

Haberlea rhodopensis Extract Tunes the Cellular Response to Stress by Modulating DNA Damage, Redox Components, and Gene Expression

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
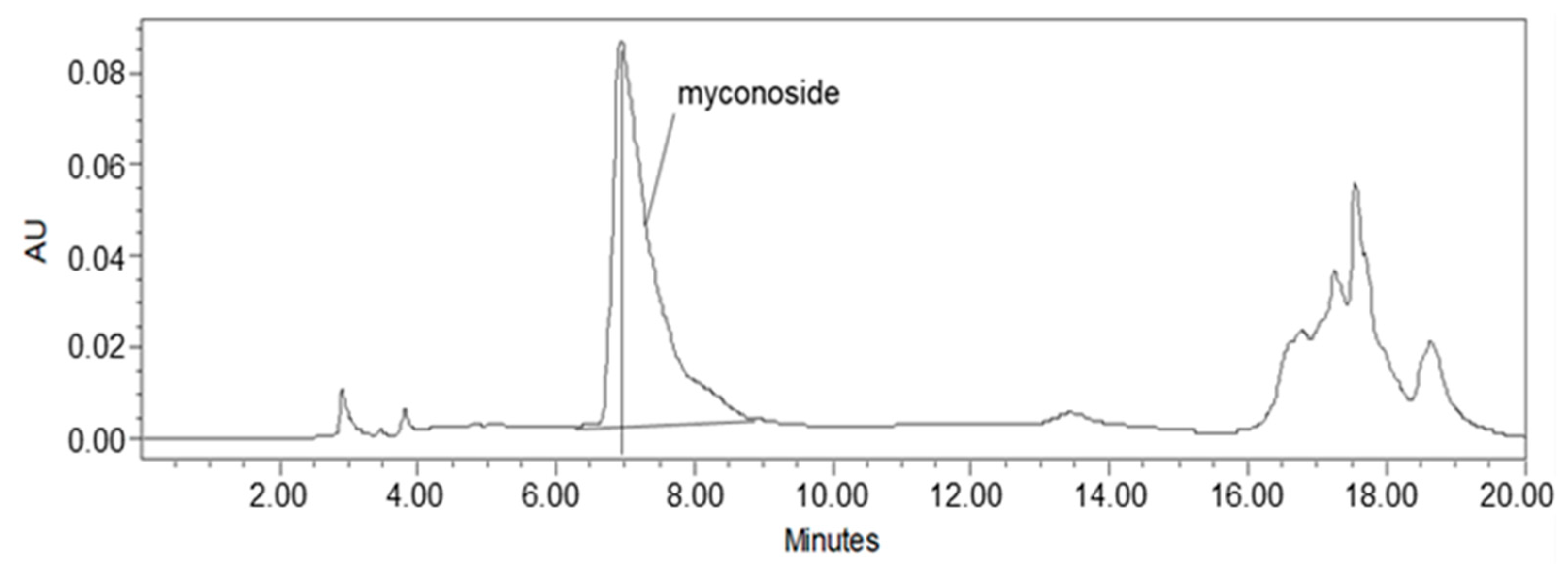
2.1. Myconoside Is the Dominant Compound in the Prepared Total Ethanolic Haberlea rhodopensis Leaves Extract
2.2. No Impact of HRE on Cellular Morphology and Cell Cycle Progression in HeLa Cells
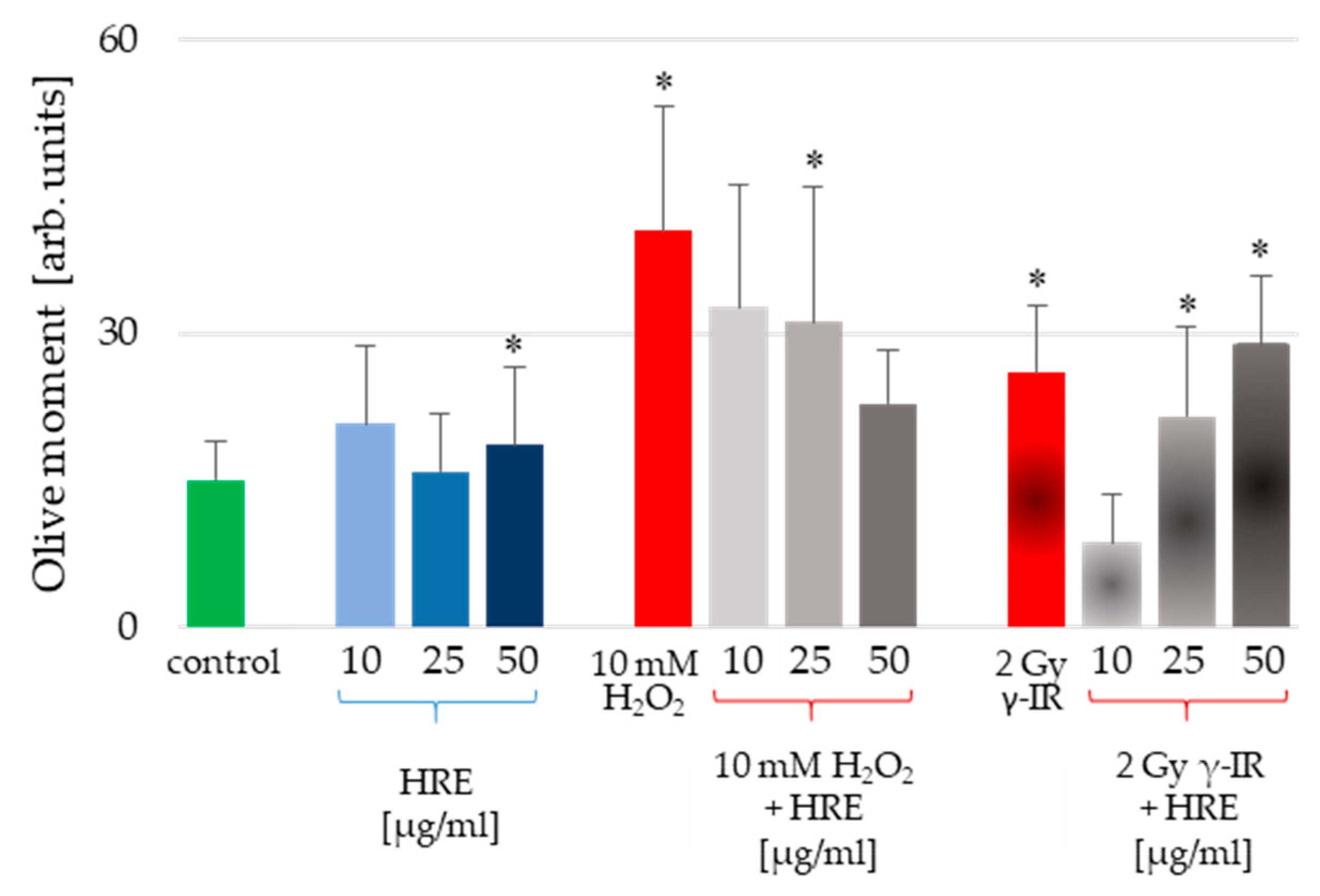
2.3. Exploring the Protective Potential of HRE Pretreatment against Genotoxicity in HeLa Cells
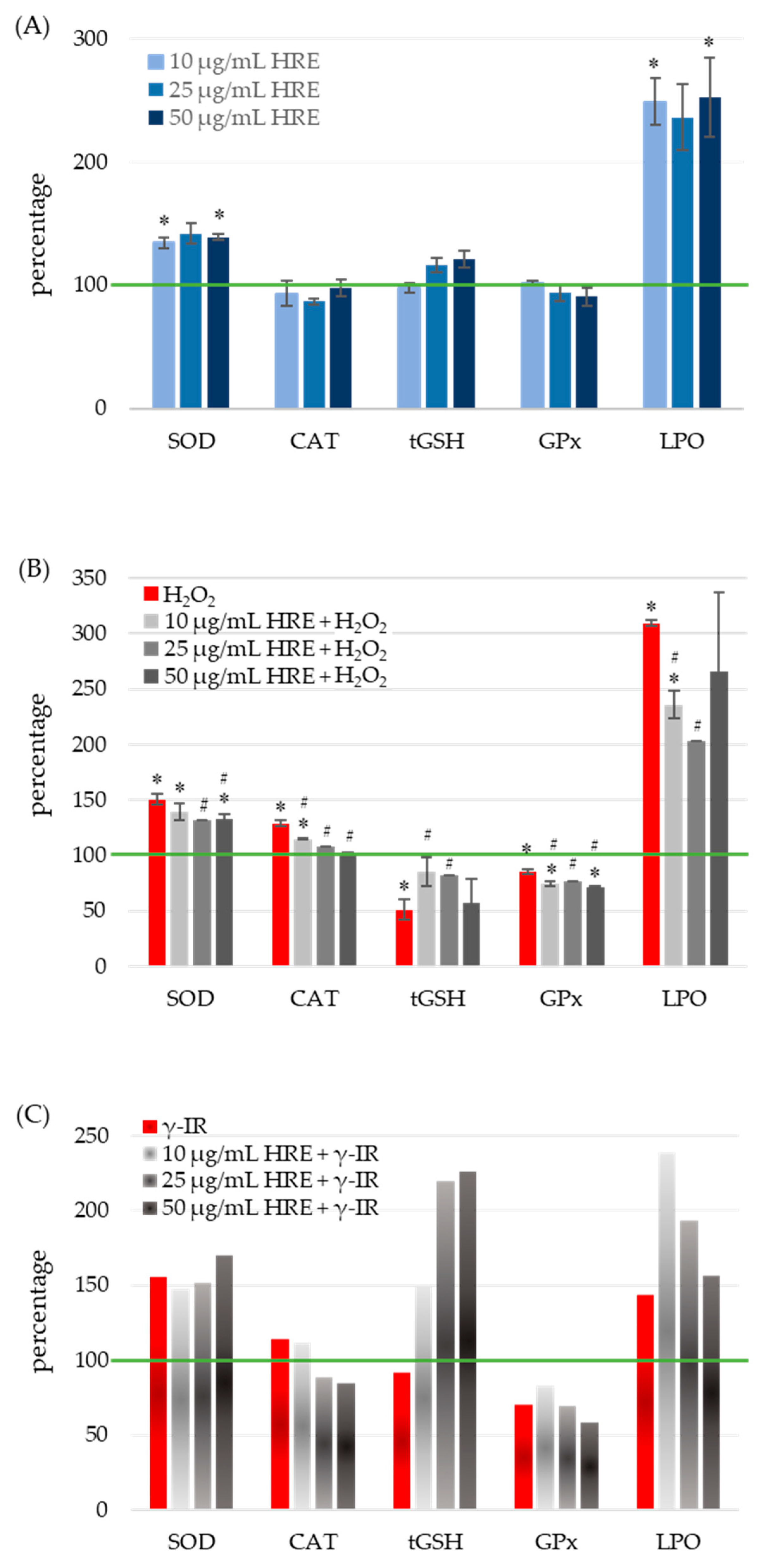
2.4. Exploring the Beneficial Effects of Pretreatment with HRE on Cellular Antioxidant Status: Protection against H2O2 and γ-IR-Induced Stress
2.4.1. Modulation of Superoxide Dismutase Activity by HRE and Genotoxic Stress
2.4.2. Impact of HRE Administration, Oxidative and Radiation Stress on Catalase Activity
2.4.3. Modulation of Glutathione Levels by HRE in Response to Oxidative and Radiation Stress
2.4.4. Modulation of Glutathione Peroxidase Activity by HRE in Response to Oxidative and Radiation Stress
2.4.5. Modulation of Lipid Peroxidation and Antioxidant Defense by HRE in HeLa Cells
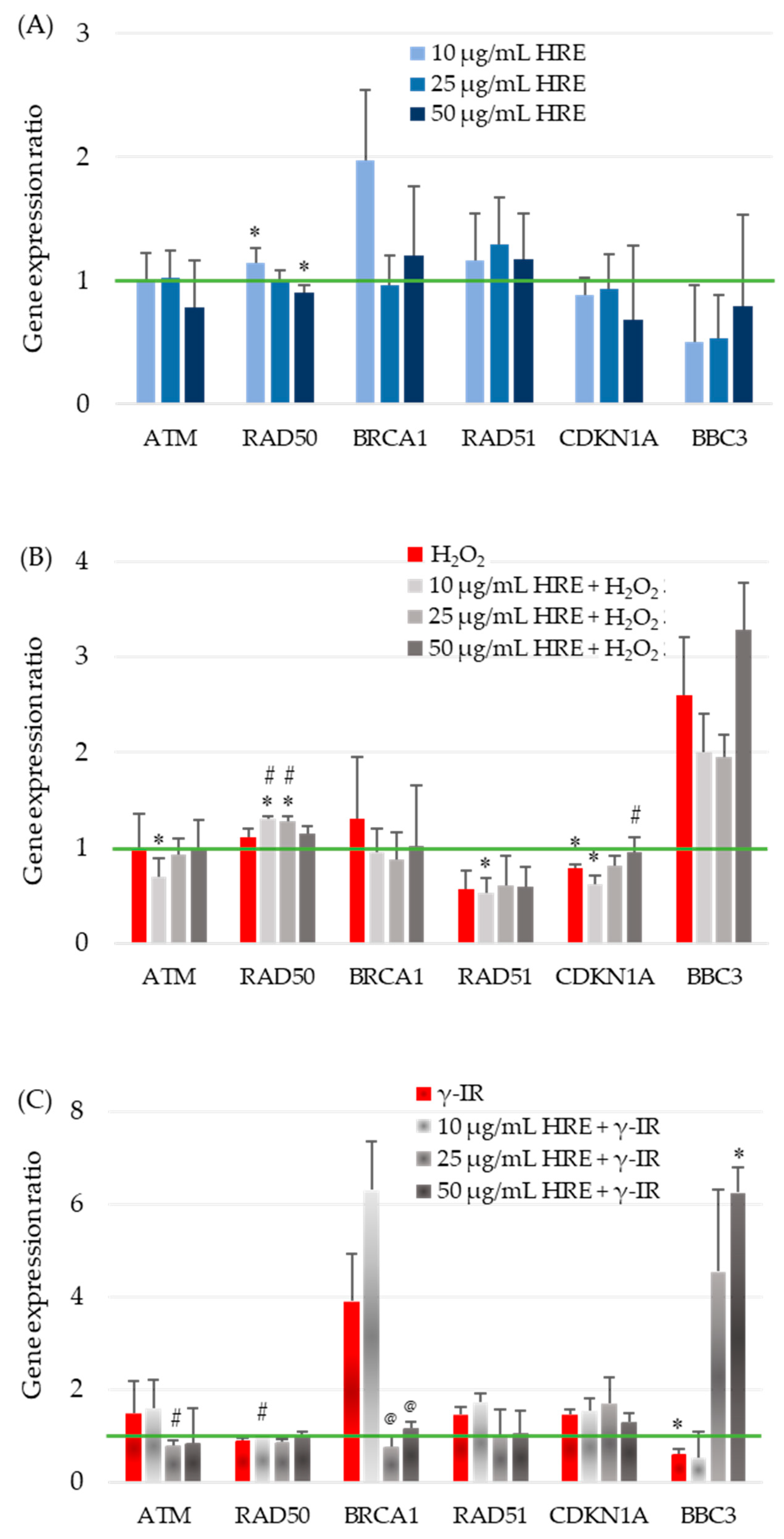
2.5. Influence of H. rhodopensis Extract on Gene Expression in Response to Oxidative Stress and Radiation
2.5.1. Evaluation of ATM Gene Expression
2.5.2. Influence of H. rhodopensis Extract on RAD50 Gene Expression in Response to Oxidative Stress and Radiation
2.5.3. Effects of HRE on BRCA1 Gene Expression and Radiosensitivity
2.5.4. Influence of Ethanolic HRE on RAD51 Gene Expression
2.5.5. Modulation of CDKN1A Gene Expression by Ethanolic HRE and Cellular Stressors
2.5.6. Expression of Human BBC3 Gene Coding for p53 Upregulated Modulator of Apoptosis (PUMA)
3. Discussion
3.1. HRE Protects HeLa Cells against Radiation and H2O2-Induced DNA Damage
3.2. Redox/Antioxidant Response to HRE and HRE Post-Applied Stress
3.3. Gene Transcription Response to Stress after HRE Pre-Incubation
3.3.1. Gene Activity upon HRE Administration Alone
3.3.2. Gene Activity upon Stressor Treatment
3.3.3. Gene Activity upon Combined Preincubation with HRE and Treatment with Stressors
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Haberlea rhodopensis Leaves Gathering and Plant Identification
4.3. Haberlea rhodopensis Extract Preparation
4.4. High Performance Liquid Chromatography
4.5. Cells Culturing and Treatment
4.6. Flow Cytometry
4.7. Redox Components Assessment
4.7.1. Lipid Peroxidation Test
4.7.2. Total Glutathione Levels
4.7.3. Catalase Enzyme Activity
4.7.4. Superoxide Dismutase Enzyme Assay
4.7.5. Glutathione Peroxidase Activity
4.8. Comet Assay (Single-Cell Gel Electrophoresis)
4.9. Gene Transcription Assessment by RT-qPCR
4.9.1. Total RNA Preparation and First-Strand cDNA Synthesis
4.9.2. Real-Time PCR
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kielczykowska, M.; Musik, I. The Great Healing Potential Hidden in Plant Preparations of Antioxidant Properties: A Return to Nature? Oxidative Med. Cell. Longev. 2020, 2020, 8163868. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Gupta, D.; Chawla, R.; Sagar, R.; Sharma, A.; Kumar, R.; Prasad, J.; Singh, S.; Samanta, N.; Sharma, R.K. Radioprotection by plant products: Present status and future prospects. Phytother. Res. 2005, 19, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Dell’Acqua, G.; Schweikert, K. Skin benefits of a myconoside-rich extract from resurrection plant Haberlea rhodopensis. Int. J. Cosmet. Sci. 2012, 34, 132–139. [Google Scholar] [CrossRef]
- Al-Snafi, A.E. Medicinal plants with antioxidant and free radical scavenging effects (part 2): Plant based review. IOSR J. Pharm. 2016, 6, 62–82. [Google Scholar] [CrossRef]
- Al-Snafi, E.A. Immunological Effects of Medicinal Plants: A Review (Part 2). Immunol. Endocr. Metab. Agents Med. Chem. (Discontin.) 2016, 16, 100–121. [Google Scholar] [CrossRef]
- Mihaylova, D.; Bahchevanska, S.; Toneva, V. Examination of the Antioxidant Activity of Haberlea rhodopensis Leaf Extracts and Their Phenolic Constituents. J. Food Biochem. 2013, 37, 255–261. [Google Scholar] [CrossRef]
- Dewanjee, S.; Joardar, S.; Bhattacharjee, N.; Dua, T.K.; Das, S.; Kalita, J.; Manna, P. Edible leaf extract of Ipomoea aquatica Forssk. (Convolvulaceae) attenuates doxorubicin-induced liver injury via inhibiting oxidative impairment, MAPK activation and intrinsic pathway of apoptosis. Food Chem. Toxicol. 2017, 105, 322–336. [Google Scholar] [CrossRef]
- Kuzmanović, N.; Vukojičić, S.; Strid, A.; Stevanović, V. Review of nomenclatural notes in the European Gesneriaceae. Bot. SERBICA 2014, 38, 247–250. [Google Scholar]
- Petrova, G.; Moyankova, D.; Nishii, K.; Forrest, L.; Tsiripidis, I.; Drouzas, A.D.; Djilianov, D.; Moller, M. The European Paleoendemic Haberlea rhodopensis (Gesneriaceae) Has an Oligocene Origin and a Pleistocene Diversification and Occurs in a Long-Persisting Refugial Area in Southeastern Europe. Int. J. Plant Sci. 2015, 176, 499–514. [Google Scholar] [CrossRef]
- Gechev, T.S.; Benina, M.; Obata, T.; Tohge, T.; Sujeeth, N.; Minkov, I.; Hille, J.; Temanni, M.R.; Marriott, A.S.; Bergstrom, E.; et al. Molecular mechanisms of desiccation tolerance in the resurrection glacial relic Haberlea rhodopensis. Cell. Mol. Life Sci. CMLS 2013, 70, 689–709. [Google Scholar] [CrossRef]
- Djilianov, D.; Ivanov, S.; Georgieva, T.; Moyankova, D.; Berkov, S.; Petrova, G.; Mladenov, P.; Christov, N.; Hristozova, N.; Peshev, D.; et al. A Holistic Approach to Resurrection Plants. Haberlea Rhodopensis—A Case Study. Biotechnol. Biotechnol. Equip. 2009, 23, 1414–1416. [Google Scholar] [CrossRef]
- Djilianov, D.; Ivanov, S.; Moyankova, D.; Miteva, L.; Kirova, E.; Alexieva, V.; Joudi, M.; Peshev, D.; Van den Ende, W. Sugar ratios, glutathione redox status and phenols in the resurrection species Haberlea rhodopensis and the closely related non-resurrection species Chirita eberhardtii. Plant Biol. 2011, 13, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Moyankova, D.; Mladenov, P.; Berkov, S.; Peshev, D.; Georgieva, D.; Djilianov, D. Metabolic profiling of the resurrection plant Haberlea rhodopensis during desiccation and recovery. Plant Physiol. 2014, 152, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Muller, J.; Sprenger, N.; Bortlik, K.; Boller, T.; Wiemken, A. Desiccation increases sucrose levels in Ramonda and Haberlea, two genera of resurrection plants in the Gesneriaceae. Plant Physiol. 1997, 100, 153–158. [Google Scholar] [CrossRef]
- Markovska, Y.; Kimenov, G.; Stefanov, K.; Popov, S. Lipid and sterol changes in leaves of Haberlea rhodopensi and Ramonda serbica at transition from biosis into anabiosis and vice versa caused by water stress. Phytochemistry 1992, 31, 2309–2314. [Google Scholar] [CrossRef]
- Radev, R.; Lazarova, G.; Nedialkov, P.; Sokolova, K.; Rukanova, D.; Tsokeva, Z. Study on antibacterial activity of Haberlea rhodopensis. Trakia J. Sci. 2009, 7, 34–36. [Google Scholar]
- Ebrahimi, S.N.; Gafner, F.; Dell’Acqua, G.; Schweikert, K.; Hamburger, M. Flavone 8-C-Glycosides from Haberlea rhodopensis FRIV. (Gesneriaceae). Helv. Chim. Acta 2011, 94, 38–45. [Google Scholar] [CrossRef]
- Zheleva-Dimitrova, D.; Nedialkov, P.; Giresser, U. A Validated HPLC Method for Simultaneous Determination of Caffeoyl Phenylethanoid Glucosides and Flavone 8-C-glycosides in Haberlea rhodopensis. Nat. Prod. Commun. 2016, 11, 791–792. [Google Scholar] [CrossRef]
- Kondeva-Burdina, M.; Zheleva-Dimitrova, D.; Nedialkov, P.; Girreser, U.; Mitcheva, M. Cytoprotective and antioxidant effects of phenolic compounds from Haberlea rhodopensis Friv. (Gesneriaceae). Pharmacogn. Mag. 2013, 9, 294–301. [Google Scholar] [CrossRef]
- Benina, M.; Petrov, V.; Toneva, V.; Teneva, A.; Gechev, T. Molecular Biology and Physiology of the Resurrection Glacial Relic Haberlea rhodopensis. In Biotechnology of Neglected and Underutilized Crops; Jain, S.M., Dutta Gupta, S., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 61–70. [Google Scholar]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, S.; Gencheva, D.; Popov, B.; Grozeva, N.; Zhelyazkova, M. Radioprotective action of resurrection plant Haberlea rhodopensis Friv. (Gesneriaceae) and role of fl avonoids and phenolic acids. Bulg. J. Agric. Sci. 2019, 25, 158–168. [Google Scholar]
- Berkov, S.H.; Nikolova, M.T.; Hristozova, N.I.; Momekov, G.Z.; Ionkova, I.I.; Đilianov, D.L. GC–MS profiling of bioactive extracts from Haberlea rhodopensis: An endemic resurrection plant. J. Serb. Chem. Soc. 2011, 76, 211–220. [Google Scholar] [CrossRef]
- Mihaylova, D.; Bahchevanska, S.T.; Toneva, V.T. Microwave-assisted extraction of flavonoid antioxidants from leaves of Haberlea rhodopensis. J. Int. Sci. Publ. Mater. Methods Technol. 2011, 5, 104–114. [Google Scholar]
- Mihaylova, D.; Lante, A.; Krastanov, A. Total Phenolic Content, Antioxidant and Antimicrobial Activity of Haberlea rhodopensis Extracts Obtained by Pressurized Liquid Extraction. Acta Aliment. Hung 2015, 44, 326–332. [Google Scholar] [CrossRef]
- Moyankova, D.; Hinkov, A.; Georgieva, D.; Shishkov, S.; Djilianov, D. Inhibitory Effect of Extracts from Haberlea rhodopensis Friv. Against Herpes Simplex Virus. Proc. Bulg. Acad. Sci. 2014, 67, 1369–1376. [Google Scholar]
- Popov, B.; Radev, R.; Georgieva, S. In vitro incidence of chromosome aberrations in gamma-irradiated rabbit lymphocytes, treated with Haberlea rhodopensis extract and vitamin C. Bulg. J. Vet. Med. 2010, 13, 148–153. [Google Scholar]
- Popov, B.; Georgieva, S.; Oblakova, M.; Bonev, G. Effects of Haberlea rhodopensis Extract on Antioxidation and Lipid Peroxidation in Rabbits after Exposure to (Co)-C-60-Gamma-Rays. Arch. Biol. Sci. 2013, 65, 91–97. [Google Scholar] [CrossRef]
- Radev, R.; Peychev, L.; Lazarova, G.; Sokolova, K.; Tsokeva, Z.; Radev, S.; Rukanova, D. Haberlea rhodopensis: A plant with multiple pharmacological activities. Trakia J. Sci. 2012, 10, 290–295. [Google Scholar]
- Popov, B.; Dobreva, Z.; Georgieva, S.; Stanilova, S. Enhancement of anti-KLH IgG antibody production in rabbits after treatment with Haberlea rhodopensis extracts. Trakia J. Sci. 2010, 8, 92–97. [Google Scholar]
- Popov, B.; Georgieva, S.; Gadjeva, V. Modulatory effects of total extract of Haberlea rhodopensis against the cyclophosphamide induced genotoxicity in rabbit lymphocytes in vivo. Trakia J. Sci. 2011, 9, 51–57. [Google Scholar]
- Popov, B.; Georgieva, S.; Gadjeva, V.; Petrov, V. Radioprotective, anticlastogenic and antioxidant effects of total extract of Haberlea rhodopensis on rabbit blood samples exposed to gamma radiation in vitro. Rev. Med. Vet Toulouse 2011, 162, 34–39. [Google Scholar]
- Popov, B.N.; Georgieva, S.Y.; Lalchev, S. Radioprotection from Genetic Damages by Resurrection Plant Haberlea rhodopensis—In Vivo/In Vitro Study with Rabbits. Trakia J. Sci. 2012, 10, 41–47. [Google Scholar]
- Georgieva, S.; Popov, B.; Bonev, G. Radioprotective effect of Haberlea rhodopensis (Friv.) leaf extract on gamma-radiation-induced DNA damage, lipid peroxidation and antioxidant levels in rabbit blood. Indian J. Exp. Biol. 2013, 51, 29–36. [Google Scholar] [PubMed]
- Georgieva, S.; Sotirov, L.; Popov, B.; Koynarski, T. Impact of the Haberlea rhodopensis extract on the innate immune system and response in rabbits following KLH-hemocyanin immunization and cyclophosphamide treatment. Turk. J. Vet. Anim. Sci. 2013, 37, 659–663. [Google Scholar] [CrossRef]
- Hayrabedyan, S.; Todorova, K.; Zasheva, D.; Moyankova, D.; Georgieva, D.; Todorova, J.; Djilianov, D. Haberlea rhodopensis has Potential as a New Drug Source Based on its Broad Biological Modalities. Biotechnol. Biotechnol. Equip. 2013, 27, 3553–3560. [Google Scholar] [CrossRef]
- Dobreva, Z.G.; Popov, B.N.; Georgieva, S.Y.; Stanilova, S.A. Immunostimulatory activities of Haberlea rhodopensis leaf extract on the specific antibody response: Protective effects against gamma-radiation-induced immunosuppression. Food Agric. Immunol. 2015, 26, 381–393. [Google Scholar] [CrossRef]
- Zasheva, D.; Mladenov, P.; Rusanov, K.; Simova, S.; Zapryanova, S.; Simova-Stoilova, L.; Moyankova, D.; Djilianov, D. Fractions of Methanol Extracts from the Resurrection Plant Haberlea rhodopensis Have Anti-Breast Cancer Effects in Model Cell Systems. Separations 2023, 10, 388. [Google Scholar] [CrossRef]
- Amirova, K.M.; Dimitrova, P.A.; Marchev, A.S.; Krustanova, S.V.; Simova, S.D.; Alipieva, K.I.; Georgiev, M.I. Biotechnologically-Produced Myconoside and Calceolarioside E Induce Nrf2 Expression in Neutrophils. Int. J. Mol. Sci. 2021, 22, 1759. [Google Scholar] [CrossRef]
- Georgieva, M.; Moyankova, D.; Djilianov, D.; Uzunova, K.; Miloshev, G. Methanol extracts from the resurrection plant Haberlea rhodopensis ameliorate cellular vitality in chronologically ageing Saccharomyces cerevisiae cells. Biogerontology 2015, 16, 461–472. [Google Scholar] [CrossRef]
- Du, Z.Y.; Bramlage, W.J. Modified Thiobarbituric Acid Assay for Measuring Lipid Oxidation in Sugar-Rich Plant-Tissue Extracts. J. Agric. Food Chem. 1992, 40, 1566–1570. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Sergediene, E.; Jonsson, K.; Szymusiak, H.; Tyrakowska, B.; Rietjens, I.M.; Cenas, N. Prooxidant toxicity of polyphenolic antioxidants to HL-60 cells: Description of quantitative structure-activity relationships. FEBS Lett. 1999, 462, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Metodiewa, D.; Jaiswal, A.K.; Cenas, N.; Dickancaite, E.; Segura-Aguilar, J. Quercetin may act as a cytotoxic prooxidant after its metabolic activation to semiquinone and quinoidal product. Free Radic. Biol. Med. 1999, 26, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, S.; Atsumi, T.; Ishihara, M.; Kadoma, Y. Cytotoxicity, ROS-generation activity and radical-scavenging activity of curcumin and related compounds. Anticancer Res. 2004, 24, 563–569. [Google Scholar] [PubMed]
- Jackson, S.P.; Bartek, J. The DNA-damage response in human biology and disease. Nature 2009, 461, 1071–1078. [Google Scholar] [CrossRef]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB J. 2003, 17, 1195–1214. [Google Scholar] [CrossRef]
- Srivastava, N.N.; Shukla, S.K.; Yashavarddhan, M.H.; Devi, M.; Tripathi, R.P.; Gupta, M.L. Modification of radiation-induced DNA double strand break repair pathways by chemicals extracted from Podophyllum hexandrum: An in vitro study in human blood leukocytes. Environ. Mol. Mutagen. 2014, 55, 436–448. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide Radical and Superoxide Dismutases. Annu. Rev. Biochem. 1995, 64, 97–112. [Google Scholar] [CrossRef]
- Kostadinova, A.; Hazarosova, R.; Topouzova-Hristova, T.; Moyankova, D.; Yordanova, V.; Veleva, R.; Nikolova, B.; Momchilova, A.; Djilianov, D.; Staneva, G. Myconoside interacts with the plasma membranes and the actin cytoskeleton and provokes cytotoxicity in human lung adenocarcinoma A549 cells. J. Bioenerg. Biomembr. 2022, 54, 31–43. [Google Scholar] [CrossRef]
- Gentile, F.; Arcaro, A.; Pizzimenti, S.; Daga, M.; Cetrangolo, G.P.; Dianzani, C.; Lepore, A.; Graf, M.; Ames, P.R.J.; Barrera, G. DNA damage by lipid peroxidation products: Implications in cancer, inflammation and autoimmunity. AIMS Genet. 2017, 4, 103–137. [Google Scholar] [CrossRef]
- Mihailova, G.; Petkova, S.; Georgieva, K. Changes in Some Antioxidant Enzyme Activities in Haberlea rhodopensis during Desiccation at High Temperature. Biotechnol. Biotechnol. Equip. 2009, 23, 561–564. [Google Scholar] [CrossRef]
- Yahubyan, G.; Gozmanova, M.; Denev, I.; Toneva, V.; Minkov, I. Prompt response of superoxide dismutase and peroxidase to dehydration and rehydration of the resurrection plant Haberlea rhodopensis. Plant Growth Regul. 2009, 57, 49–56. [Google Scholar] [CrossRef]
- Khan, S.G.; Katiyar, S.K.; Agarwal, R.; Mukhtar, H. Enhancement of antioxidant and phase II enzymes by oral feeding of green tea polyphenols in drinking water to SKH-1 hairless mice: Possible role in cancer chemoprevention. Cancer Res. 1992, 52, 4050–4052. [Google Scholar] [PubMed]
- Georgiev, Y.N.; Ognyanov, M.H.; Denev, P.N. The ancient Thracian endemic plant Haberlea rhodopensis Friv. and related species: A review. J. Ethnopharmacol. 2020, 249, 112359. [Google Scholar] [CrossRef]
- Bhosle, S.M.; Huilgol, N.G.; Mishra, K.P. Enhancement of radiation-induced oxidative stress and cytotoxicity in tumor cells by ellagic acid. Clin. Chim. Acta 2005, 359, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Shiloh, Y.; Ziv, Y. The ATM protein kinase: Regulating the cellular response to genotoxic stress, and more. Nat. Rev. Mol. Cell Biol. 2013, 14, 197–210. [Google Scholar] [CrossRef]
- Huang, R.X.; Zhou, P.K. DNA damage response signaling pathways and targets for radiotherapy sensitization in cancer. Signal Transduct. Target. Ther. 2020, 5, 60. [Google Scholar] [CrossRef]
- Kastenhuber, E.R.; Lowe, S.W. Putting p53 in Context. Cell 2017, 170, 1062–1078. [Google Scholar] [CrossRef]
- Guleria, A.; Chandna, S. ATM kinase: Much more than a DNA damage responsive protein. DNA Repair 2016, 39, 1–20. [Google Scholar] [CrossRef]
- Klopfleisch, R.; Gruber, A.D. Increased expression of BRCA2 and RAD51 in lymph node metastases of canine mammary adenocarcinomas. Vet. Pathol. 2009, 46, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Nakano, K.; Vousden, K.H. PUMA, a novel proapoptotic gene, is induced by p53. Mol. Cell 2001, 7, 683–694. [Google Scholar] [CrossRef]
- Li, M.J.; Wang, W.W.; Chen, S.W.; Shen, Q.; Min, R. Radiation dose effect of DNA repair-related gene expression in mouse white blood cells. Med. Sci. Monit. 2011, 17, Br290–Br297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, F.; Thakur, K.; Wang, J.; Wang, H.; Hu, F.; Zhang, J.G.; Wei, Z.J. Molecular mechanism of anti-cancerous potential of Morin extracted from mulberry in Hela cells. Food Chem. Toxicol. 2018, 112, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Dimitrova, N.; Staneva, D.; Popov, B.; Alexandrova, A.; Georgieva, M.; Miloshev, G. Haberlea rhodopensis alcohol extract normalizes stress-responsive transcription of the human TP53 gene. J. Exp. Biol. Agric. Sci. 2023, 11, 405–415. [Google Scholar] [CrossRef]
- Rezaeejam, H.; Shirazi, A.; Valizadeh, M.; Izadi, P. Candidate gene biodosimeters of mice and human exposure to ionizing radiation by quantitative reverse transcription polymerase chain reaction. J. Cancer Res. Ther. 2015, 11, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Kabacik, S.; Mackay, A.; Tamber, N.; Manning, G.; Finnon, P.; Paillier, F.; Ashworth, A.; Bouffler, S.; Badie, C. Gene expression following ionising radiation: Identification of biomarkers for dose estimation and prediction of individual response. Int. J. Radiat. Biol. 2011, 87, 115–129. [Google Scholar] [CrossRef]
- Filiano, A.N.; Fathallah-Shaykh, H.M.; Fiveash, J.; Gage, J.; Cantor, A.; Kharbanda, S.; Johnson, M.R. Gene Expression Analysis in Radiotherapy Patients and C57BL/6 Mice as a Measure of Exposure to Ionizing Radiation. Radiat. Res. 2011, 176, 49–61. [Google Scholar] [CrossRef]
- Mahaney, B.L.; Meek, K.; Lees-Miller, S.P. Repair of ionizing radiation-induced DNA double-strand breaks by non-homologous end-joining. Biochem. J. 2009, 417, 639–650. [Google Scholar] [CrossRef]
- Kan, C.; Zhang, J. BRCA1 Mutation: A Predictive Marker for Radiation Therapy? Int. J. Radiat. Oncol. Biol. Phys. 2015, 93, 281–293. [Google Scholar] [CrossRef]
- Khanna, K.K.; Jackson, S.P. DNA double-strand breaks: Signaling, repair and the cancer connection. Nat. Genet. 2001, 27, 247–254. [Google Scholar] [CrossRef]
- Pfeiffer, P.; Goedecke, W.; Kuhfittig-Kulle, S.; Obe, G. Pathways of DNA double-strand break repair and their impact on the prevention and formation of chromosomal aberrations. Cytogenet. Genome Res. 2004, 104, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Mladenov, E.; Magin, S.; Soni, A.; Iliakis, G. DNA double-strand break repair as determinant of cellular radiosensitivity to killing and target in radiation therapy. Front. Oncol. 2013, 3, 113. [Google Scholar] [CrossRef]
- Jeggo, P.A.; Geuting, V.; Lobrich, M. The role of homologous recombination in radiation-induced double-strand break repair. Radiother. Oncol. 2011, 101, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Boboila, C.; Alt, F.W.; Schwer, B. Classical and alternative end-joining pathways for repair of lymphocyte-specific and general DNA double-strand breaks. Adv. Immunol. 2012, 116, 1–49. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.X. BRCA1: Cell cycle checkpoint, genetic instability, DNA damage response and cancer evolution. Nucleic Acids Res. 2006, 34, 1416–1426. [Google Scholar] [CrossRef]
- Abbott, D.W.; Thompson, M.E.; Robinson-Benion, C.; Tomlinson, G.; Jensen, R.A.; Holt, J.T. BRCA1 expression restores radiation resistance in BRCA1-defective cancer cells through enhancement of transcription-coupled DNA repair. J. Biol. Chem. 1999, 274, 18808–18812. [Google Scholar] [CrossRef]
- Ding, S.L.; Sheu, L.F.; Yu, J.C.; Yang, T.L.; Chen, B.F.; Leu, F.J.; Shen, C.Y. Abnormality of the DNA double-strand-break checkpoint/repair genes, ATM, BRCA1 and TP53, in breast cancer is related to tumour grade. Br. J. Cancer 2004, 90, 1995–2001. [Google Scholar] [CrossRef]
- Cao, L.; Kim, S.; Xiao, C.; Wang, R.H.; Coumoul, X.; Wang, X.; Li, W.M.; Xu, X.L.; De Soto, J.A.; Takai, H.; et al. ATM-Chk2-p53 activation prevents tumorigenesis at an expense of organ homeostasis upon Brca1 deficiency. EMBO J. 2006, 25, 2167–2177. [Google Scholar] [CrossRef]
- Hunter, F.E., Jr.; Gebicki, J.M.; Hoffsten, P.E.; Weinstein, J.; Scott, A. Swelling and lysis of rat liver mitochondria induced by ferrous ions. J. Biol. Chem. 1963, 238, 828–835. [Google Scholar] [CrossRef]
- Tietze, F. Enzymic method for quantitative determination of nanogram amounts of total and oxidized glutathione: Applications to mammalian blood and other tissues. Anal. Biochem. 1969, 27, 502–522. [Google Scholar] [CrossRef]
- Aebi, H. Catalase Invitro. Method Enzym. 1984, 105, 121–126. [Google Scholar]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on Quantitative and Qualitative Characterization of Erythrocyte Glutathione Peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar] [PubMed]
- Lovell, D.P. Statistical Analysis of Comet Assay Data. Issues Toxicol. 2009, 5, 424–450. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | NCBI Ref Seq | Oligonucleotide Sequence 5′-3′ | Amplicon, bp |
|---|---|---|---|
| ATM | NM_000051.3 | For TGCTGTGAGAAAACCATGGAAGTGA Rev TCCGGCCTCTGCTGTAAATACAAAG | 137 |
| BRCA1 | NM_007294.3 | For CACCCAATTGTGGTTGTGCAGC Rev GTCCAGCTCCTGGCACTGGTAGAG | 141 |
| p21 (CDKN1A) | NM_000389.4 | For AGAGGAAGACCATGTGGACCTGTCA Rev AGAAATCTGTCATGCTGGTCTGCC | 134 |
| BBC3 (PUMA) | NM_014417.4 | For GGATGGCGGACGACCTCAA Rev GGGTAAGGGCAGGAGTCCCATG | 119 |
| RAD50 | NM_005732.3 | For TGGTGATGCTGAAGGGAGACACA Rev TTGTTGGCTCATCCAAGGCAATG | 147 |
| RAD51 | NM_133487.3 | For CAAGCATCAGCCATGATGGTAGAA Rev AGAAACCTGGCCAAGTGCATCTG | 132 |
| GAPDH | NM_002046.5 | For ACCAGGTGGTCTCCTCTGACTTCAA Rev ACCCTGTTGCTGTAGCCAAATTCG | 136 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staneva, D.; Dimitrova, N.; Popov, B.; Alexandrova, A.; Georgieva, M.; Miloshev, G. Haberlea rhodopensis Extract Tunes the Cellular Response to Stress by Modulating DNA Damage, Redox Components, and Gene Expression. Int. J. Mol. Sci. 2023, 24, 15964. https://doi.org/10.3390/ijms242115964
Staneva D, Dimitrova N, Popov B, Alexandrova A, Georgieva M, Miloshev G. Haberlea rhodopensis Extract Tunes the Cellular Response to Stress by Modulating DNA Damage, Redox Components, and Gene Expression. International Journal of Molecular Sciences. 2023; 24(21):15964. https://doi.org/10.3390/ijms242115964
Chicago/Turabian StyleStaneva, Dessislava, Neli Dimitrova, Borislav Popov, Albena Alexandrova, Milena Georgieva, and George Miloshev. 2023. "Haberlea rhodopensis Extract Tunes the Cellular Response to Stress by Modulating DNA Damage, Redox Components, and Gene Expression" International Journal of Molecular Sciences 24, no. 21: 15964. https://doi.org/10.3390/ijms242115964
APA StyleStaneva, D., Dimitrova, N., Popov, B., Alexandrova, A., Georgieva, M., & Miloshev, G. (2023). Haberlea rhodopensis Extract Tunes the Cellular Response to Stress by Modulating DNA Damage, Redox Components, and Gene Expression. International Journal of Molecular Sciences, 24(21), 15964. https://doi.org/10.3390/ijms242115964