
Molecular Mechanisms for Changing Brain Connectivity in Mice and Humans


Abstract
:1. Introduction
2. Results
2.1. Training
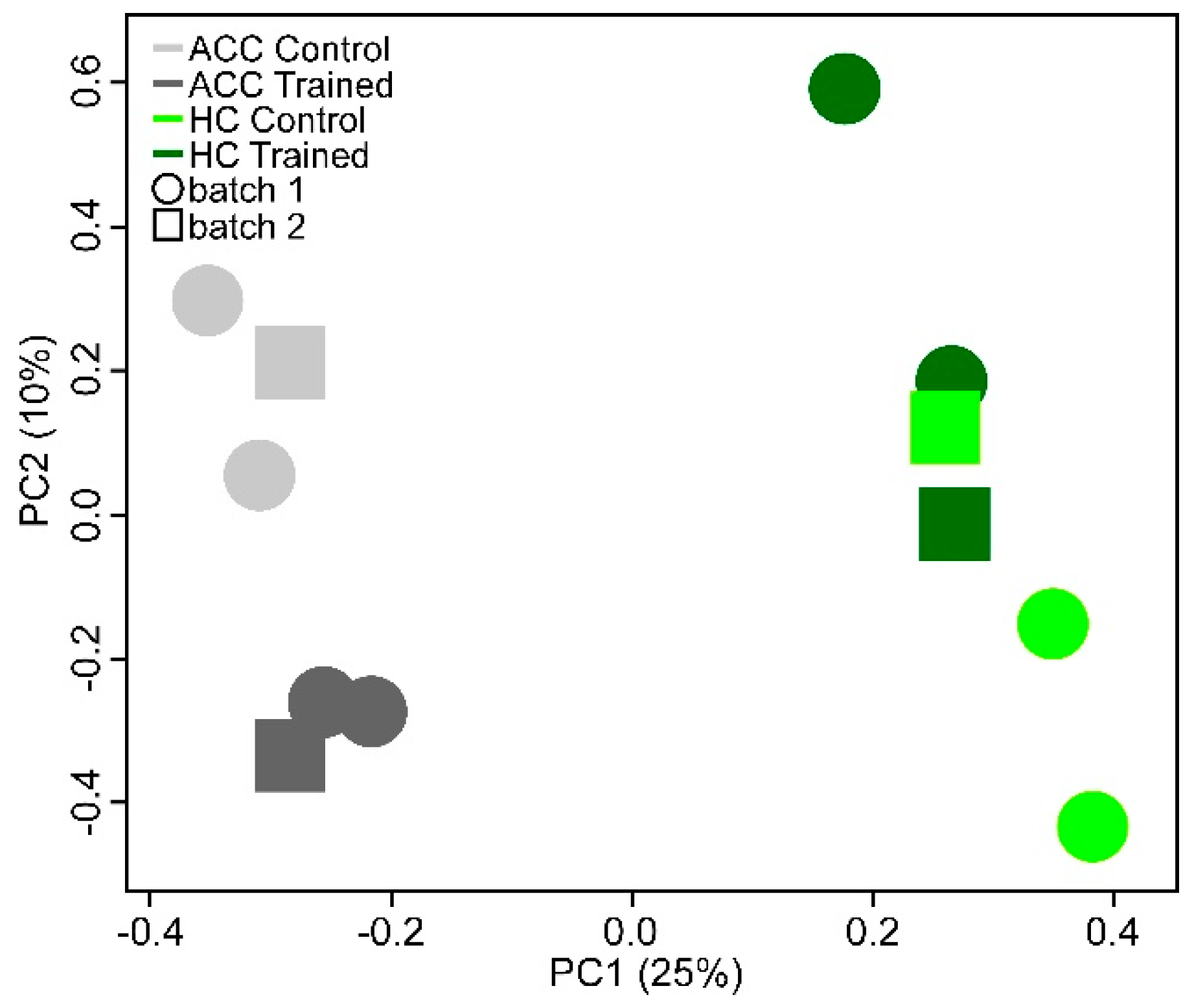
2.2. Gene Expression in the Mouse Brain
2.3. Gene Expression in Mouse Blood
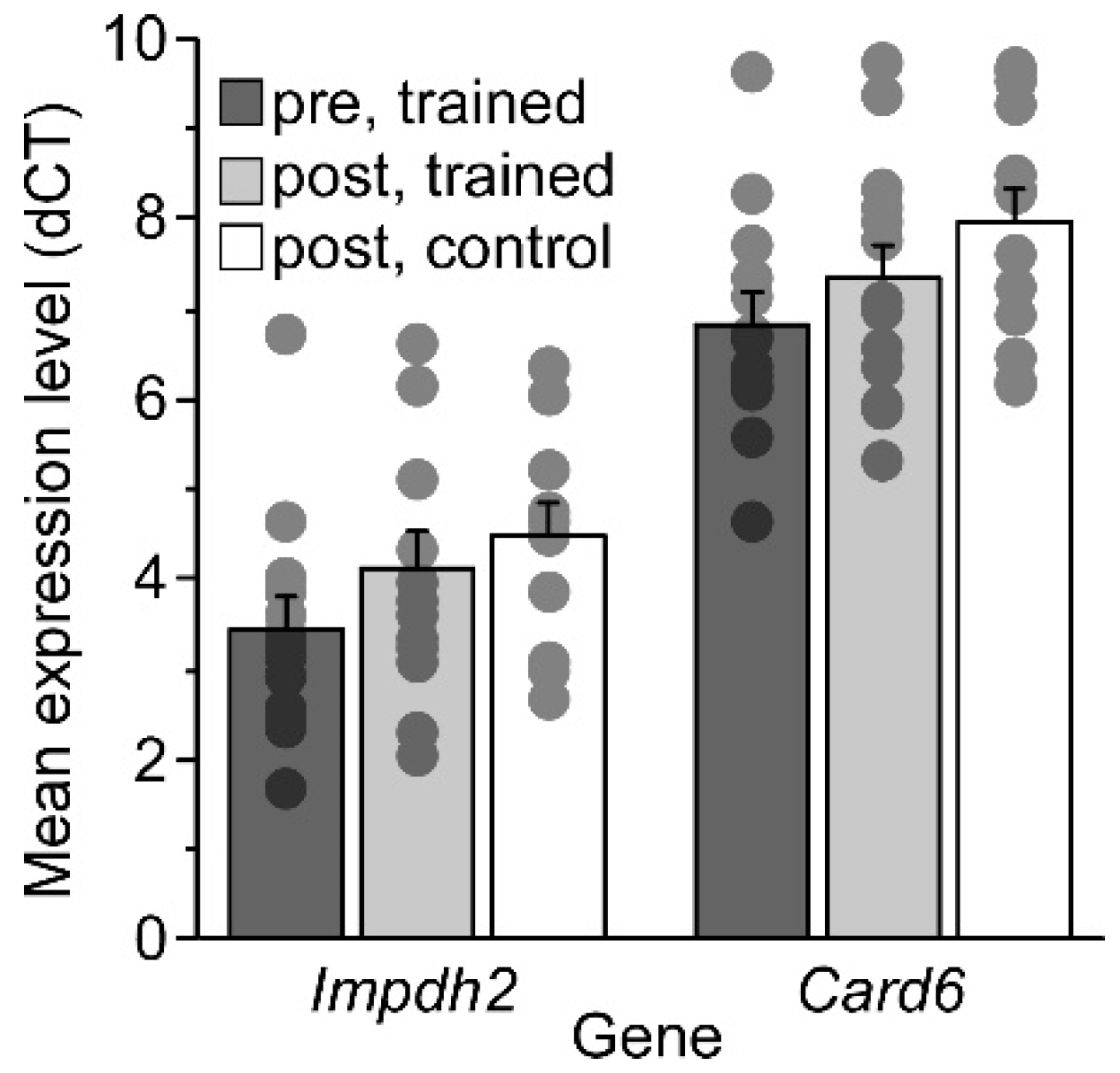
2.4. Mouse RT-PCR
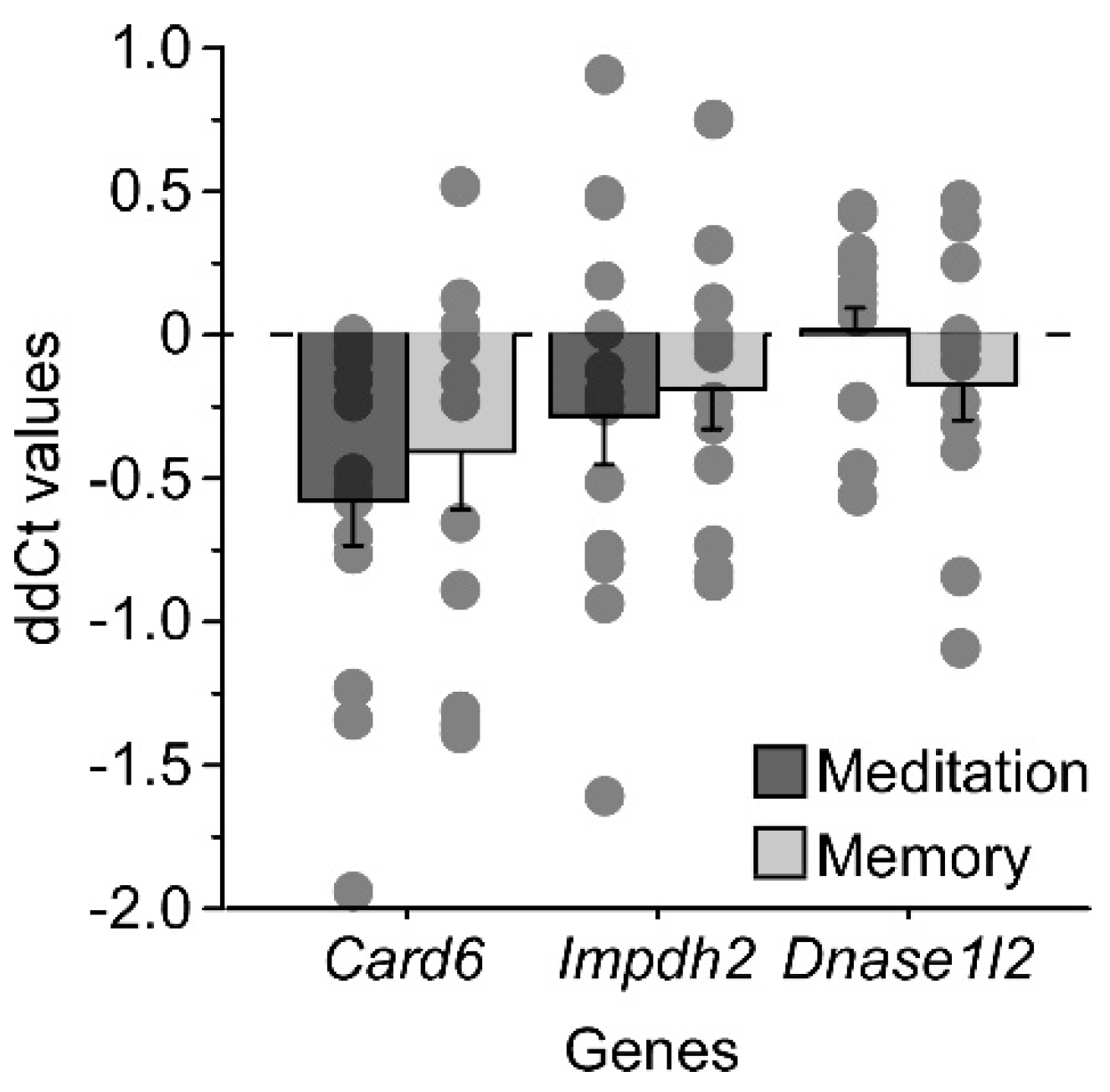
2.5. Genetic Expression in Human Blood
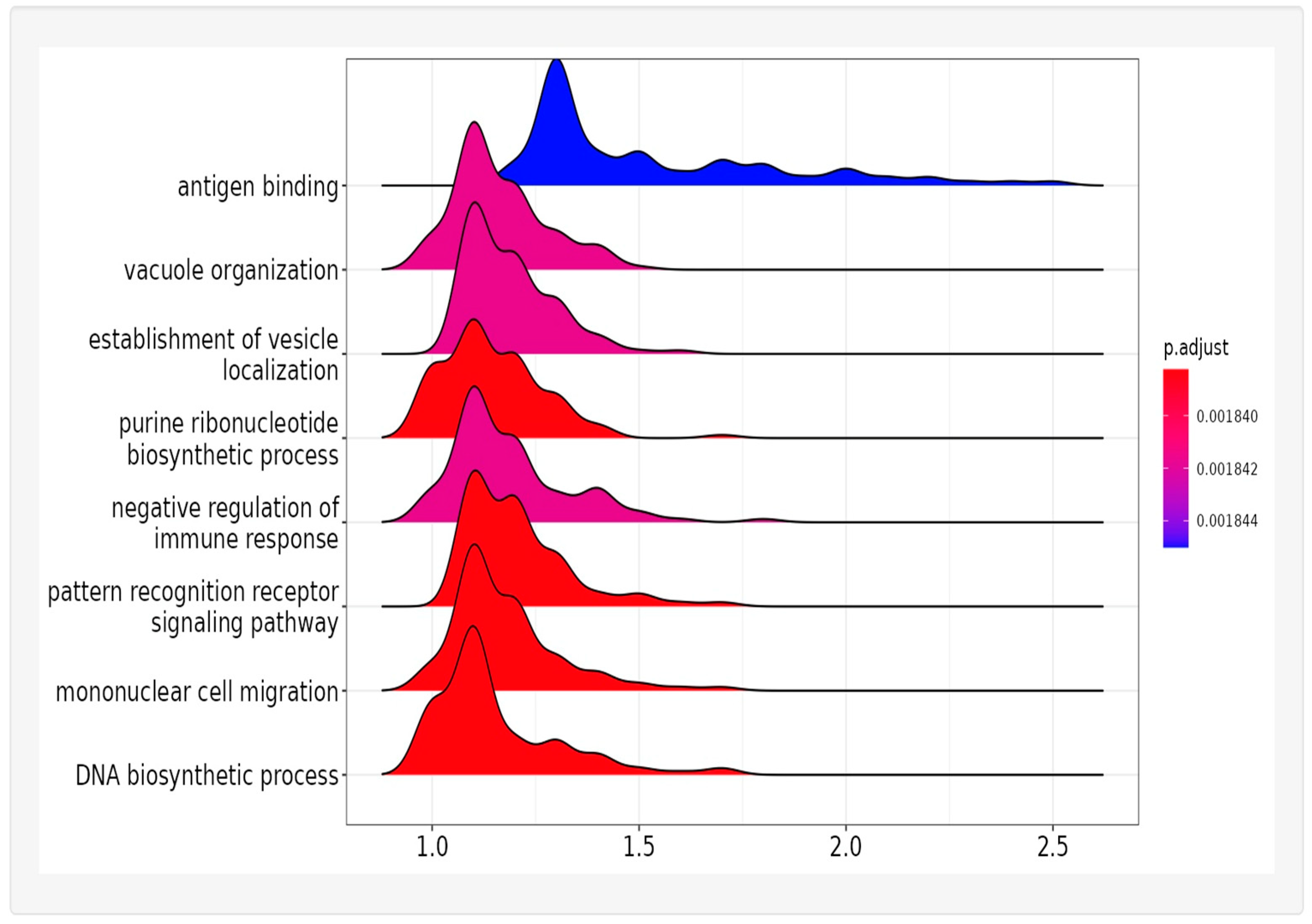
2.6. Pathways
3. Discussion
3.1. Mice
3.2. Humans
3.3. Common Pathway
4. Materials and Methods
4.1. Mice
4.2. Surgery
4.3. Training and Sample Harvesting
4.4. RNA Preparation/Microarray
4.5. Microarray Analysis
4.6. Gene Set Enrichment Analysis (GSEA)
4.7. Real-Time PCR (Mouse)
4.8. Human
4.9. Real-Time PCR (Human)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barth, A.L.; Ray, A. Progressive Circuit Changes during Learning and Disease. Neuron 2019, 104, 37–46. [Google Scholar] [CrossRef]
- Amoah, D.K. Advances in the understanding and enhancement of the human cognitive functions of learning and memory. Brain Sci. Adv. 2022, 8, 276–297. [Google Scholar] [CrossRef]
- Liu, Q.; Wu, Y.; Wang, H.; Jia, F.; Xu, F. Viral Tools for Neural Circuit Tracing. Neurosci. Bull. 2022, 38, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Saunders, A.; Macosko, E.Z.; Wysoker, A.; Goldman, M.; Krienen, F.M.; de Rivera, H.; Bien, E.; Baum, M.; Bortolin, L.; Wang, S.; et al. Molecular Diversity and Specializations among the Cells of the Adult Mouse Brain. Cell 2018, 174, 1015–1030.e16. [Google Scholar] [CrossRef] [PubMed]
- Posner, M.I.; Sheese, B.E.; Odludaş, Y.; Tang, Y. Analyzing and shaping human attentional networks. Neural Netw. 2006, 19, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Cullen, C.L.; Pepper, R.E.; Clutterbuck, M.T.; Pitman, K.A.; Oorschot, V.; Auderset, L.; Tang, A.D.; Ramm, G.; Emery, B.; Rodger, J.; et al. Periaxonal and nodal plasticities modulate action potential conduction in the adult mouse brain. Cell Rep. 2021, 34, 108641. [Google Scholar] [CrossRef]
- Xin, W.; Chan, J.R. Myelin plasticity: Sculpting circuits in learning and memory. Nat. Rev. Neurosci. 2020, 21, 682–694. [Google Scholar] [CrossRef]
- Tang, Y.; Posner, M.I. Attention training and Attention State Training. Trends Cogn. Sci. 2002, 13, 222–227. [Google Scholar] [CrossRef]
- Melby-Lervåg, M.; Hulme, C. Is working memory training effective? A meta-analytic review. Dev. Psychol. 2013, 49, 270–291. [Google Scholar] [CrossRef]
- Tang, Y.Y.; Holzel, B.K.; Posner, M.I. The neuroscience of mindfulness meditation. Nat. Rev. Neurosci. 2015, 16, 213–225. [Google Scholar] [CrossRef]
- Posner, M.I.; Weible, A.P.; Voelker, P.; Rothbart, M.K.; Niell, C.M. Decision Making as a Learned Skill in Mice and Humans. Front. Neurosci. 2022, 16, 834701. [Google Scholar] [CrossRef] [PubMed]
- Weible, A.P. Remembering to Attend: The Anterior Cingulate Cortex and Remote Memory. Behav. Brain Res. 2013, 245, 63–75. [Google Scholar] [CrossRef]
- Xu, W.; Südhof, T.C. A Neural Circuit for Memory Specificity and Generalization. Science 2013, 339, 1290–1295. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.C.; Ochsner, K.N.; Kuhl, B.; Cooper, J.; Robertson, E.; Gabrieli, S.W.; Glover, G.H.; Gabrieli, J.D.E. Neural Systems Underlying the Suppression of Unwanted Memories. Science 2004, 303, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Bowman, C.R.; Zeithamova, D. Abstract Memory Representations in the Ventromedial Prefrontal Cortex and Hippocampus Support Concept Generalization. J. Neurosci. 2018, 38, 2605–2614. [Google Scholar] [CrossRef] [PubMed]
- Ciaramelli, E.; Grady, C.; Levine, B.; Ween, J.; Moscovitch, M. Top-Down and Bottom-Up Attention to Memory Are Dissociated in Posterior Parietal Cortex: Neuroimaging and Neuropsychological Evidence. J. Neurosci. 2010, 30, 4943–4956. [Google Scholar] [CrossRef]
- Weible, A.P.; Posner, M.I.; Neill, C.M. Differential involvement of three brain regions during mouse skill learning. eNeuro 2019, 6, 1–12. [Google Scholar] [CrossRef]
- Tang, Y.Y.; Posner, M.I. Training brain networks and states. Trends Cogn. Sci. 2014, 18, 345–350. [Google Scholar] [CrossRef]
- Tang, Y.Y.; Lu, Q.; Geng, X.; Stein, E.A.; Yang, Y.; Posner, M.I. Short-term meditation induces white matter changes in the anterior cingulate. Proc. Natl. Acad. Sci. USA 2010, 107, 15649–15652. [Google Scholar] [CrossRef]
- Tang, Y.Y.; Ma, Y.; Wang, J.; Fan, Y.; Feng, S.; Lu, Q.; Yu, Q.; Sui, D.; Rothbart, M.K.; Fan, M.; et al. Short term meditation training improves attention and self-regulation. Proc. Natl. Acad. Sci. USA 2007, 104, 17152–17156. [Google Scholar] [CrossRef]
- Sánchez-Pérez, N.; Inuggi, A.; Castillo, A.; Campoy, G.; García-Santos, J.M.; González-Salinas, C.; Fuentes, L.J. Computer-Based Cognitive Training Improves Brain Functional Connectivity in the Attentional Networks: A Study with Primary School-Aged Children. Front. Behav. Neurosci. 2019, 13, 247. [Google Scholar] [CrossRef] [PubMed]
- Román, F.J.; Iturria-Medina, Y.; Martínez, K.; Karama, S.; Burgaleta, M.; Evans, A.C.; Jaeggi, S.M.; Colom, R. Enhanced structural connectivity within a brain sub-network supporting working memory and engagement processes after cognitive training. Neurobiol. Learn. Mem. 2017, 141, 33–43. [Google Scholar] [CrossRef]
- Bontempi, B.; Laurent-Demir, C.; Destrade, C.; Jaffard, R. Time-dependent reorganization of brain circuitry underlying long-term memory storage. Nature 1999, 400, 671–675. [Google Scholar] [CrossRef]
- Frankland, P.W.; Bontempi, B.; Talton, L.E.; Kaczmarek, L.; Silva, A.J. The involvement of the anterior cingulate cortex in remote contextual fear memory. Science 2004, 304, 881–883. [Google Scholar] [CrossRef]
- Rajasethupathy, P.; Sankaran, S.; Marshel, J.H.; Kim, C.K.; Ferenczi, E.; Lee, S.Y.; Berndt, A.; Ramakrishnan, C.; Jaffe, A.; Lo, M.; et al. Projections from neocortex mediate top-down control of memory retrieval. Nature 2015, 526, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fukushima, H.; Kida, S. Induction and requirement of gene expression in the anterior cingulate cortex and medial prefrontal cortex for the consolidation of inhibitory avoidance memory. Mol. Brain 2011, 4, 4. [Google Scholar] [CrossRef]
- Tanimizu, T.; Kenney, J.W.; Okano, E.; Kadoma, K.; Frankland, P.W.; Kida, S. Functional Connectivity of Multiple Brain Regions Required for the Consolidation of Social Recognition Memory. J. Neurosci. 2017, 37, 4103–4116. [Google Scholar] [CrossRef]
- Nadler, J.J.; Zou, F.; Huang, H.; Moy, S.S.; Lauder, J.; Crawley, J.N.; Threadgill, D.W.; Wright, F.A.; Magnuson, T.R. Large-scale gene expression differences across brain regions and inbred strains correlate with a behavioral phenotype. Genetics 2006, 174, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Langfelder, P.; Fuller, T.F.; Oldham, M.C.; Luo, R.; van den Berg, L.H.; Ophoff, R.A.; Horvath, S. Is human blood a good surrogate for brain tissue in transcriptional studies? BMC Genom. 2010, 11, 589. [Google Scholar] [CrossRef]
- Harris, S.E.; Cox, S.R.; Bell, S.; Marioni, R.E.; Prins, B.P.; Pattie, A.; Corley, J.; Muñoz Maniega, S.; Valdés Hernández, M.; Morris, Z.; et al. Eurology-related protein biomarkers are associated with cognitive ability and brain volume in older age. Nat. Commun. 2020, 11, 800. [Google Scholar] [CrossRef]
- Vrillon, A.; Mouton-Liger, F.; Martinet, M.; Cognate, E.; Hourregue, C.; Dumurgier, J.; Bouaziz-Amar, E.; Brinkmalm, A.; Blennow, K.; Zetterberg, H.; et al. Plasma neuregulin 1 as a synaptic biomarker in Alzheimer’s disease: A discovery cohort study. Alzheimer’s Res. Ther. 2022, 14, 71. [Google Scholar] [CrossRef] [PubMed]
- McPhee, G.M.; Downey, L.A.; Stough, C. Neurotrophins as a reliable biomarker for brain function, structure and cognition: A systematic review and meta-analysis. Neurobiol. Learn. Mem. 2020, 175, 107298. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.; Jarvis, P. Review of the potential use of blood neuro-biomarkers in the diagnosis of mild traumatic brain injury. Clin. Exp. Emerg. Med. 2017, 4, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Pedreño, C.; Obiols, J.; Taurines, R.; Pàmias, M.; Grünblatt, E.; Gella, A. Neurotrophin blood-based gene expression and social cognition analysis in patients with autism spectrum disorder. Neurogens 2015, 16, 123–131. [Google Scholar] [CrossRef]
- Santiago, J.A.; Bottero, V.; Potashkin, J.A. Evaluation of RNA Blood Biomarkers in the Parkinson’s Disease Biomarkers Program. Front. Aging Neurosci. 2018, 10, 157. [Google Scholar] [CrossRef]
- Atif, H.; Hicks, S.D. A Review of MicroRNA Biomarkers in Traumatic Brain Injury. J. Exp. Neurosci. 2019, 13, 1179069519832286. [Google Scholar] [CrossRef]
- Ma, G.; Liu, M.; Du, K.; Zhong, X.; Gong, S.; Jiao, L.; Wei, M. Differential Expression of mRNAs in the Brain Tissues of Patients with Alzheimer’s Disease Based on GEO Expression Profile and Its Clinical Significance. BioMed Res. Int. 2019, 2019, 8179145. [Google Scholar] [CrossRef]
- Milanesi, E.; Cucos, C.A.; Matias-Guiu, J.A.; Piñol-Ripoll, G.; Manda, G.; Dobre, M.; Cuadrado, A. Reduced Blood RGS2 Expression in Mild Cognitive Impairment Patients. Front. Aging Neurosci. 2021, 13, 738244. [Google Scholar] [CrossRef]
- Pérez-González, M.; Badesso, S.; Lorenzo, E.; Guruceaga, E.; Pérez-Mediavilla, A.; García-Osta, A.; Cuadrado-Tejedor, M. Identifying the Main Functional Pathways Associated with Cognitive Resilience to Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 9120. [Google Scholar] [CrossRef]
- Klein, A.B.; Williamson, R.; Santini, M.A.; Clemmensen, C.; Ettrup, A.; Rios, M.; Knudsen, G.M.; Aznar, S. Blood BDNF concentrations reflect brain-tissue BDNF levels across species. Int. J. Neuropsychopharmacol. 2011, 14, 347–353. [Google Scholar] [CrossRef]
- Amagase, Y.; Kambayashi, R.; Sugiyama, A.; Takei, Y. Peripheral Regulation of Central Brain-Derived Neurotrophic Factor Expression through the Vagus Nerve. Int. J. Mol. Sci. 2023, 24, 3543. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Dufner, A.; Pownall, S.; Mak, T.W. Caspase recruitment domain protein 6 is a microtubule-interacting protein that positively modulates NF-kappaB activation. Proc. Natl. Acad. Sci. USA 2006, 103, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Toubiana, J.; Rossi, A.L.; Grimaldi, D.; Belaidouni, N.; Chafey, P.; Clary, G.; Courtine, E.; Pene, F.; Mira, J.P.; Claessens, Y.E.; et al. IMPDHII protein inhibits Toll-like receptor 2-mediated activation of NF-kappaB. J. Biol. Chem. 2011, 286, 23319–23333. [Google Scholar] [CrossRef]
- Shiokawa, D.; Matsushita, T.; Kobayashi, T.; Matsumoto, Y.; Tanuma, S. Characterization of the human DNAS1L2 gene and the molecular mechanism for its transcriptional activation induced by inflammatory cytokines. Genomics 2004, 84, 95–105. [Google Scholar] [CrossRef]
- Vijayendran, C.; Bermudez, M.-L.; Koka, M.; Chandran, B.; Pawale, D.; Vishnubhotla, R.; Alankar, S.; Maturi, R.; Subramaniam, B.; Sadhasivam, S. Large-scale genomic study reveals robust activation of the immune system following advanced Inner Engineering meditation retreat. Proc. Natl. Acad. Sci. USA 2021, 118, e2110455118. [Google Scholar] [CrossRef]
- Bonefeld, B.E.; Elfving, B.; Wegener, G. Reference genes for normalization: A study of rat brain tissue. Synapse 2008, 62, 302–309. [Google Scholar] [CrossRef]
- Gladkikh, A.; Potashnikova, D.; Korneva, E.; Khudoleeva, O.; Vorobjev, I. Cyclin D1 expression in B-cell lymphomas. Exp. Hematol. 2010, 38, 1047–1057. [Google Scholar] [CrossRef]
- Yan, Z.; Gao, J.; Lv, X.; Yang, W.; Wen, S.; Tong, H.; Tang, C. Quantitative Evaluation and Selection of Reference Genes for Quantitative RT-PCR in Mouse Acute Pancreatitis. BioMed Res. Int. 2016, 2016, 8367063, Erratum in BioMed Res. Int. 2017, 2017, 3498537. [Google Scholar] [CrossRef]
- Yousif, A.; Drou, N.; Rowe, J.; Khalfan, M.; Gunsalus, K.C. NASQAR: A web-based platform for high-throughput sequencing data analysis and visualization. BMC Bioinform. 2020, 21, 267. [Google Scholar] [CrossRef]
- Capece, D.; Verzella, D.; Flati, I.; Arboreta, P.; Cornice, J.; Franzoso, G. NF-κB: Blending metabolism, immunity, and inflammation. Trends Immunol. 2022, 43, 757–775. [Google Scholar] [CrossRef] [PubMed]
- Calabria, E.; Mazza, E.M.; Dyar, K.A.; Pogliaghi, S.; Bruseghini, P.; Morandi, C.; Salvagno, G.L.; Gelati, M.; Guidi, G.C.; Bicciato, S.; et al. Aging: A portrait from gene expression profile in blood cells. Aging 2016, 8, 1802–1821. [Google Scholar] [CrossRef] [PubMed]
- Jaeggi, S.M.; Buschkuehl, M.; Jonides, J.; Perrig, W.J. Improving fluid intelligence with training on working memory. Proc. Natl. Acad. Sci. USA 2008, 105, 6829–6833. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.Y.; Ma, Y.; Fan, Y.; Feng, H.; Wang, J.; Feng, S.; Lu, Q.; Hu, B.; Lin, Y.; Li, J.; et al. Central and autonomic nervous system interaction is altered by short-term meditation. Proc. Natl. Acad. Sci. USA 2009, 106, 8865–8870. [Google Scholar] [CrossRef] [PubMed]
- Long, K.R.; Husttner, W.B. How the extracellular matrix shapes neural development. Open Biol. 2019, 9, 180216. [Google Scholar] [CrossRef] [PubMed]
- Chernousov, M.A.; Rothblum, K.; Stahl, R.C.; Evans, A.; Prentiss, L.; Carey, D.J. Glypican-1 and alpha4(V) collagen are required for Schwann cell myelination. J. Neurosci. 2006, 26, 508–517. [Google Scholar] [CrossRef]
- Hubert, T.; Grimal, S.; Carroll, P.; Fichard-Carroll, A. Collagens in the developing and diseased nervous system. Cell Mol. Life Sci. 2009, 66, 1223–1238. [Google Scholar] [CrossRef]
- Fox, M.A. Novel roles for collagens in wiring the vertebrate nervous system. Curr. Opin. Cell Biol. 2008, 20, 508–513. [Google Scholar] [CrossRef]
- Kakoi, C.; Udo, H.; Matsukawa, T.; Ohnuki, K. Collagen peptides enhance hippocampal neurogenesis and reduce anxiety related behavior in mice. Biomed. Res. 2012, 33, 273–279. [Google Scholar] [CrossRef]
- Shin, S.S.; Grandhi, R.; Henchir, J.; Yan, H.Q.; Badylak, S.F.; Dixon, C.E. Neuroprotective effects of collagen matrix in rats after traumatic brain injury. Restor. Neurol. Neurosci. 2015, 33, 95–104. [Google Scholar] [CrossRef]
- Stehlik, C.; Hayashi, H.; Pio, F.; Godzik, A.; Reed, J.C. Card6 is a modulator of NF-kappa B activation by Nod1- and Cardiak-mediated pathways. J. Biol. Chem. 2003, 278, 31941–31949. [Google Scholar] [CrossRef]
- Zhao, M.; He, H.; Yin, J. Card6 protects against collagen-induced rheumatoid arthritis in mice through attenuating the inflammatory response and joint destruction via suppression of TNFR1/TRAF2 signaling. Biochem. Biophys. Res. Commun. 2020, 526, 1092–1099. [Google Scholar] [CrossRef]
- Wang, J.L.; Luo, X.; Liu, L. Targeting Card6 attenuates spinal cord injury (SCI) in mice through inhibiting apoptosis, inflammation and oxidative stress associated ROS production. Aging 2019, 11, 12213–12235. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.; Nakamura, H.; Natsumeda, Y.; Szekeres, T.; Nagai, M. Regulation of GTP biosynthesis. Adv. Enzym. Regul. 1992, 32, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Kuukasjärvi, A.; Landoni, J.C.; Kaukonen, J.; Juhakoski, M.; Auranen, M.; Torkkeli, T.; Velagapudi, V.; Suomalainen, A. IMPDH2: A new gene associated with dominant juvenile-onset dystonia-tremor disorder. Eur. J. Hum. Genet. 2021, 29, 1833–1837. [Google Scholar] [CrossRef]
- Zanon, A.; Rakovic, A.; Blankenburg, H.; Doncheva, N.T.; Schwienbacher, C.; Serafin, A.; Alexa, A.; Weichenberger, C.X.; Albrecht, M.; Klein, C.; et al. Profiling of Parkin-binding partners using tandem affinity purification. PLoS ONE 2013, 8, e78648. [Google Scholar] [CrossRef]
- Li, T.W.; Kenney, A.D.; Park, J.G.; Fiches, G.N.; Liu, H.; Zhou, D.; Biswas, A.; Zhao, W.; Que, J.; Santoso, N.; et al. SARS-CoV-2 Nsp14 protein associates with IMPDH2 and activates NF-κB signaling. Front. Immunol. 2022, 13, 1007089. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.X.; Song, X.M.; Wang, L.C.; Lv, H.N.; Chen, J.F.; Liu, D.; Fu, G.; Zhao, M.B.; Jiang, Y.; Zeng, K.W.; et al. Highly selective inhibition of IMPDH2 provides the basis of anti-neuroinflammation therapy. Proc. Natl. Acad. Sci. USA 2017, 114, E5986–E5994. [Google Scholar] [CrossRef]
- Available online: https://maayanlab.cloud/Harmonizome/gene_set/NF-KappaB/MotifMap+Predicted+Transcription+Factor+Targets (accessed on 1 June 2023).
- Brown, G.R.; Hem, V.; Katz, K.S.; Ovetsky, M.; Wallin, C.; Ermolaeva, O.; Tolstoy, I.; Tatusova, T.; Pruitt, K.D.; Maglott, D.R.; et al. Gene: A gene-centered information resource at NCBI. Nucleic Acids Res. 2015, 43, D36–D42. [Google Scholar] [CrossRef]
- Kaltschmidt, B.; Kaltschmidt, C. NF-KappaB in long-term memory and structural plasticity in the adult mammalian brain. Front. Mol. Neurosci. 2015, 8, 69. [Google Scholar] [CrossRef]
- Alberini, C.M. Transcription Factors in Long-Term Memory and Synaptic Plasticity. Physiol. Rev. 2009, 89, 121–145. [Google Scholar] [CrossRef]
- Marin, I.; Kipnis, J. Learning and memory... and the immune system. Learn. Mem. 2013, 20, 601–606. [Google Scholar] [CrossRef]
- Alani, B.; Salehi, R.; Sadeghi, P.; Zare, M.; Khodagholi, F.; Arefian, E.; Hakemi, M.G.; Digaleh, H. Silencing of Hsp90 chaperone expression protects against 6-hydroxydopamine toxicity in PC12 cells. J. Mol. Neurosci. 2014, 52, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Sanati, M.; Aminyavari, S.; Khodagholi, F.; Hajipour, M.J.; Sadeghi, P.; Noruzi, M.; Moshtagh, A.; Behmadi, H.; Sharifzadeh, M. PEGylated superparamagnetic iron oxide nanoparticles (SPIONs) ameliorate learning and memory deficit in a rat model of Alzheimer’s disease: Potential participation of STIMs. Neurotox 2021, 85, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef]
- Barnabei, L.; Laplantine, E.; Mbongo, W.; Rieux-Laucat, F.; Weil, R. NF-κB: At the Borders of Autoimmunity and Inflammation. Front. Immunol. 2021, 12, 716469. [Google Scholar] [CrossRef] [PubMed]
- Filiano, A.J.; Gadani, S.P.; Kipnis, J. Interactions of innate and adaptive immunity in brain development and function. Brain Res. 2015, 1617, 18–27. [Google Scholar] [CrossRef]
- Zhao, F.; Li, B.; Yang, W.; Ge, T.; Cui, R. Brain-immune interaction mechanisms: Implications for cognitive dysfunction in psychiatric disorders. Cell Prolif. 2022, 55, e13295. [Google Scholar] [CrossRef]
- Werneburg, S.; Feinberg, P.A.; Johnson, K.M.; Schafer, D.P. A microglia-cytokine axis to modulate synaptic connectivity and function. Curr. Opin. Neurobiol. 2017, 47, 138–145. [Google Scholar] [CrossRef]
- Brynskikh, A.; Warren, T.; Zhu, J.; Kipnis, J. Adaptive immunity affects learning behavior in mice. Brain Behav. Immun. 2008, 22, 861–869. [Google Scholar] [CrossRef]
- Ziv, Y.; Schwartz, M. Immune-based regulation of adult neurogenesis: Implications for learning and memory. Brain Behav. Immun. 2008, 22, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y. Vacuoles in mammals: A subcellular structure indispensable for early embryogenesis. Bioarchitecture 2013, 3, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Qi, C.; Luo, L.D.; Feng, I.; Ma, S. Molecular mechanisms of synaptogenesis. Front. Synaptic Neurosci. 2022, 14, 939793. [Google Scholar] [CrossRef] [PubMed]
- Batool, A.; Cheng, J.; Liu, Y.-X. Role of EZH2 in cell lineage determination and relative signaling pathways. Front. Biosci. 2019, 24, 947–960. [Google Scholar] [CrossRef]
- Bilbo, S.D.; Schwarz, J.M. The immune system and developmental programming of brain and behavior. Front. Neuroendocrinol. 2012, 33, 267–286. [Google Scholar] [CrossRef]
- Dantzer, R.; Cohen, S.; Russo, S.J.; Dinan, T.G. Resilience and immunity. Brain Behav. Immun. 2018, 74, 28–42. [Google Scholar] [CrossRef]
- Rollins, B.; Martin, M.V.; Morgan, L.; Vawter, M.P. Analysis of whole genome biomarker expression in blood and brain. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2010, 153, 919–936. [Google Scholar] [CrossRef]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostats 2003, 4, 249–264. [Google Scholar] [CrossRef]
- Dai, M.; Wang, P.; Boyd, A.D.; Kostov, G.; Athey, B.; Jones, E.G.; Bunney, W.E.; Myers, R.M.; Speed, T.P.; Akil, H.; et al. Evolving gene/transcript definitions significantly alter the interpretation of GeneChip data. Nucleic Acids Res. 2005, 33, e175. [Google Scholar] [CrossRef]
- Tang, Y.Y.; Tang, R.; Gross, J.J. Promoting Psychological Well-Being Through an Evidence-Based Mindfulness Training Program. Front. Hum. Neurosci. 2019, 13, 237. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | ACC | HC | Blood | Annotated Function |
|---|---|---|---|---|
| Card6 | 2.97 | 2.43 | 2.19 | Signal transduction modulation |
| Impdh2 | 2.08 | 2.26 | 2.18 | Nucleotide biosynthesis, cell growth |
| Dnase1l2 | 3.34 | 3.72 | 2.04 | DNA catabolism, programmed cell death |
| Gene | Pre, Control | Post, Control | Pre, Trained | Post, Trained |
|---|---|---|---|---|
| Card6 | 2.60 | 1.57 | 2.89 | 2.19 |
| Impdh2 | 1.32 | 1.99 | 1.92 | 1.79 |
| Dnase1l2 | 1.22 | 1.14 | 1.14 | 1.40 |
| Irf8 | 4.12 | 3.31 | 3.93 | 3.86 |
| Ikbkb | 4.01 | 3.35 | 3.74 | 3.62 |
| NF-κB2 | 2.95 | 2.58 | 3.22 | 2.82 |
| Ywhaz (reference) | 3.93 | 3.99 | 3.93 | 3.97 |
| Meditation (n = 14) | Memory (n = 12) | ||||||
|---|---|---|---|---|---|---|---|
| Sample | Card6 | Impdh2 | Dnase1l2 | Sample | Card6 | Impdh2 | Dnase1l2 |
| 11 | −0.482 | −1.616 | 0.422 | 1 | −1.320 | −0.833 | 0.255 |
| 13 | −0.064 | −0.197 | 0.279 | 2 | −0.026 | 0.315 | 0.391 |
| 14 | −0.766 | 0.191 | 0.113 | 3 | −0.898 | −0.010 | −0.020 |
| 15 | −0.014 | −0.934 | −0.470 | 4 | −1.366 | 0.757 | −1.100 |
| 16 | −1.339 | −0.510 | 0.176 | 6 | 0.038 | −0.855 | −0.407 |
| 17 | −0.542 | −0.748 | −0.566 | 7 | 0.518 | −0.736 | −0.319 |
| 18 | −0.231 | −0.791 | −0.228 | 8 | 0.508 | −0.314 | −0.851 |
| 19 | −0.158 | −0.754 | 0.135 | 9 | −0.229 | −0.448 | −0.101 |
| 20 | −0.081 | 0.021 | −0.227 | 29 | −1.391 | 0.004 | −0.006 |
| 22 | −1.241 | 0.906 | 0.060 | 30 | −0.155 | −0.057 | −0.228 |
| 23 | −0.585 | 0.471 | 0.432 | 31 | 0.119 | −0.230 | 0.467 |
| 25 | −1.968 | 0.489 | 0.229 | 32 | −0.658 | 0.117 | −0.053 |
| 26 | −0.701 | −0.125 | −0.474 | ||||
| 27 | −0.003 | −0.244 | 0.242 | ||||
| Mean | −0.584 | −0.274 | 0.009 | Mean | −0.405 | −0.191 | −0.164 |
| SD | 0.586 | 0.672 | 0.338 | SD | 0.703 | 0.484 | 0.465 |
| Fold change | 1.50× | 1.21× | 0.99× | Fold change | 1.32× | 1.14× | 1.12× |
| Gene | Irf8 | NF-κB2 | Ikbkb | |||
|---|---|---|---|---|---|---|
| Condition | Memory | Meditation | Memory | Meditation | Memory | Meditation |
| Mean | −0.308 | 0.655 | −0.181 | 0.562 | −0.270 | 0.161 |
| SD | 1.159 | 0.992 | 0.491 | 0.469 | 0.505 | 0.380 |
| fold change | 1.24× | 0.64× | 1.13× | 0.68× | 1.21× | 0.89× |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voelker, P.; Weible, A.P.; Niell, C.M.; Rothbart, M.K.; Posner, M.I. Molecular Mechanisms for Changing Brain Connectivity in Mice and Humans. Int. J. Mol. Sci. 2023, 24, 15840. https://doi.org/10.3390/ijms242115840
Voelker P, Weible AP, Niell CM, Rothbart MK, Posner MI. Molecular Mechanisms for Changing Brain Connectivity in Mice and Humans. International Journal of Molecular Sciences. 2023; 24(21):15840. https://doi.org/10.3390/ijms242115840
Chicago/Turabian StyleVoelker, Pascale, Aldis P. Weible, Cristopher M. Niell, Mary K. Rothbart, and Michael I. Posner. 2023. "Molecular Mechanisms for Changing Brain Connectivity in Mice and Humans" International Journal of Molecular Sciences 24, no. 21: 15840. https://doi.org/10.3390/ijms242115840
APA StyleVoelker, P., Weible, A. P., Niell, C. M., Rothbart, M. K., & Posner, M. I. (2023). Molecular Mechanisms for Changing Brain Connectivity in Mice and Humans. International Journal of Molecular Sciences, 24(21), 15840. https://doi.org/10.3390/ijms242115840