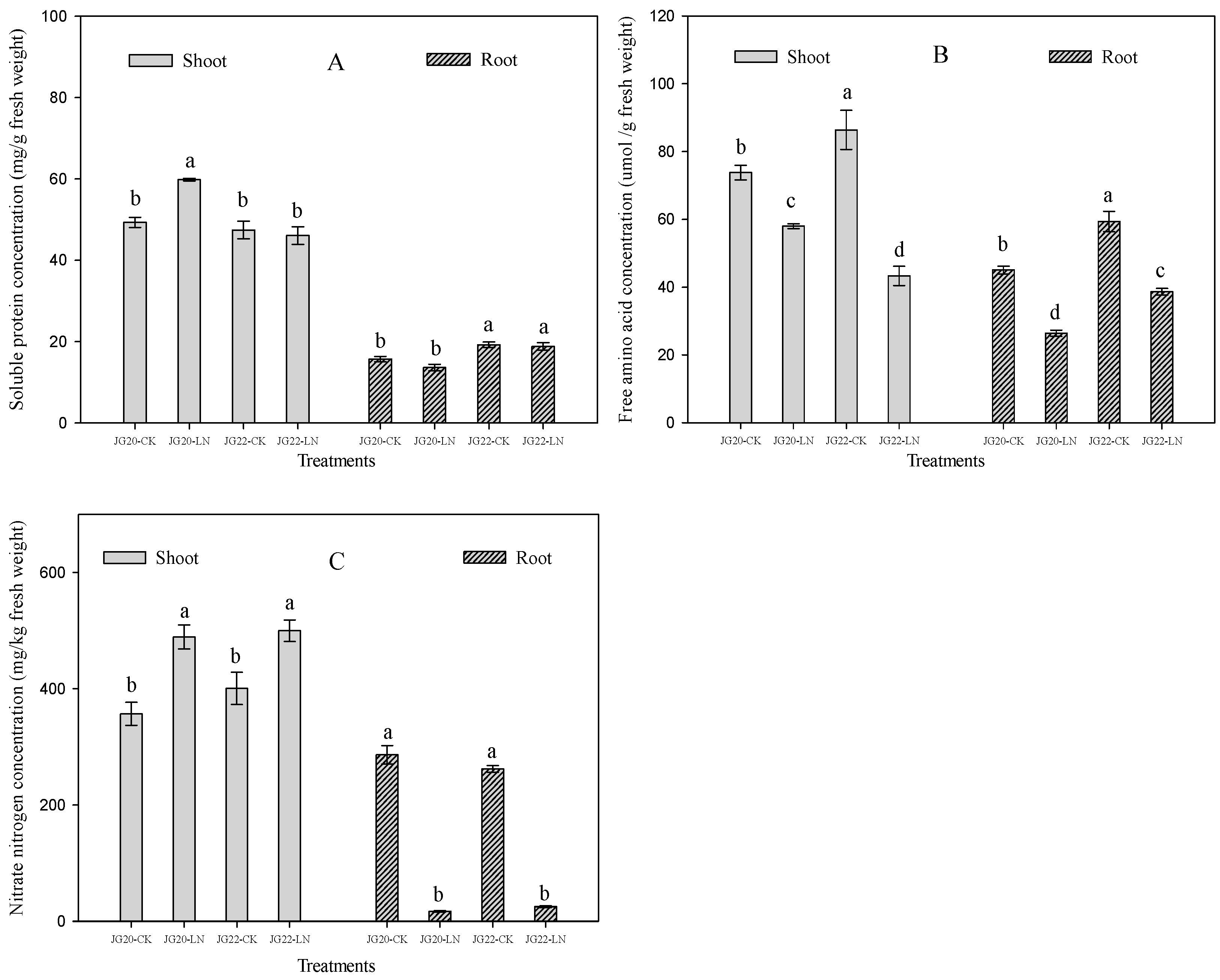
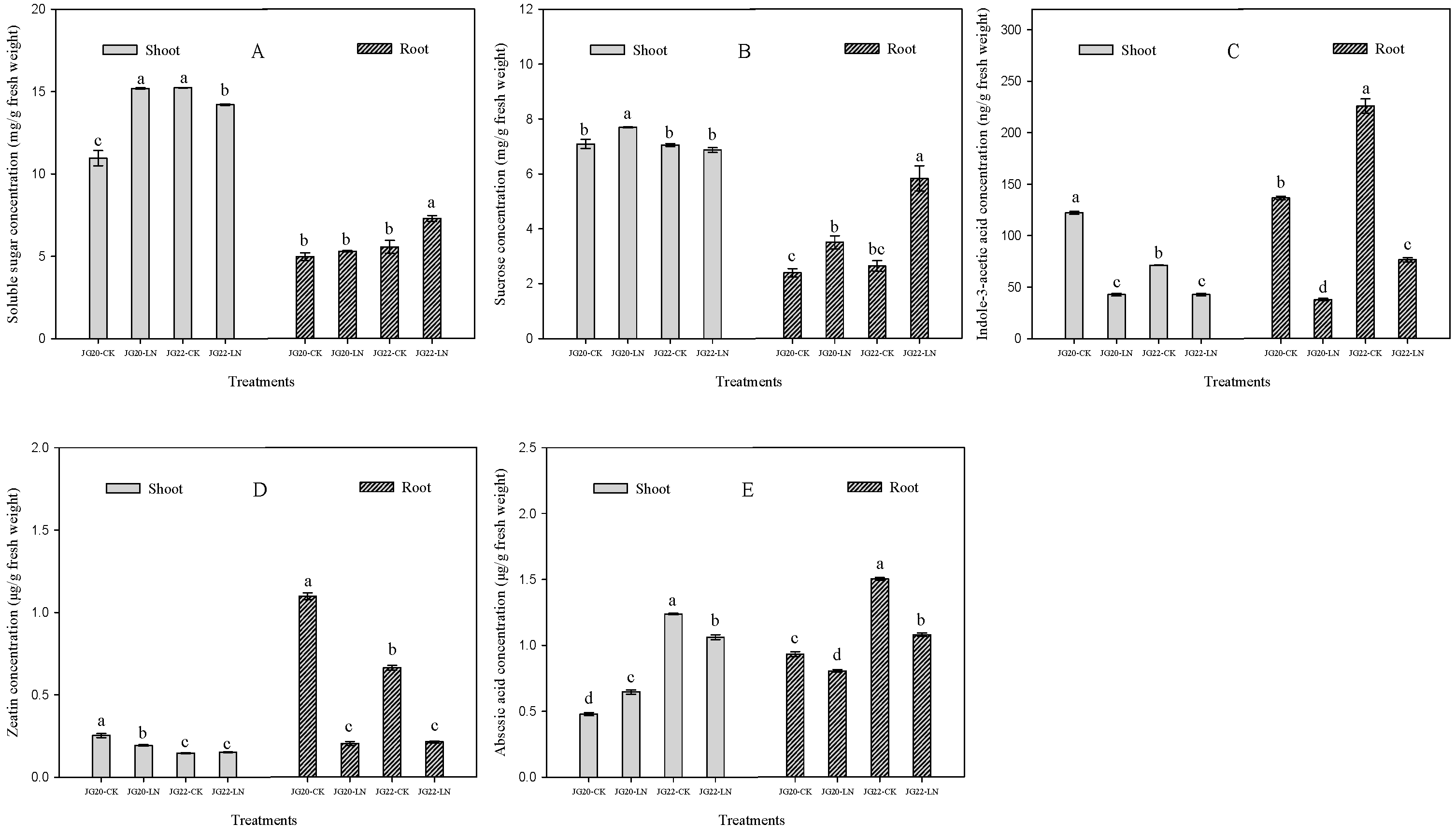
2.2. Physiological Characteristics and Response of Foxtail Millet to Low Nitrogen
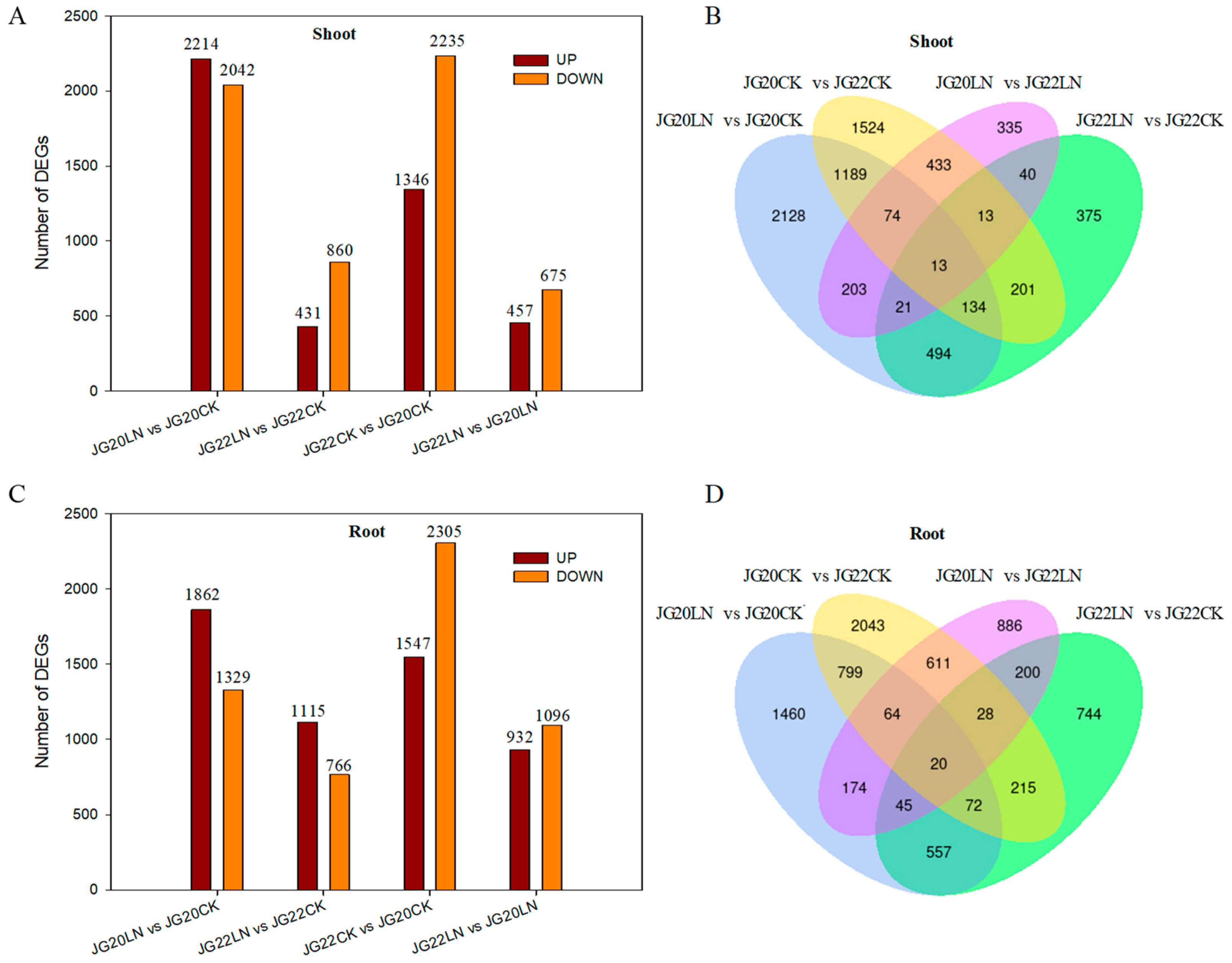
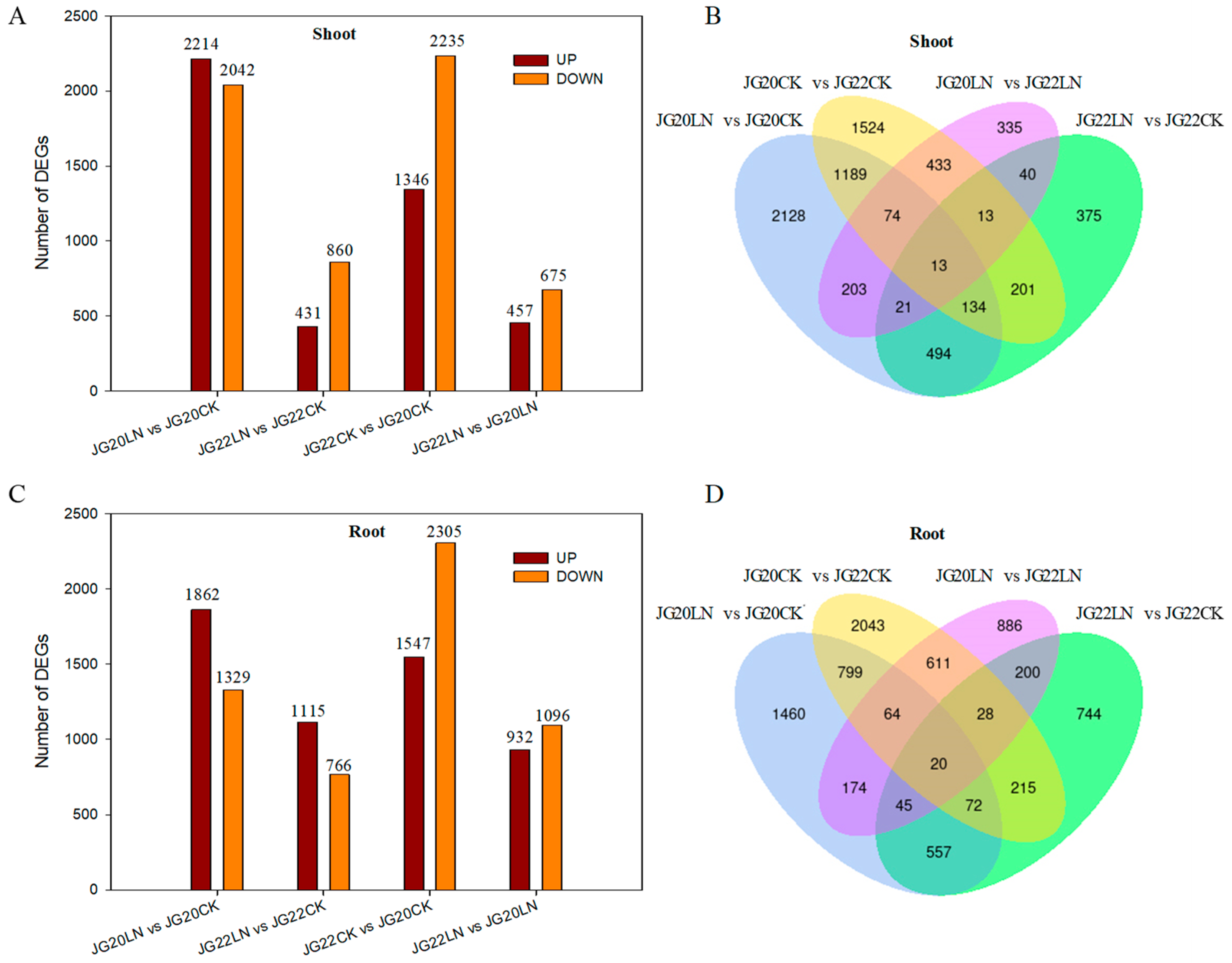
2.3. Differentially Expressed Genes (DEGs) between JG20 and JG22 under Low Nitrogen
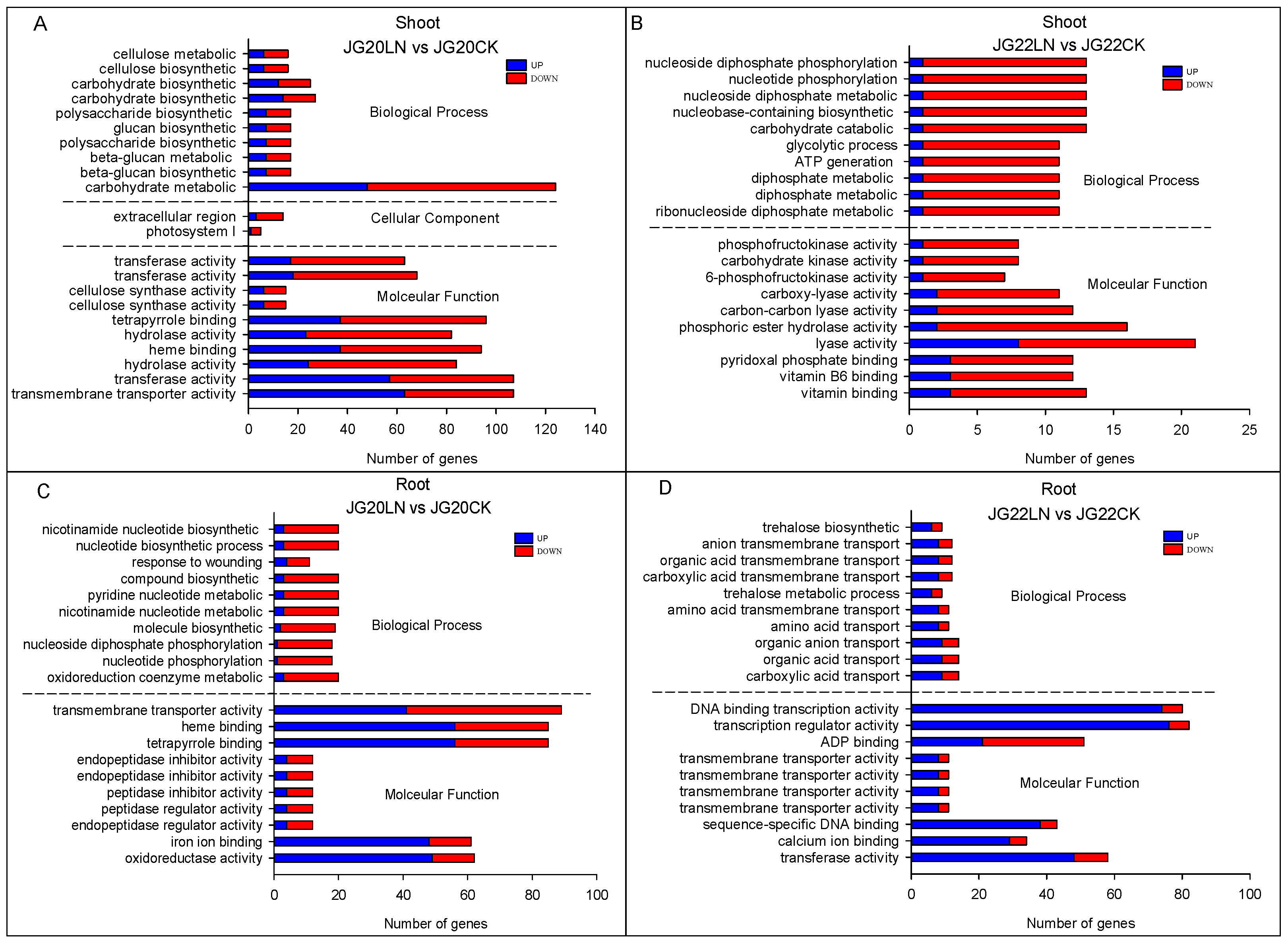
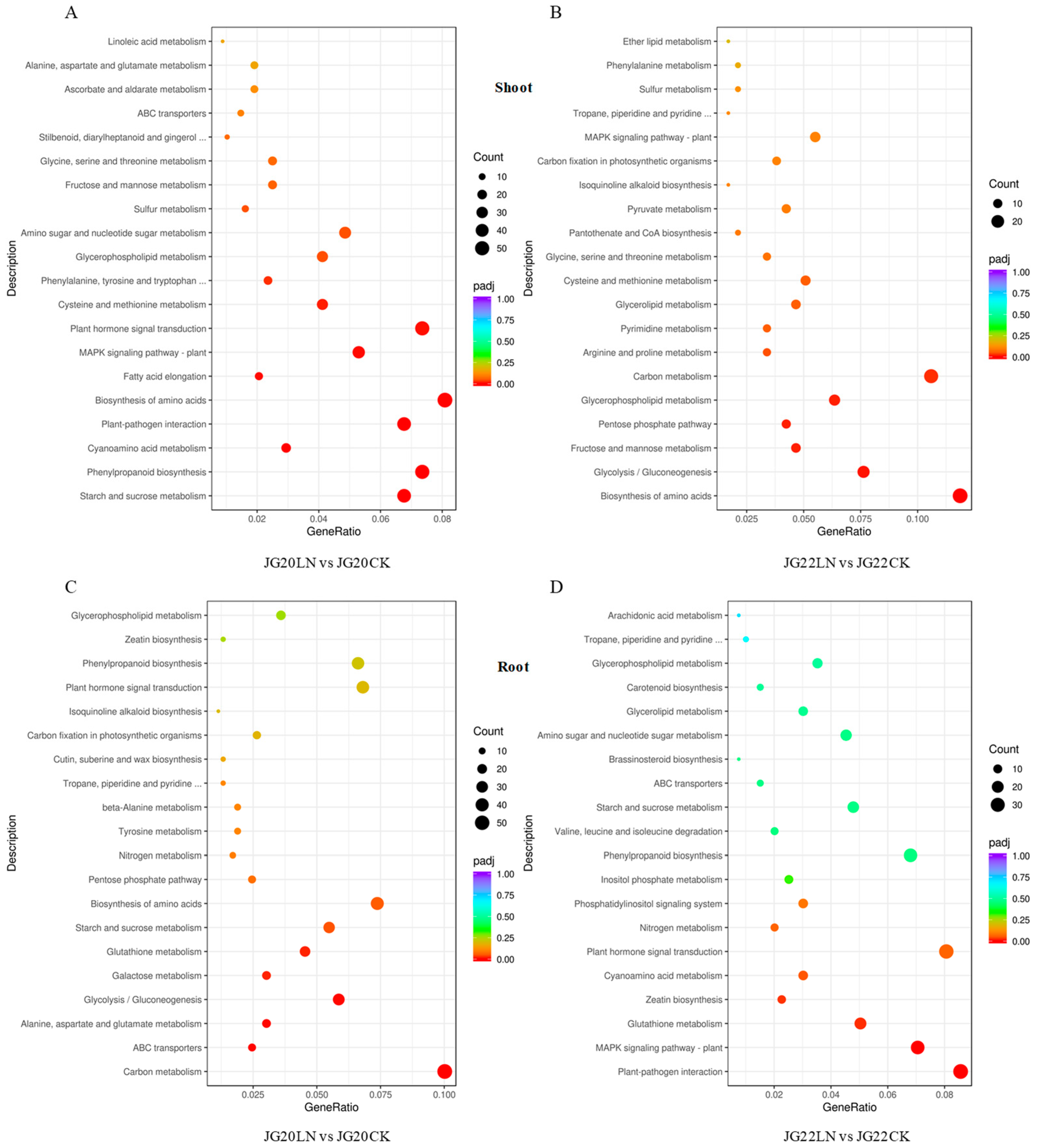
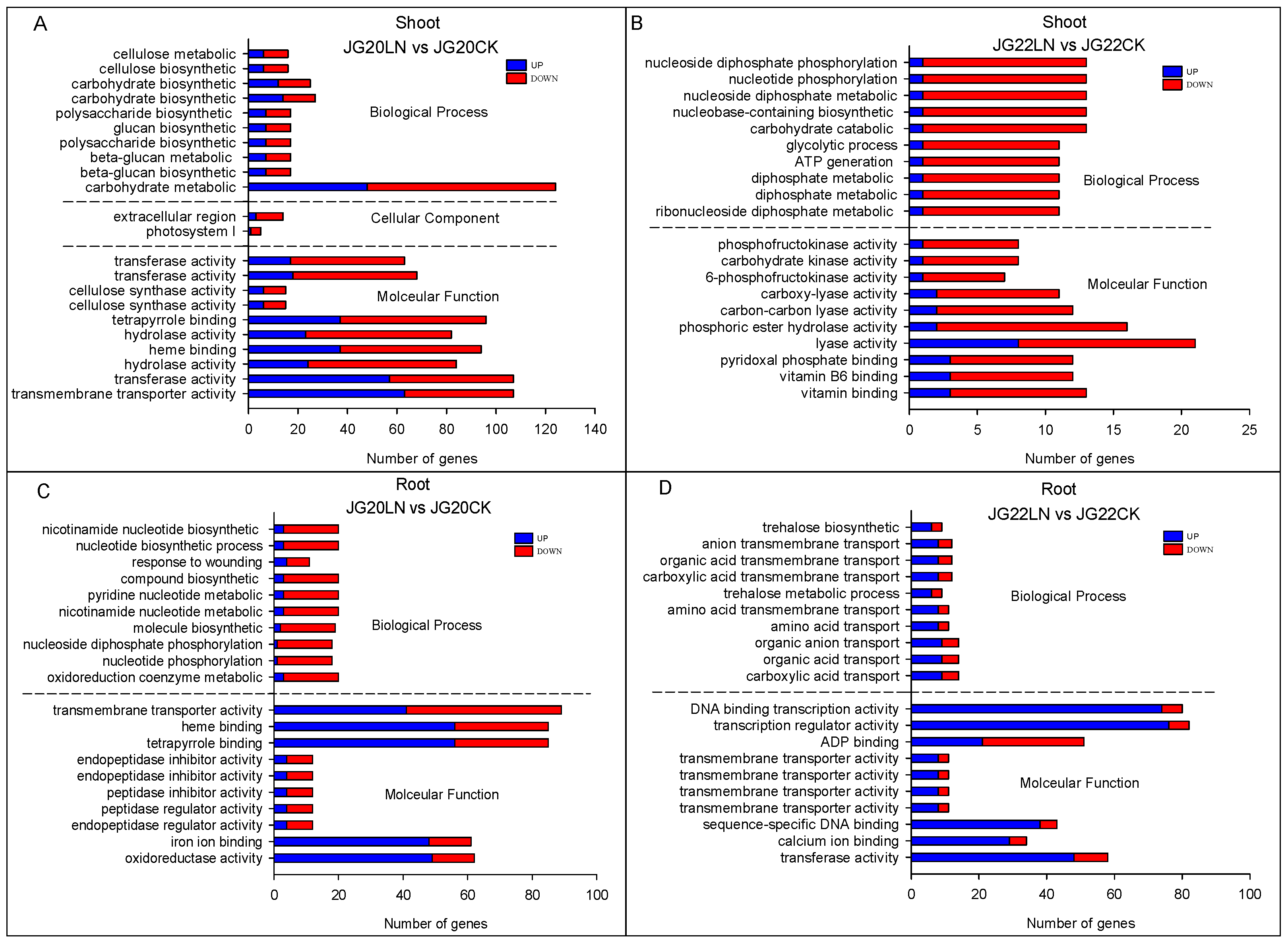
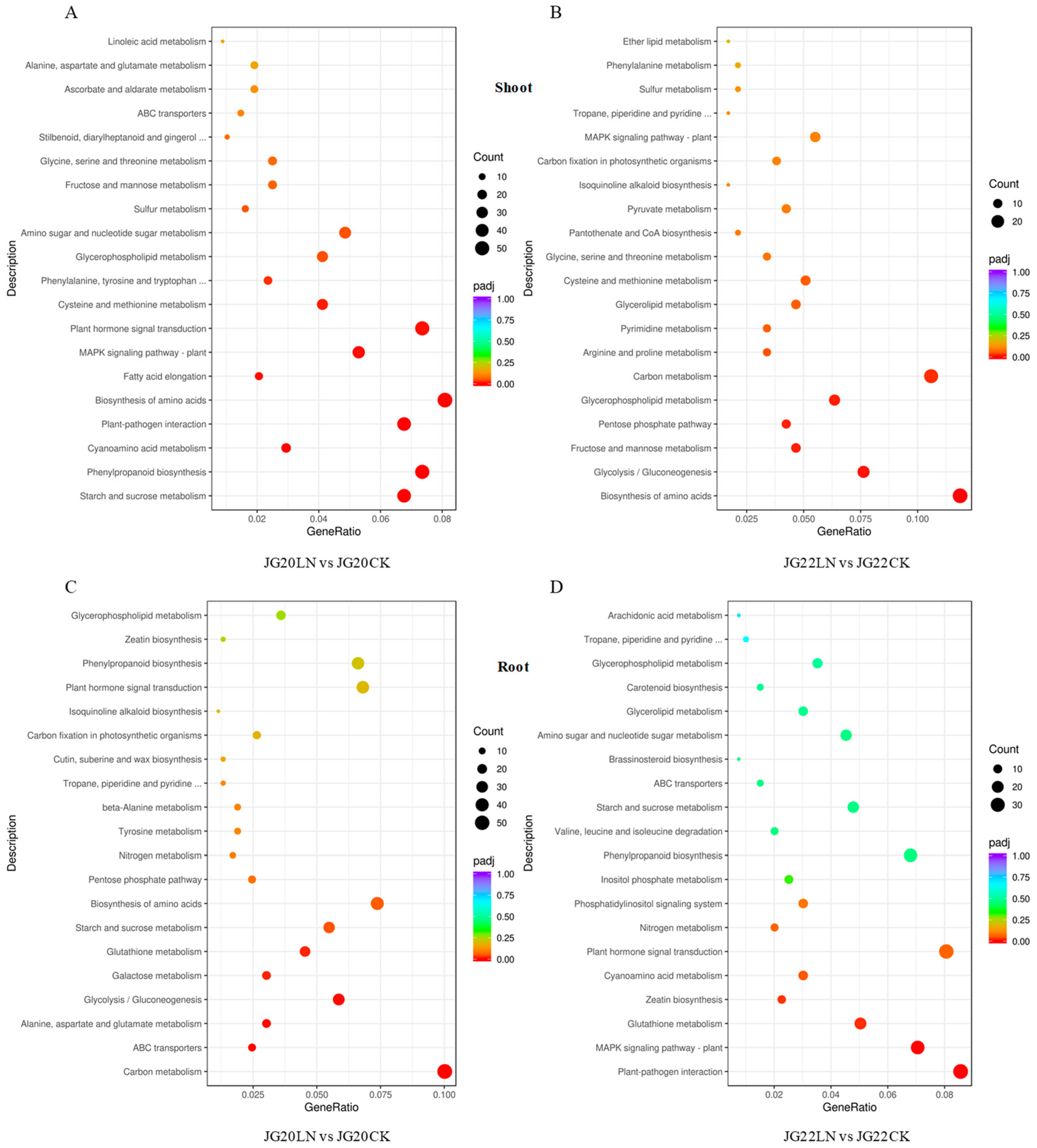
2.4. Functional Categorization of DEGs in Response to Low Nitrogen in Two Foxtail Millet Varieties
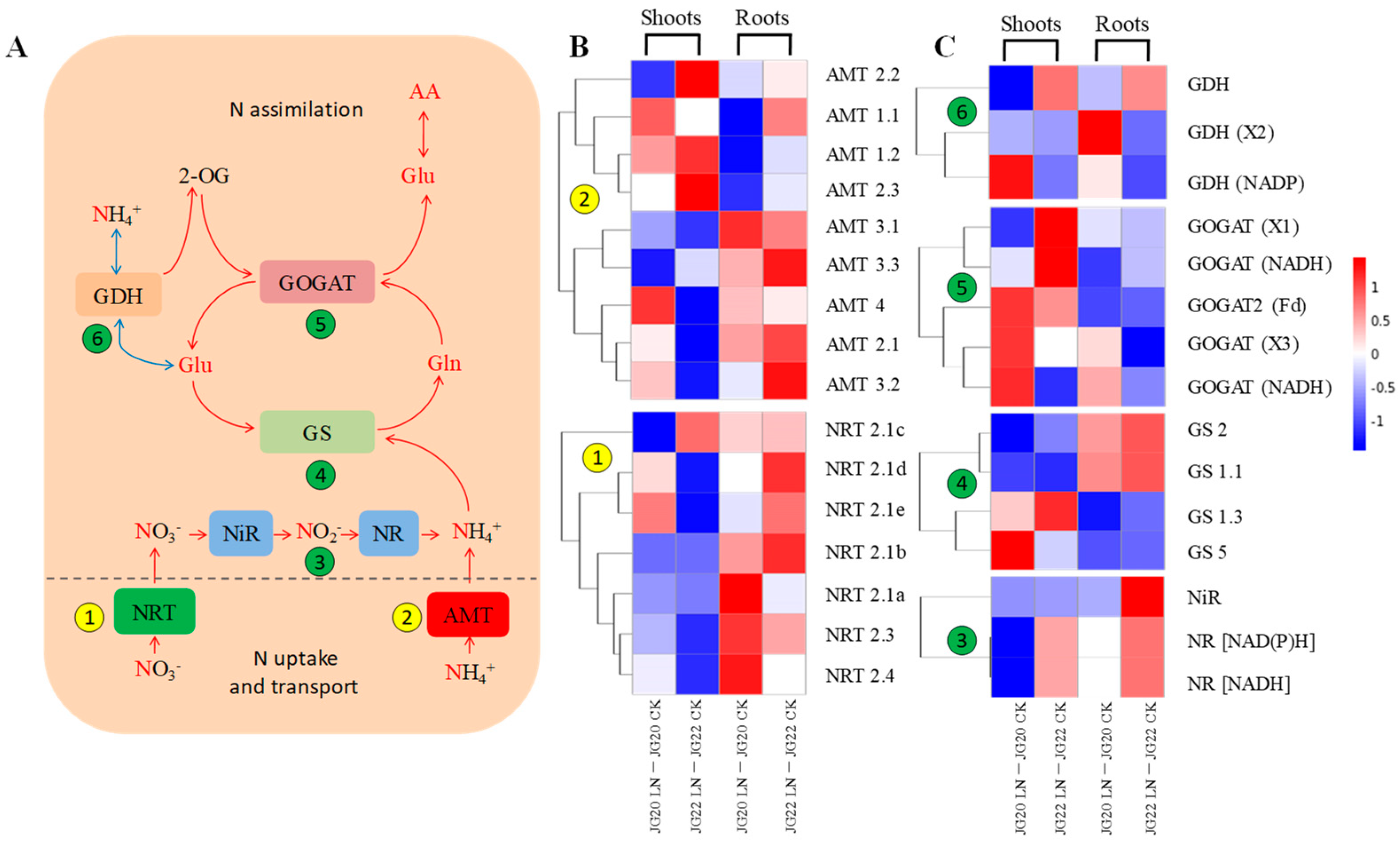
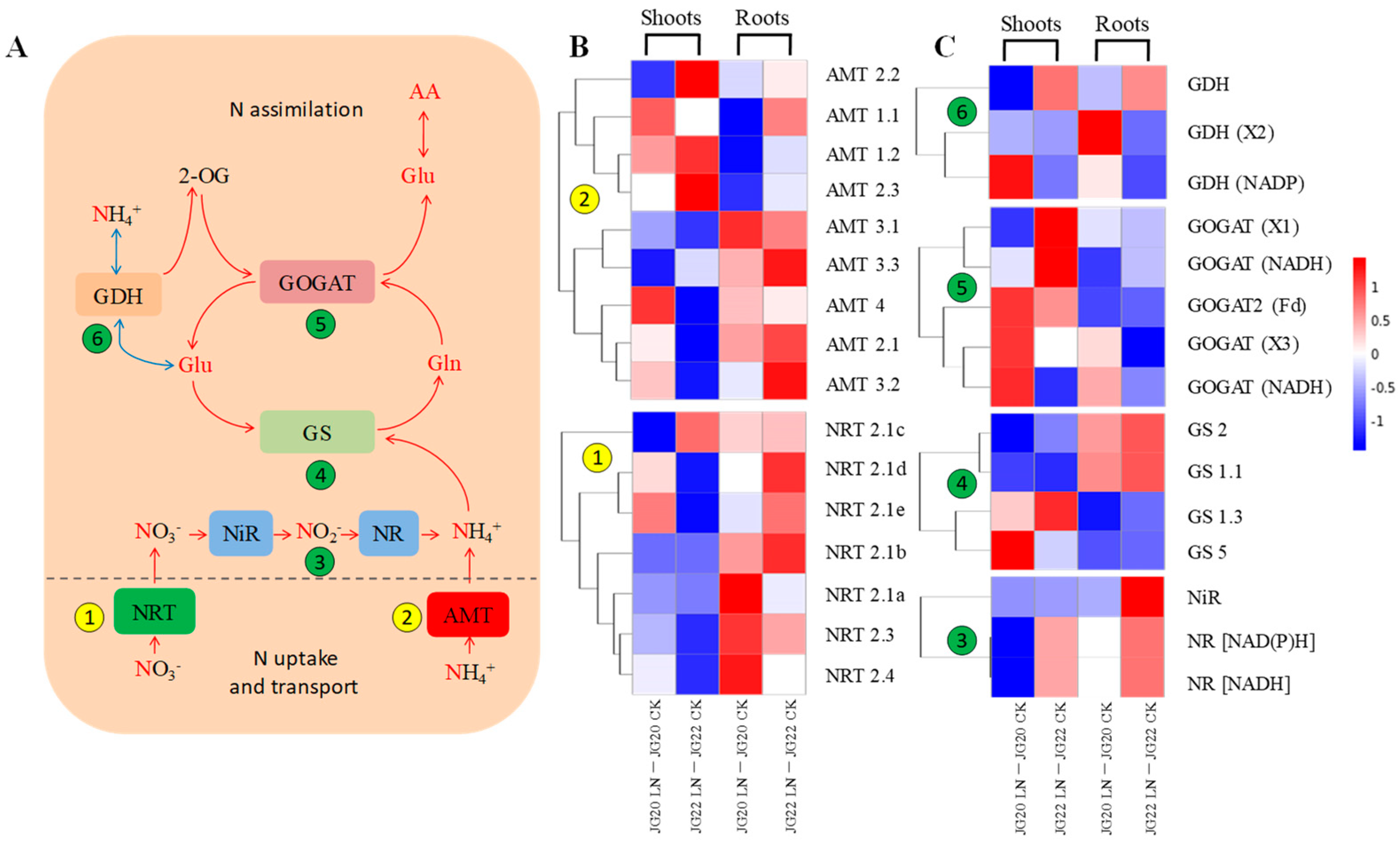
2.5. Gene Expression Profiling of Genes Involved in N Source Transport and Assimilation in Response to Low Nitrogen
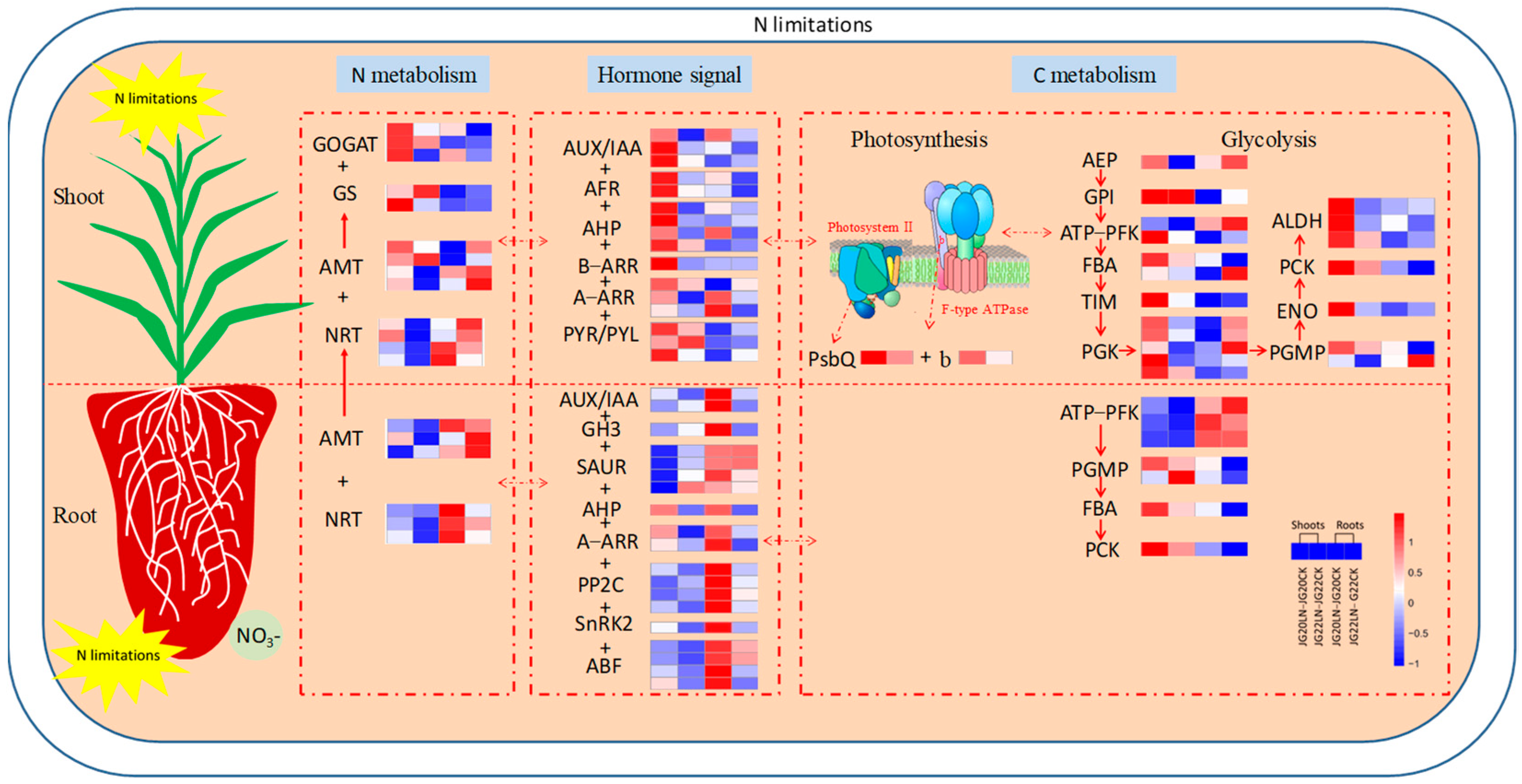
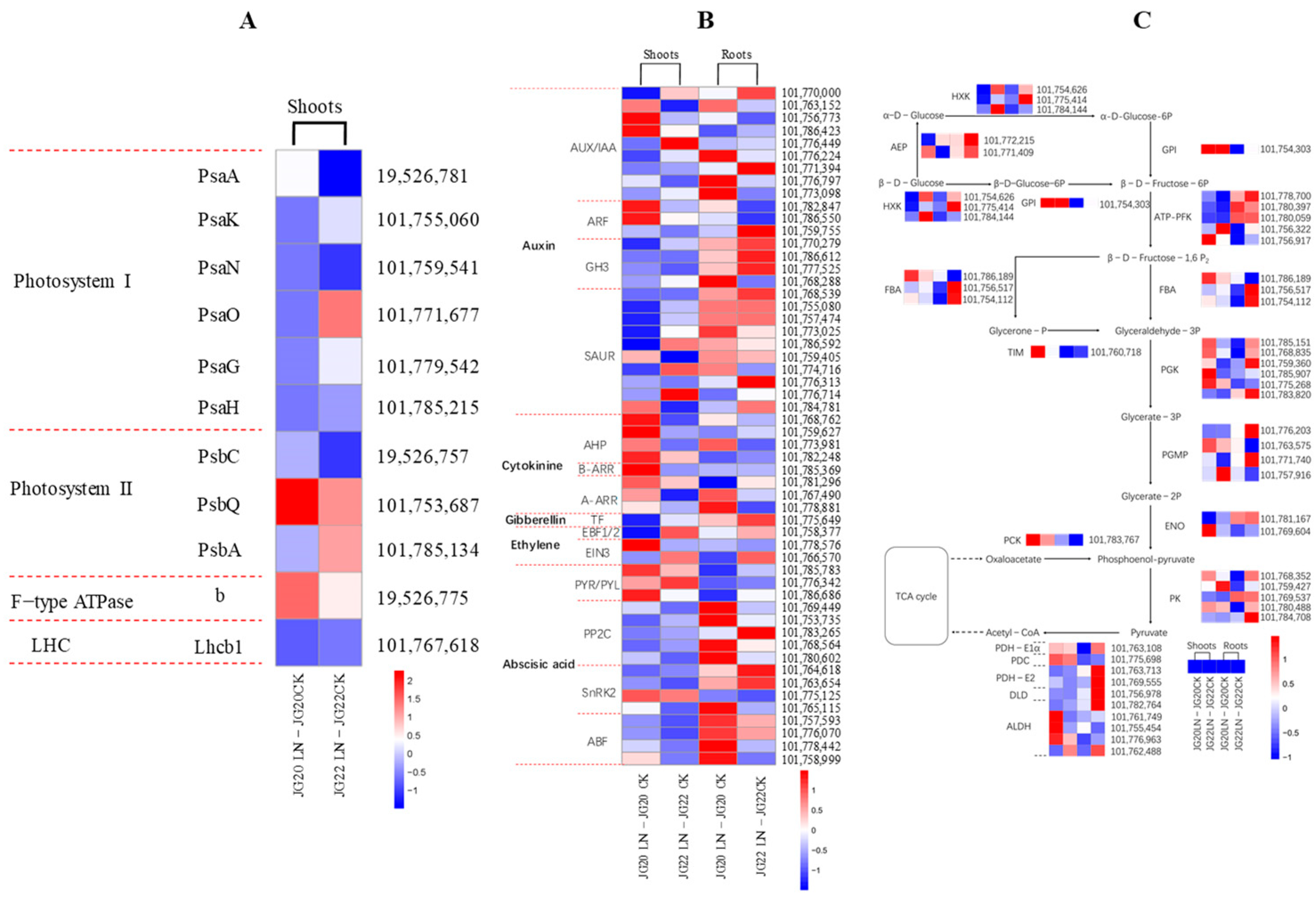
2.6. Gene Expression Profiling of Genes Involved in Photosynthesis, Hormone Signal Transduction, and Glycolysis in Response to Low Nitrogen
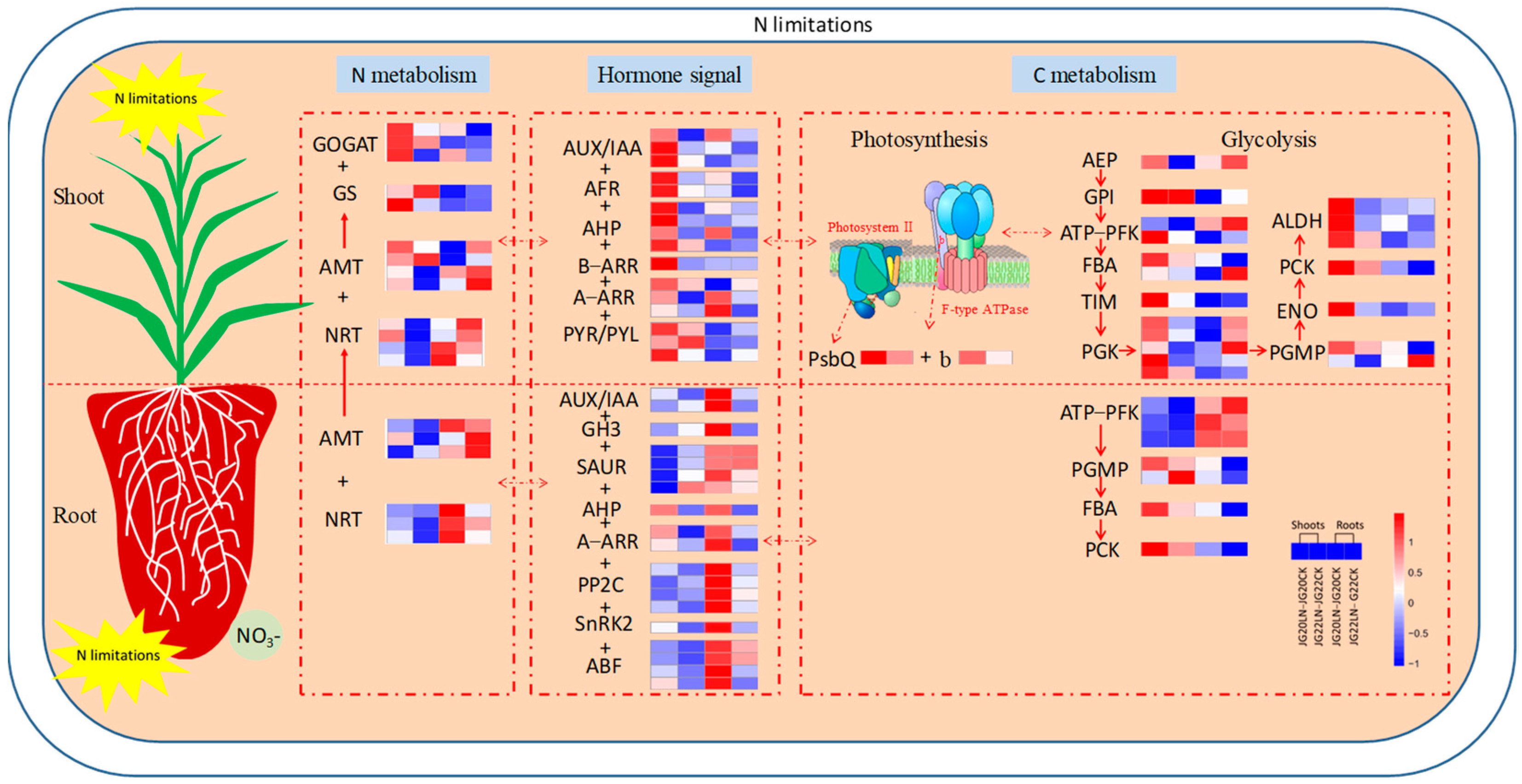
2.7. Schematic Representation of the Main Processes and Genes Involved in Foxtail Millet in Response to Low Nitrogen
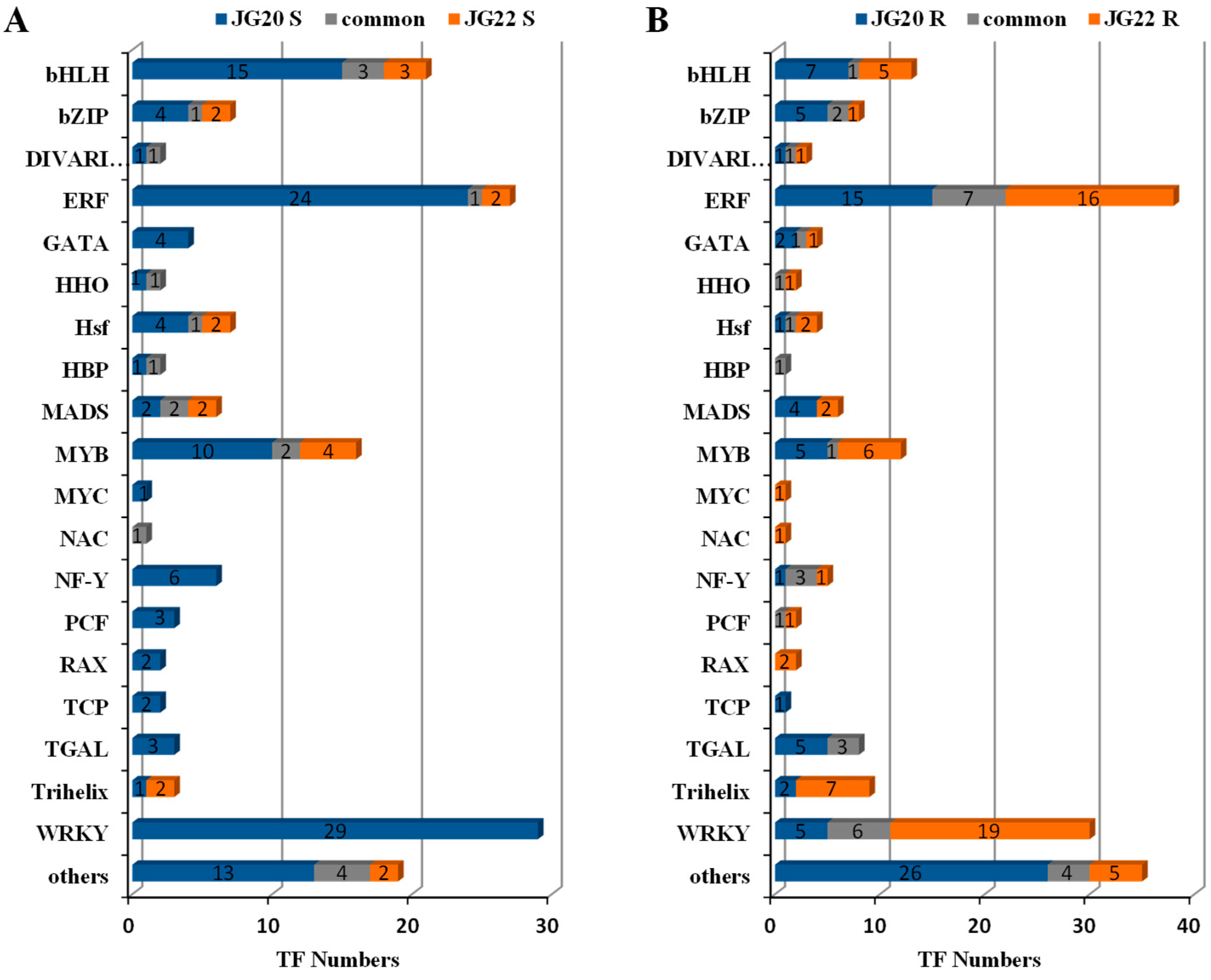
2.8. Expression Inventories of LN-Responsive TFs in Different Foxtail Millet Varieties
3. Discussion
4. Materials and Methods
4.1. Materials and Experimental Design
4.2. Measurement of NUE
4.3. Physiological Measurements
4.4. cDNA Library Contruction and Sequencing for RNA-Seq
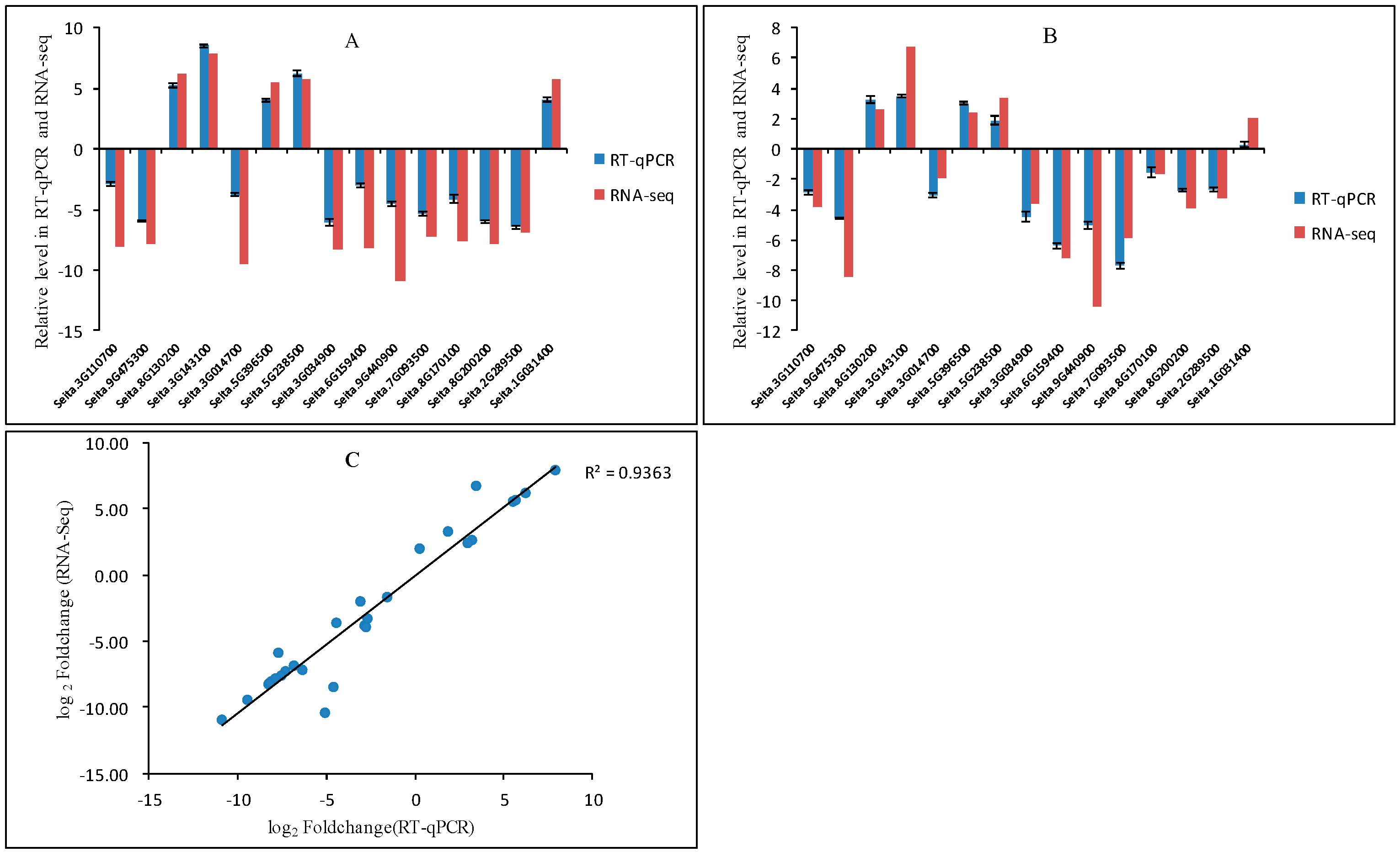
4.5. Validation of DEGs Using qRT-PCR
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weber, K.; Burow, M. Nitrogen—Essential macronutrient and signal controlling flowering time. Physiol. Plant 2018, 162, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hu, B.; Chu, C. Nitrogen use efficiency in crops: Lessons from Arabidopsis and rice. J. Exp. Bot. 2017, 68, 2477–2488. [Google Scholar] [CrossRef] [PubMed]
- Meharg, A. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA, 2011; p. 684. [Google Scholar]
- Xu, G.; Fan, X.; Miller, A.J. Plant Nitrogen Assimilation and Use Efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed]
- Vidal, E.; Alvarez, J.; Araus, V.; Riveras, E.; Brooks, M.; Krouk, G.; Ruffel, S.; Lejay, L.; Crawford, N.; Coruzzi, G.; et al. Nitrate 2020: Thirty years from transport to signaling networks. Plant Cell. 2020, 32, 2094–2119. [Google Scholar] [CrossRef]
- Sebilo, M.; Mayer, B.; Nicolardot, B.; Pinay, G.; Mariotti, A. Long-term fate of nitrate fertilizer in agricultural soils. Proc. Natl. Acad. Sci. USA 2013, 110, 18185–18189. [Google Scholar] [CrossRef]
- Zhu, Q.; de Vries, W.; Liu, X.; Hao, T.; Zeng, M.; Shen, J.; Zhang, F. Enhanced acidification in Chinese croplands as derived from element budgets in the period 1980–2010. Sci. Total Environ. 2018, 618, 1497–1505. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, X.; Vitousek, P. Chinese agriculture: An experiment for the world. Nature 2013, 497, 33–35. [Google Scholar] [CrossRef]
- Van der Straeten, B.; Buysse, J.; Nolte, S.; Lauwers, L.; Claeys, D.; Van Huylenbroeck, G. The effect of EU derogation strategies on the compliance costs of the nitrate directive. Sci. Total Environ. 2012, 421–422, 94–101. [Google Scholar] [CrossRef]
- Velthof, G.L.; Lesschen, J.P.; Webb, J.; Pietrzak, S.; Miatkowski, Z.; Pinto, M.; Kros, J.; Oenema, O. The impact of the Nitrates Directive on nitrogen emissions from agriculture in the EU-27 during 2000–2008. Sci. Total Environ. 2014, 468–469, 1225–1233. [Google Scholar] [CrossRef]
- Zhang, X.; Li, F.; Ding, Y.; Ma, Q.; Yi, Y.; Zhu, M.; Ding, J.; Li, C.; Guo, W.; Zhu, X. Transcriptome Analysis of Two Near-Isogenic Lines with Different NUE under Normal Nitrogen Conditions in Wheat. Biology 2021, 10, 1126. [Google Scholar] [CrossRef]
- Han, M.; Okamoto, M.; Beatty, P.H.; Rothstein, S.J.; Good, A.G. The Genetics of Nitrogen Use Efficiency in Crop Plants. Annu. Rev. Genet. 2015, 49, 269–289. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Ding, G.; Li, L.; Cai, H.; Ye, X.; Zou, J.; Xu, F. Identification and characterization of improved nitrogen efficiency in interspecific hybridized new-type Brassica napus. Ann. Bot. 2014, 114, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Wang, W.; Ou, S.; Tang, J.; Li, H.; Che, R.; Zhang, Z.; Chai, X.; Wang, H.; Wang, Y.; et al. Variation in NRT1.1B contributes to nitrate-use divergence between rice subspecies. Nat. Genet. 2015, 47, 834–838. [Google Scholar]
- Stahl, A.; Vollrath, P.; Samans, B.; Frisch, M.; Wittkop, B.; Snowdon, R.J. Effect of breeding on nitrogen use efficiency-associated traits in oilseed rape. J. Exp. Bot. 2019, 70, 1969–1986. [Google Scholar] [CrossRef] [PubMed]
- Bingham, I.J.; Karley, A.J.; White, P.J.; Thomas, W.T.B.; Russell, J.R. Analysis of improvements in nitrogen use efficiency associated with 75 years of spring barley breeding. Eur. J. Agron. 2012, 42, 49–58. [Google Scholar] [CrossRef]
- Palenchar, P.M.; Kouranov, A.; Lejay, L.V.; Coruzzi, G.M. Genome-wide patterns of carbon and nitrogen regulation of gene expression validate the combined carbon and nitrogen (CN)-signaling hypothesis in plants. Genome Biol. 2004, 5, R91. [Google Scholar] [CrossRef] [PubMed]
- Xuan, W.; Beeckman, T.; Xu, G. Plant nitrogen nutrition: Sensing and signaling. Curr. Opin. Plant Biol. 2017, 39, 57–65. [Google Scholar] [CrossRef]
- Baslam, M.; Mitsui, T.; Sueyoshi, K.; Ohyama, T. Recent Advances in Carbon and Nitrogen Metabolism in C3 Plants. Int. J. Mol. Sci. 2021, 22, 318. [Google Scholar] [CrossRef]
- Shen, T.; Xiong, Q.; Zhong, L.; Shi, X.; Cao, C.; He, H.; Chen, X. Analysis of main metabolisms during nitrogen deficiency and compensation in rice. Acta Physiol. Plant. 2019, 41, 68. [Google Scholar] [CrossRef]
- Krouk, G. Hormones and nitrate: A two-way connection. Plant Mol. Biol. 2016, 91, 599–606. [Google Scholar] [CrossRef]
- Yang, N.; Li, S.; Wang, S.; Li, Q.; Xu, F.; Shi, L.; Wang, C.; Ye, X.; Cai, H.; Ding, G. Dynamic transcriptome analysis indicates extensive and discrepant transcriptomic reprogramming of two rapeseed genotypes with contrasting NUE in response to nitrogen deficiency. Plant Soil. 2020, 456, 369–390. [Google Scholar] [CrossRef]
- Yang, X.; Wan, Z.; Perry, L.; Lu, H.; Wang, Q.; Zhao, C.; Li, J.; Xie, F.; Yu, J.; Cui, T.; et al. Early millet use in northern China. Proc. Natl. Acad. Sci. USA 2012, 109, 3726–3730. [Google Scholar] [CrossRef]
- Diao, X.; Schnable, J.; Bennetzen, J.L.; Li, J. Initiation of Setaria as a model plant. Front. Agric. Sci. Eng. 2014, 1, 16. [Google Scholar]
- Tang, S.; Li, L.; Wang, Y.; Chen, Q.; Zhang, W.; Jia, G.; Zhi, H.; Zhao, B.; Diao, X. Genotype-specific physiological and transcriptomic responses to drought stress in Setaria italica (an emerging model for Panicoideae grasses). Sci. Rep. 2017, 7, 10009. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, H.; Li, X.; Shen, H.; Gao, J.; Hou, S.; Zhang, B.; Mayes, S.; Bennett, M.; Ma, J.; et al. A mini foxtail millet with an Arabidopsis-like life cycle as a C4 model system. Nat. Plants 2020, 6, 1167–1178. [Google Scholar] [CrossRef]
- Nadeem, F.; Ahmad, Z.; Wang, R.; Han, J.; Shen, Q.; Chang, F.; Diao, X.; Zhang, F.; Li, X. Foxtail Millet [Setaria italica (L.) Beauv.] Grown under Low Nitrogen Shows a Smaller Root System, Enhanced Biomass Accumulation, and Nitrate Transporter Expression. Front. Plant Sci. 2018, 9, 205. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Qian, J.; Fang, Z.; Tang, J. Transcriptomic and physiological analyses of rice seedlings under different nitrogen supplies provide insight into the regulation involved in axillary bud outgrowth. BMC Plant Biol. 2020, 20, 197. [Google Scholar] [CrossRef] [PubMed]
- Quan, X.; Zeng, J.; Chen, G.; Zhang, G. Transcriptomic analysis reveals adaptive strategies to chronic low nitrogen in Tibetan wild barley. BMC Plant Biol. 2019, 19, 68. [Google Scholar] [CrossRef]
- Ge, L.; Dou, Y.; Li, M.; Qu, P.; He, Z.; Liu, Y.; Xu, Z.; Chen, J.; Chen, M.; Ma, Y. SiMYB3 in Foxtail Millet (Setaria italica) Confers Tolerance to Low-Nitrogen Stress by Regulating Root Growth in Transgenic Plants. Int. J. Mol. Sci. 2019, 20, 5741. [Google Scholar] [CrossRef]
- Erying, C.; Ling, Q.; Yanbing, Y.; Huawen, Z.; Hailian, W.; Bin, L.; Shuting, Y.; Runfeng, W.; Yanan, G. Variability of nitrogen use efficiency by foxtail millet cultivars at the seedling stage. Pesqui. Agropecu. Bras. 2020, 55, 00832. [Google Scholar] [CrossRef]
- Hawkesford, M.J.; Riche, A.B. Impacts of G x E x M on Nitrogen Use Efficiency in Wheat and Future Prospects. Front. Plant Sci. 2020, 11, 1157. [Google Scholar] [CrossRef]
- Chen, E.; Yang, Y.; Cheng, B.; Qin, L.; Zhang, H.; Liu, B.; Wang, H.; Guan, Y. Yield and nitrogen use efficiency in summer foxtail millet. Soil Fert. Sci. China 2015, 2, 93–97. [Google Scholar]
- Chun, L.; Mi, G.; Li, J.; Chen, F.; Zhang, F. Genetic Analysis of Maize Root Characteristics in Response to Low Nitrogen Stress. Plant Soil. 2005, 276, 369–382. [Google Scholar] [CrossRef]
- Tian, Q.; Chen, F.; Liu, J.; Zhang, F.; Mi, G. Inhibition of maize root growth by high nitrate supply is correlated with reduced IAA levels in roots. J. Plant Physiol. 2008, 165, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.U.N.; Chen, F.; Yuan, L.; Zhang, F.; Mi, G. A comprehensive analysis of root morphological changes and nitrogen allocation in maize in response to low nitrogen stress. Plant Cell Environ. 2015, 38, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ding, G.; Yang, Y.; White, P.; Ye, X.; Cai, H.; Lu, J.; Shi, L.; Xu, F. Comparative genome and transcriptome analysis unravel key factors of nitrogen use efficiency in Brassica napus L. Plant Cell Environ. 2020, 43, 712–731. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Han, J.; Wang, R.; Li, X. Down-regulation of nitrogen/carbon metabolism coupled with coordinative hormone modulation contributes to developmental inhibition of the maize ear under nitrogen limitation. Planta 2016, 244, 111–124. [Google Scholar] [CrossRef]
- Iqbal, A.; Dong, Q.; Wang, X.; Gui, H.; Zhang, H.; Zhang, X.; Song, M. High Nitrogen Enhance Drought Tolerance in Cotton through Antioxidant Enzymatic Activities, Nitrogen Metabolism and Osmotic Adjustment. Plants 2020, 9, 178. [Google Scholar] [CrossRef]
- Wolters, H.; Jürgens, G. Survival of the flexible: Hormonal growth control and adaptation in plant development. Nat. Rev. Genet. 2009, 10, 305–317. [Google Scholar] [CrossRef]
- Jones, B.; Gunnerås, S.A.; Petersson, S.V.; Tarkowski, P.; Graham, N.; May, S.; Dolezal, K.; Sandberg, G.; Ljung, K. Cytokinin Regulation of Auxin Synthesis in Arabidopsis Involves a Homeostatic Feedback Loop Regulated via Auxin and Cytokinin Signal Transduction. Plant Cell. 2010, 22, 2956–2969. [Google Scholar] [CrossRef]
- Sakakibara, H. CYTOKININS: Activity, Biosynthesis, and Translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and Signaling Aspects Underpinning the Regulation of Plant Carbon Nitrogen Interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef]
- Ruan, Y.-L. Sucrose Metabolism: Gateway to Diverse Carbon Use and Sugar Signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Ding, G.; Xu, F.; Cai, H.; Zou, J.; Ye, X. Genotype differences in photosynthetic characteristics and nitrogen efficiency of new-type oilseed rape responding to low nitrogen stress. J. Agric. Sci. 2015, 153, 1030–1043. [Google Scholar] [CrossRef]
- Mu, X.; Chen, Q.; Chen, F.; Yuan, L.; Mi, G. A RNA-Seq Analysis of the Response of Photosynthetic System to Low Nitrogen Supply in Maize Leaf. Int. J. Mol. Sci. 2017, 18, 2624. [Google Scholar] [CrossRef] [PubMed]
- Merchante, C.; Alonso, J.M.; Stepanova, A.N. Ethylene signaling: Simple ligand, complex regulation. Curr. Opin. Plant Biol. 2013, 16, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.-M.; Wang, R.-L.; Zhu, T.; Rothstein, S.J. Global transcription profiling reveals differential responses to chronic nitrogen stress and putative nitrogen regulatory components in Arabidopsis. BMC Genom. 2007, 8, 281. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Cheng, Y.-H.; Chen, K.-E.; Tsay, Y.-F. Nitrate Transport, Signaling, and Use Efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef]
- Cheng, J.; Tan, H.; Shan, M.; Duan, M.; Ye, L.; Yang, Y.; He, L.; Shen, H.; Yang, Z.; Wang, X. Genome-wide identification and characterization of the NPF genes provide new insight into low nitrogen tolerance in Setaria. Front. Plant Sci. 2022, 13, 1043832. [Google Scholar] [CrossRef]
- Dai, J.; Han, P.; Walk, T.C.; Yang, L.; Chen, L.; Li, Y.; Gu, C.; Liao, X.; Qin, L. Genome-Wide Identification and Characterization of Ammonium Transporter (AMT) Genes in Rapeseed (Brassica napus L.). Genes 2023, 14, 658. [Google Scholar] [CrossRef]
- Sinsirimongkol, K.; Buasong, A.; Teppabut, Y.; Pholmanee, N.; Chen, Y.; Miller, A.J.; Punyasuk, N. EgNRT2.3 and EgNAR2 expression are controlled by nitrogen deprivation and encode proteins that function as a two-component nitrate uptake system in oil palm. J. Plant Physiol. 2022, 279, 153833. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Bai, Z.-G.; Zhu, L.-F.; Zhang, J.-H.; Zhu, C.-Q.; Huang, J.-L.; Jin, Q.-Y.; Cao, X.-C. Nitrogen-mediated alleviation of photosynthetic inhibition under moderate water deficit stress in rice (Oryza sativa L.). Environ. Exp. Bot. 2019, 157, 269–282. [Google Scholar] [CrossRef]
- Zhou, W.; Yang, Z.; Wang, T.; Fu, Y.; Chen, Y.; Hu, B.; Yamagishi, J.; Ren, W. Environmental Compensation Effect and Synergistic Mechanism of Optimized Nitrogen Management Increasing Nitrogen Use Efficiency in Indica Hybrid Rice. Front. Plant Sci. 2019, 10, 245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiong, S.; Wei, Y.; Meng, X.; Wang, X.; Ma, X. The role of glutamine synthetase isozymes in enhancing nitrogen use efficiency of N-efficient winter wheat. Sci. Rep. 2017, 7, 1000. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Yang, S.; Wei, T.; Li, Y.; Wang, S.; Su, Y. Overexpression of OsGS1;2 for improved nitrogen use efficiency and grain yield of rice: A field test. Field Crops Res. 2023, 303, 109146. [Google Scholar] [CrossRef]
- Miflin, B.J.; Habash, D.Z. The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in the nitrogen utilization of crops. J. Exp. Bot. 2002, 53, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Ng, D.W.; Abeysinghe, J.K.; Kamali, M. Regulating the Regulators: The Control of Transcription Factors in Plant Defense Signaling. Int. J. Mol. Sci. 2018, 19, 3737. [Google Scholar] [CrossRef]
- Kong, L.; Zhang, Y.; Du, W.; Xia, H.; Fan, S.; Zhang, B. Signaling Responses to N Starvation: Focusing on Wheat and Filling the Putative Gaps with Findings Obtained in Other Plants. A Review. Front. Plant. Sci. 2021, 12, 656696. [Google Scholar] [CrossRef]
- Kant, S.; Bi, Y.-M.; Rothstein, S.J. Understanding plant response to nitrogen limitation for the improvement of crop nitrogen use efficiency. J. Exp. Bot. 2011, 62, 1499–1509. [Google Scholar] [CrossRef]
- Andrews, M.; Sprent, J.I.; Raven, J.A.; Eady, P.E. Relationships between shoot to root ratio, growth and leaf soluble protein concentration of Pisum sativum, Phaseolus vulgaris and Triticum aestivum under different nutrient deficiencies. Plant Cell Environ. 2010, 22, 949–958. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid Colorimetric Determination of Nitrate in Plant-Tissue by Nitration of Salicylic-Acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Luo, Y.; Li, W.; Huang, C.; Yang, J.; Jin, M.; Chen, J.; Pang, D.; Chang, Y.; Li, Y.; Wang, Z. Exogenous abscisic acid coordinating leaf senescence and transport of assimilates into wheat grains under drought stress by regulating hormones homeostasis. Crop J. 2021, 9, 901–914. [Google Scholar] [CrossRef]
- Li, W.; Zhang, M.; Qiao, L.; Chen, Y.; Zhang, D.; Jing, X.; Gan, P.; Huang, Y.; Gao, J.; Liu, W.; et al. Characterization of wavy root 1, an agravitropism allele, reveals the functions of OsPIN2 in fine regulation of auxin transport and distribution and in ABA biosynthesis and response in rice (Oryza sativa L.). Crop J. 2022, 12, 4. [Google Scholar] [CrossRef]
- Qin, L.; Chen, E.; Li, F.; Yu, X.; Liu, Z.; Yang, Y.; Wang, R.; Zhang, H.; Wang, H.; Liu, B.; et al. Genome-Wide Gene Expression Profiles Analysis Reveal Novel Insights into Drought Stress in Foxtail Millet (Setaria italica L.). Int. J. Mol. Sci. 2020, 21, 8520. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods. 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Robinson, M.; Oshlack, A.; Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol 11: R25. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef]
- Yoav, B.; Daniel, Y. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Raw Reads | Clean Reads | Clean Bases | Error Rate (20%) | Q20 (%) | Q30 (%) | GC Content (%) | Total Mapped | Uniquely Mapped | Multiple Mapped |
|---|---|---|---|---|---|---|---|---|---|---|
| SJG20-CK | 45,434,210 | 44,021,351 | 6.60 G | 0.02 | 98.18 | 94.78 | 56.54 | 42,466,340 (96.47%) | 40,838,306 (92.77%) | 1,628,034 (3.70%) |
| RJG20-CK | 46,926,017 | 44,908,641 | 6.73 G | 0.02 | 98.32 | 95.10 | 53.21 | 38,087,242 (84.81%) | 37,391,122 (83.26) | 696,119 (1.55%) |
| SJG20-LN | 46,568,083 | 45,266,295 | 6.79 G | 0.02 | 98.15 | 94.69 | 56.37 | 43,654,878 (96.44%) | 42,879,943 (94.73%) | 774,935 (1.71%) |
| RJG20-LN | 45,728,815 | 44,559,130 | 6.68 G | 0.02 | 98.11 | 94.52 | 52.95 | 33,272,477 (74.67%) | 32,684,085 (73.35%) | 588,392 (1.32%) |
| SJG22-CK | 42,193,254 | 41,110,815 | 6.17 G | 0.02 | 98.36 | 95.08 | 53.98 | 39,846,088 (96.92%) | 38,835,094 (94.46%) | 1,010,994 (2.46%) |
| RJG22-CK | 43,185,923 | 42,196,855 | 6.33 G | 0.02 | 98.33 | 94.95 | 50.22 | 30,066,139 (71.25%) | 29,460,001 (69.82%) | 606,138 (1.44%) |
| SJG22-LN | 44,891,345 | 43,960,698 | 6.59 G | 0.02 | 98.13 | 94.55 | 54.73 | 42,392,060 (96.43%) | 41,541,056 (94.50%) | 851,004 (1.94%) |
| RJG22-LN | 43,616,102 | 42,607,394 | 6.39 G | 0.02 | 98.24 | 94.77 | 52.21 | 31,884,267 (74.83%) | 31,199,409 (73.23%) | 684,858 (1.61%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, E.; Qin, L.; Li, F.; Yang, Y.; Liu, Z.; Wang, R.; Yu, X.; Niu, J.; Zhang, H.; Wang, H.; et al. Physiological and Transcriptomic Analysis Provides Insights into Low Nitrogen Stress in Foxtail Millet (Setaria italica L.). Int. J. Mol. Sci. 2023, 24, 16321. https://doi.org/10.3390/ijms242216321
Chen E, Qin L, Li F, Yang Y, Liu Z, Wang R, Yu X, Niu J, Zhang H, Wang H, et al. Physiological and Transcriptomic Analysis Provides Insights into Low Nitrogen Stress in Foxtail Millet (Setaria italica L.). International Journal of Molecular Sciences. 2023; 24(22):16321. https://doi.org/10.3390/ijms242216321
Chicago/Turabian StyleChen, Erying, Ling Qin, Feifei Li, Yanbing Yang, Zhenyu Liu, Runfeng Wang, Xiao Yu, Jiahong Niu, Huawen Zhang, Hailian Wang, and et al. 2023. "Physiological and Transcriptomic Analysis Provides Insights into Low Nitrogen Stress in Foxtail Millet (Setaria italica L.)" International Journal of Molecular Sciences 24, no. 22: 16321. https://doi.org/10.3390/ijms242216321
APA StyleChen, E., Qin, L., Li, F., Yang, Y., Liu, Z., Wang, R., Yu, X., Niu, J., Zhang, H., Wang, H., Liu, B., & Guan, Y. (2023). Physiological and Transcriptomic Analysis Provides Insights into Low Nitrogen Stress in Foxtail Millet (Setaria italica L.). International Journal of Molecular Sciences, 24(22), 16321. https://doi.org/10.3390/ijms242216321